

Induced Remodelling of Astrocytes In Vitro and In Vivo by Manipulation of Astrocytic RhoA Activity
 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Design and Production of Recombinant Adeno-Associated Viruses (rAAVs)
2.3. Stereotactic Virus Injections
2.4. Preparation and Transduction of Dissociated Cultures
2.5. Immunohistochemistry
2.6. Microscopy and Image Analysis
2.7. Statistical Analysis
3. Results
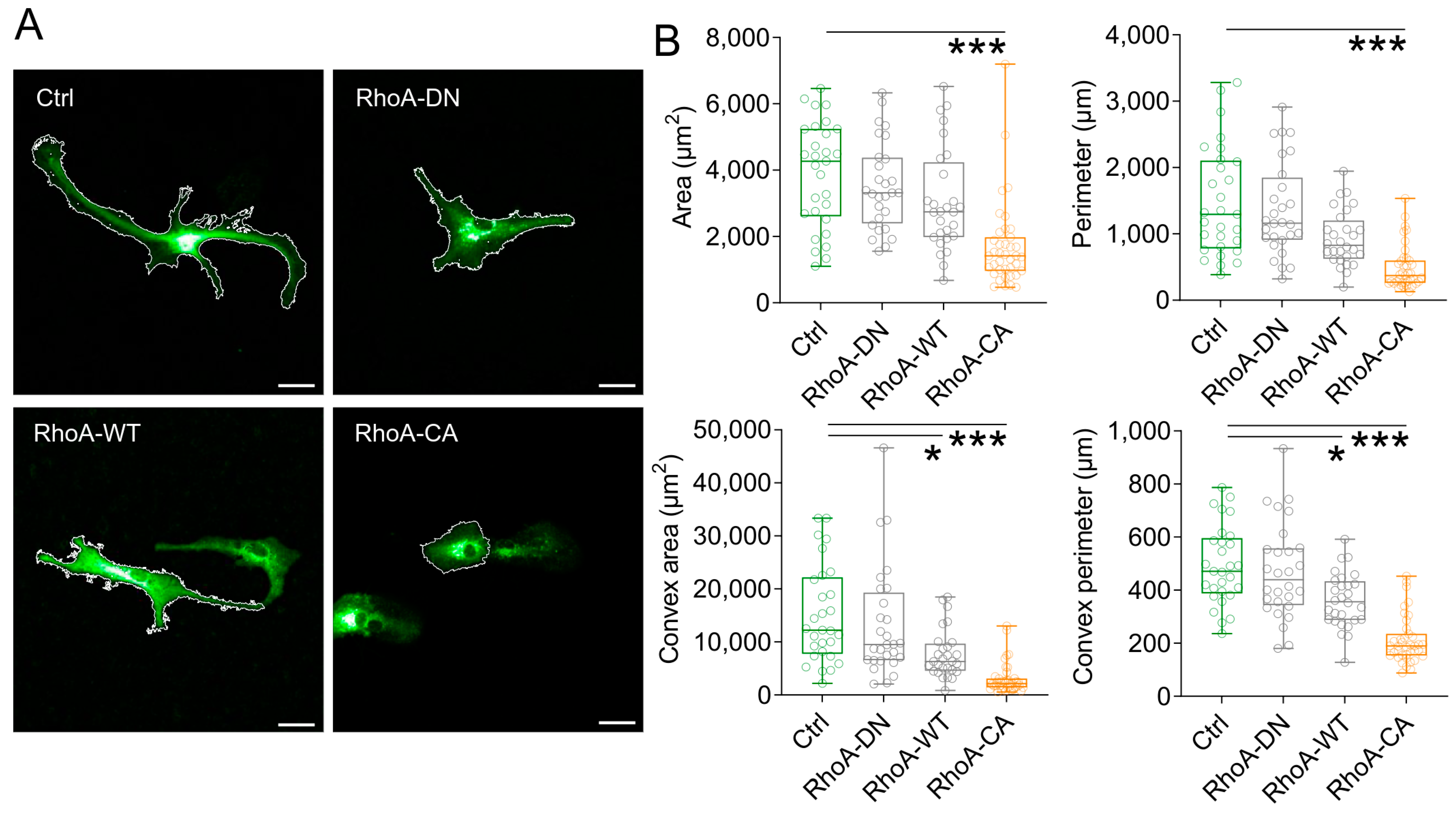
3.1. Increased RhoA Activity Modifies Astrocyte Structure In Vitro
3.2. Increased RhoA Activity Alters Astrocytic Actin Assembly in Vitro
3.3. Remodelling of Astrocytes In Vivo by Increased RhoA Activity
3.4. Unaltered Large Process Structure after Increasing RhoA Activity
4. Discussion
4.1. Mechanisms Underlying RhoA-CA Effect on Astrocyte Morphology
4.2. Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chai, H.; Diaz-Castro, B.; Shigetomi, E.; Monte, E.; Octeau, J.C.; Yu, X.; Cohn, W.; Rajendran, P.S.; Vondriska, T.M.; Whitelegge, J.P.; et al. Neural Circuit-Specialized Astrocytes: Transcriptomic, Proteomic, Morphological, and Functional Evidence. Neuron 2017, 95, 531–549.e9. [Google Scholar] [CrossRef]
- Lanjakornsiripan, D.; Pior, B.-J.; Kawaguchi, D.; Furutachi, S.; Tahara, T.; Katsuyama, Y.; Suzuki, Y.; Fukazawa, Y.; Gotoh, Y. Layer-Specific Morphological and Molecular Differences in Neocortical Astrocytes and Their Dependence on Neuronal Layers. Nat. Commun. 2018, 9, 1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minge, D.; Domingos, C.; Unichenko, P.; Behringer, C.; Pauletti, A.; Anders, S.; Herde, M.K.; Delekate, A.; Gulakova, P.; Schoch, S.; et al. Heterogeneity and Development of Fine Astrocyte Morphology Captured by Diffraction-Limited Microscopy. Front. Cell. Neurosci. 2021, 15, 669280. [Google Scholar] [CrossRef] [PubMed]
- Bushong, E.A.; Martone, M.E.; Ellisman, M.H. Maturation of Astrocyte Morphology and the Establishment of Astrocyte Domains during Postnatal Hippocampal Development. Int. J. Dev. Neurosci. 2004, 22, 73–86. [Google Scholar] [CrossRef]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.R.; Robitaille, R.; Volterra, A. Gliotransmitters Travel in Time and Space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef] [Green Version]
- Rusakov, D.A.; Bard, L.; Stewart, M.G.; Henneberger, C. Diversity of Astroglial Functions Alludes to Subcellular Specialisation. Trends Neurosci. 2014, 37, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Heller, J.P.; Rusakov, D.A. The Nanoworld of the Tripartite Synapse: Insights from Super-Resolution Microscopy. Front. Cell. Neurosci. 2017, 11, 374. [Google Scholar] [CrossRef] [Green Version]
- Arizono, M.; Inavalli, V.V.G.K.; Bancelin, S.; Fernández-Monreal, M.; Nägerl, U.V. Super-Resolution Shadow Imaging Reveals Local Remodeling of Astrocytic Microstructures and Brain Extracellular Space after Osmotic Challenge. Glia 2021, 69, 1605–1613. [Google Scholar] [CrossRef]
- Aten, S.; Kiyoshi, C.M.; Arzola, E.P.; Patterson, J.A.; Taylor, A.T.; Du, Y.; Guiher, A.M.; Philip, M.; Camacho, E.G.; Mediratta, D.; et al. Ultrastructural View of Astrocyte Arborization, Astrocyte-Astrocyte and Astrocyte-Synapse Contacts, Intracellular Vesicle-like Structures, and Mitochondrial Network. Prog. Neurobiol. 2022, 213, 102264. [Google Scholar] [CrossRef]
- Haber, M.; Zhou, L.; Murai, K.K. Cooperative Astrocyte and Dendritic Spine Dynamics at Hippocampal Excitatory Synapses. J. Neurosci. 2006, 26, 8881–8891. [Google Scholar] [CrossRef] [Green Version]
- Hirrlinger, J.; Hülsmann, S.; Kirchhoff, F. Astroglial Processes Show Spontaneous Motility at Active Synaptic Terminals in Situ. Eur. J. Neurosci. 2004, 20, 2235–2239. [Google Scholar] [CrossRef] [PubMed]
- Bernardinelli, Y.; Randall, J.; Janett, E.; Nikonenko, I.; König, S.; Jones, E.V.; Flores, C.E.; Murai, K.K.; Bochet, C.G.; Holtmaat, A.; et al. Activity-Dependent Structural Plasticity of Perisynaptic Astrocytic Domains Promotes Excitatory Synapse Stability. Curr. Biol. 2014, 24, 1679–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henneberger, C.; Bard, L.; Panatier, A.; Reynolds, J.P.; Kopach, O.; Medvedev, N.I.; Minge, D.; Herde, M.K.; Anders, S.; Kraev, I.; et al. LTP Induction Boosts Glutamate Spillover by Driving Withdrawal of Perisynaptic Astroglia. Neuron 2020, 108, 919–936.e11. [Google Scholar] [CrossRef]
- Lushnikova, I.; Skibo, G.; Muller, D.; Nikonenko, I. Synaptic Potentiation Induces Increased Glial Coverage of Excitatory Synapses in CA1 Hippocampus. Hippocampus 2009, 19, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, J.; Lammert, G.; Meyer, U.; Krug, M. The Influence of Long-Term Potentiation on the Spatial Relationship between Astrocyte Processes and Potentiated Synapses in the Dentate Gyrus Neuropil of Rat Brain. Brain Res. 1991, 560, 122–131. [Google Scholar] [CrossRef]
- Theodosis, D.T.; Poulain, D.A. Activity-Dependent Neuronal-Glial and Synaptic Plasticity in the Adult Mammalian Hypothalamus. Neuroscience 1993, 57, 501–535. [Google Scholar] [CrossRef]
- McCauley, J.P.; Petroccione, M.A.; D’Brant, L.Y.; Todd, G.C.; Affinnih, N.; Wisnoski, J.J.; Zahid, S.; Shree, S.; Sousa, A.A.; De Guzman, R.M.; et al. Circadian Modulation of Neurons and Astrocytes Controls Synaptic Plasticity in Hippocampal Area CA1. Cell Rep. 2020, 33, 108255. [Google Scholar] [CrossRef]
- Bellesi, M.; de Vivo, L.; Tononi, G.; Cirelli, C. Effects of Sleep and Wake on Astrocytes: Clues from Molecular and Ultrastructural Studies. BMC Biol. 2015, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Genoud, C.; Quairiaux, C.; Steiner, P.; Hirling, H.; Welker, E.; Knott, G.W. Plasticity of Astrocytic Coverage and Glutamate Transporter Expression in Adult Mouse Cortex. PLoS Biol. 2006, 4, e343. [Google Scholar] [CrossRef] [Green Version]
- Popov, A.; Denisov, P.; Bychkov, M.; Brazhe, A.; Lyukmanova, E.; Shenkarev, Z.; Lazareva, N.; Verkhratsky, A.; Semyanov, A. Caloric Restriction Triggers Morphofunctional Remodeling of Astrocytes and Enhances Synaptic Plasticity in the Mouse Hippocampus. Cell Death Dis. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Herde, M.K.; Bohmbach, K.; Domingos, C.; Vana, N.; Komorowska-Müller, J.A.; Passlick, S.; Schwarz, I.; Jackson, C.J.; Dietrich, D.; Schwarz, M.K.; et al. Local Efficacy of Glutamate Uptake Decreases with Synapse Size. Cell Rep. 2020, 32, 108182. [Google Scholar] [CrossRef]
- Oliet, S.H.; Piet, R.; Poulain, D.A. Control of Glutamate Clearance and Synaptic Efficacy by Glial Coverage of Neurons. Science 2001, 292, 923–926. [Google Scholar] [CrossRef]
- Zeug, A.; Müller, F.E.; Anders, S.; Herde, M.K.; Minge, D.; Ponimaskin, E.; Henneberger, C. Control of Astrocyte Morphology by Rho GTPases. Brain Res. Bull. 2018, 136, 44–53. [Google Scholar] [CrossRef]
- Ramakers, G.J.A.; Moolenaar, W.H. Regulation of Astrocyte Morphology by RhoA and Lysophosphatidic Acid. Exp. Cell Res. 1998, 245, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Kalman, D.; Gomperts, S.N.; Hardy, S.; Kitamura, M.; Bishop, J.M. Ras Family GTPases Control Growth of Astrocyte Processes. Mol. Biol. Cell 1999, 10, 1665–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, V.; Bouschet, T.; Fernandez, C.; Bockaert, J.; Journot, L. Dynamic Reorganization of the Astrocyte Actin Cytoskeleton Elicited by CAMP and PACAP: A Role for PhosphatidylInositol 3-Kinase Inhibition. Eur. J. Neurosci. 2005, 21, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Murk, K.; Suarez, E.M.B.; Cockbill, L.M.R.; Banks, P.; Hanley, J.G. The Antagonistic Modulation of Arp2/3 Activity by N-WASP, WAVE2 and PICK1 Defines Dynamic Changes in Astrocyte Morphology. J. Cell Sci. 2013, 126, 3873–3883. [Google Scholar] [CrossRef] [Green Version]
- Stern, S.; Hilton, B.J.; Burnside, E.R.; Dupraz, S.; Handley, E.E.; Gonyer, J.M.; Brakebusch, C.; Bradke, F. RhoA Drives Actin Compaction to Restrict Axon Regeneration and Astrocyte Reactivity after CNS Injury. Neuron 2021, 109, 3436–3455.e9. [Google Scholar] [CrossRef]
- Müller, F.E.; Schade, S.K.; Cherkas, V.; Stopper, L.; Breithausen, B.; Minge, D.; Varbanov, H.; Wahl-Schott, C.; Antoniuk, S.; Domingos, C.; et al. Serotonin Receptor 4 Regulates Hippocampal Astrocyte Morphology and Function. Glia 2021, 69, 872–889. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A. Rho Gtpases: Integrating Integrin Signaling. J. Cell Biol. 2000, 150, F107–F109. [Google Scholar] [CrossRef]
- Zhang, X.; Machii, T.; Matsumura, I.; Ezoe, S.; Kawasaki, A.; Tanaka, H.; Ueda, S.; Sugahara, H.; Shibayama, H.; Mizuki, M.; et al. Constitutively Activated Rho Guanosine Triphosphatases Regulate the Growth and Morphology of Hairy Cell Leukemia Cells. Int. J. Hematol. 2003, 77, 263–273. [Google Scholar] [CrossRef]
- Nolte, C.; Matyash, M.; Pivneva, T.; Schipke, C.G.; Ohlemeyer, C.; Hanisch, U.K.; Kirchhoff, F.; Kettenmann, H. GFAP Promoter-Controlled EGFP-Expressing Transgenic Mice: A Tool to Visualize Astrocytes and Astrogliosis in Living Brain Tissue. Glia 2001, 33, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Marvin, J.S.; Borghuis, B.G.; Tian, L.; Cichon, J.; Harnett, M.T.; Akerboom, J.; Gordus, A.; Renninger, S.L.; Chen, T.-W.; Bargmann, C.I.; et al. An Optimized Fluorescent Probe for Visualizing Glutamate Neurotransmission. Nat. Methods 2013, 10, 162–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, L.C.; Mittag, M.; Poll, S.; Steffen, J.; Wagner, J.; Geis, H.-R.; Schwarz, I.; Schmidt, B.; Schwarz, M.K.; Remy, S.; et al. Dysfunction of Somatostatin-Positive Interneurons Associated with Memory Deficits in an Alzheimer’s Disease Model. Neuron 2016, 92, 114–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woitecki, A.M.H.; Müller, J.A.; van Loo, K.M.J.; Sowade, R.F.; Becker, A.J.; Schoch, S. Identification of Synaptotagmin 10 as Effector of NPAS4-Mediated Protection from Excitotoxic Neurodegeneration. J. Neurosci. 2016, 36, 2561–2570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfes, A.C.; Ahmed, S.; Awasthi, A.; Stahlberg, M.A.; Rajput, A.; Magruder, D.S.; Bonn, S.; Dean, C. A Novel Method for Culturing Stellate Astrocytes Reveals Spatially Distinct Ca2+ Signaling and Vesicle Recycling in Astrocytic Processes. J. Gen. Physiol. 2016, 149, 149–170. [Google Scholar] [CrossRef] [Green Version]
- Pirnat, S.; Božić, M.; Dolanc, D.; Horvat, A.; Tavčar, P.; Vardjan, N.; Verkhratsky, A.; Zorec, R.; Stenovec, M. Astrocyte Arborization Enhances Ca2+ but Not CAMP Signaling Plasticity. Glia 2021, 69, 2899–2916. [Google Scholar] [CrossRef]
- Badia-Soteras, A.; de Vries, J.; Dykstra, W.; Broersen, L.M.; Verkuyl, J.M.; Smit, A.B.; Verheijen, M.H.G. High-Throughput Analysis of Astrocyte Cultures Shows Prevention of Reactive Astrogliosis by the Multi-Nutrient Combination Fortasyn Connect. Cells 2022, 11, 1428. [Google Scholar] [CrossRef]
- Zack, G.W.; Rogers, W.E.; Latt, S.A. Automatic Measurement of Sister Chromatid Exchange Frequency. J. Histochem. Cytochem. 1977, 25, 741–753. [Google Scholar] [CrossRef]
- Hansen, J.N.; Brückner, M.; Pietrowski, M.J.; Jikeli, J.F.; Plescher, M.; Beckert, H.; Schnaars, M.; Fülle, L.; Reitmeier, K.; Langmann, T.; et al. MotiQ: An Open-Source Toolbox to Quantify the Cell Motility and Morphology of Microglia. Mol. Biol. Cell 2022, 33, ar99. [Google Scholar] [CrossRef]
- Anders, S.; Minge, D.; Griemsmann, S.; Herde, M.K.; Steinhäuser, C.; Henneberger, C. Spatial Properties of Astrocyte Gap Junction Coupling in the Rat Hippocampus. Phil. Trans. R. Soc. B 2014, 369, 20130600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medvedev, N.; Popov, V.; Henneberger, C.; Kraev, I.; Rusakov, D.A.; Stewart, M.G. Glia Selectively Approach Synapses on Thin Dendritic Spines. Phil. Trans. R. Soc. B 2014, 369, 20140047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maekawa, M.; Ishizaki, T.; Boku, S.; Watanabe, N.; Fujita, A.; Iwamatsu, A.; Obinata, T.; Ohashi, K.; Mizuno, K.; Narumiya, S. Signaling from Rho to the Actin Cytoskeleton Through Protein Kinases ROCK and LIM-Kinase. Science 1999, 285, 895–898. [Google Scholar] [CrossRef]
- Sholl, D.A. Dendritic Organization in the Neurons of the Visual and Motor Cortices of the Cat. J. Anat. 1953, 87, 387–406.1. [Google Scholar]
- Korogod, N.; Petersen, C.C.; Knott, G.W. Ultrastructural Analysis of Adult Mouse Neocortex Comparing Aldehyde Perfusion with Cryo Fixation. eLife 2015, 4, e05793. [Google Scholar] [CrossRef]
- King, C.M.; Bohmbach, K.; Minge, D.; Delekate, A.; Zheng, K.; Reynolds, J.; Rakers, C.; Zeug, A.; Petzold, G.C.; Rusakov, D.A.; et al. Local Resting Ca2+ Controls the Scale of Astroglial Ca2+ Signals. Cell Rep. 2020, 30, 3466–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namme, J.N.; Bepari, A.K.; Takebayashi, H. Cofilin Signaling in the CNS Physiology and Neurodegeneration. Int. J. Mol. Sci. 2021, 22, 10727. [Google Scholar] [CrossRef]
- Nishida, H.; Okabe, S. Direct Astrocytic Contacts Regulate Local Maturation of Dendritic Spines. J. Neurosci. 2007, 27, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Robel, S.; Bardehle, S.; Lepier, A.; Brakebusch, C.; Götz, M. Genetic Deletion of Cdc42 Reveals a Crucial Role for Astrocyte Recruitment to the Injury Site in Vitro and in Vivo. J. Neurosci. 2011, 31, 12471–12482. [Google Scholar] [CrossRef] [Green Version]
- Racchetti, G.; D’Alessandro, R.; Meldolesi, J. Astrocyte Stellation, a Process Dependent on Rac1 Is Sustained by the Regulated Exocytosis of Enlargeosomes. Glia 2012, 60, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Vardjan, N.; Kreft, M.; Zorec, R. Dynamics of β-Adrenergic/CAMP Signaling and Morphological Changes in Cultured Astrocytes. Glia 2014, 62, 566–579. [Google Scholar] [CrossRef]
- Schiweck, J.; Eickholt, B.J.; Murk, K. Important Shapeshifter: Mechanisms Allowing Astrocytes to Respond to the Changing Nervous System During Development, Injury and Disease. Front. Cell. Neurosci. 2018, 12, 261. [Google Scholar] [CrossRef] [Green Version]
- Tsukita, S.; Yonemura, S. Cortical Actin Organization: Lessons from ERM (Ezrin/Radixin/Moesin) Proteins. J. Biol. Chem. 1999, 274, 34507–34510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehon, R.G.; McClatchey, A.I.; Bretscher, A. Organizing the Cell Cortex: The Role of ERM Proteins. Nat. Rev. Mol. Cell Biol. 2010, 11, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Derouiche, A.; Frotscher, M. Peripheral Astrocyte Processes: Monitoring by Selective Immunostaining for the Actin-Binding ERM Proteins. Glia 2001, 36, 330–341. [Google Scholar] [CrossRef]
- Lavialle, M.; Aumann, G.; Anlauf, E.; Pröls, F.; Arpin, M.; Derouiche, A. Structural Plasticity of Perisynaptic Astrocyte Processes Involves Ezrin and Metabotropic Glutamate Receptors. Proc. Natl. Acad. Sci. USA 2011, 108, 12915–12919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bros, M.; Haas, K.; Moll, L.; Grabbe, S. RhoA as a Key Regulator of Innate and Adaptive Immunity. Cells 2019, 8, 733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunskine, C.; Passlick, S.; Henneberger, C. Structural Heterogeneity of the GABAergic Tripartite Synapse. Cells 2022, 11, 3150. [Google Scholar] [CrossRef]
- Endo, F.; Kasai, A.; Soto, J.S.; Yu, X.; Qu, Z.; Hashimoto, H.; Gradinaru, V.; Kawaguchi, R.; Khakh, B.S. Molecular Basis of Astrocyte Diversity and Morphology across the CNS in Health and Disease. Science 2022, 378, eadc9020. [Google Scholar] [CrossRef] [PubMed]
- Badia-Soteras, A.; Heistek, T.S.; Kater, M.S.J.; Mak, A.; Negrean, A.; Oever, M.C.V.D.; Mansvelder, H.D.; Khakh, B.S.; Min, R.; Smit, A.B.; et al. Retraction of Astrocyte Leaflets from the Synapse Enhances Fear Memory. Biol. Psychiatry 2022. [Google Scholar] [CrossRef]
- Renault-Mihara, F.; Mukaino, M.; Shinozaki, M.; Kumamaru, H.; Kawase, S.; Baudoux, M.; Ishibashi, T.; Kawabata, S.; Nishiyama, Y.; Sugai, K.; et al. Regulation of RhoA by STAT3 Coordinates Glial Scar Formation. J. Cell Biol. 2017, 216, 2533–2550. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domingos, C.; Müller, F.E.; Passlick, S.; Wachten, D.; Ponimaskin, E.; Schwarz, M.K.; Schoch, S.; Zeug, A.; Henneberger, C. Induced Remodelling of Astrocytes In Vitro and In Vivo by Manipulation of Astrocytic RhoA Activity. Cells 2023, 12, 331. https://doi.org/10.3390/cells12020331
Domingos C, Müller FE, Passlick S, Wachten D, Ponimaskin E, Schwarz MK, Schoch S, Zeug A, Henneberger C. Induced Remodelling of Astrocytes In Vitro and In Vivo by Manipulation of Astrocytic RhoA Activity. Cells. 2023; 12(2):331. https://doi.org/10.3390/cells12020331
Chicago/Turabian StyleDomingos, Cátia, Franziska E. Müller, Stefan Passlick, Dagmar Wachten, Evgeni Ponimaskin, Martin K. Schwarz, Susanne Schoch, André Zeug, and Christian Henneberger. 2023. "Induced Remodelling of Astrocytes In Vitro and In Vivo by Manipulation of Astrocytic RhoA Activity" Cells 12, no. 2: 331. https://doi.org/10.3390/cells12020331
APA StyleDomingos, C., Müller, F. E., Passlick, S., Wachten, D., Ponimaskin, E., Schwarz, M. K., Schoch, S., Zeug, A., & Henneberger, C. (2023). Induced Remodelling of Astrocytes In Vitro and In Vivo by Manipulation of Astrocytic RhoA Activity. Cells, 12(2), 331. https://doi.org/10.3390/cells12020331