
Effects of Dickkopf-1 (DKK-1) on Prostate Cancer Growth and Bone Metastasis

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Conditioned Medium (CM)
2.2. In Vitro Proliferation and Migration
2.3. Murine Primary Bone Cell Differentiation Assay
2.4. RNA Extraction and qRT-PCR
2.5. Intratibial and Intracardiac Injections of Cancer Cells into Aythmic Nude Mice
2.6. Bioluminescent Imaging
2.7. Micro-Computed Tomography (µCT)
2.8. Histopathology
2.9. Protein Extraction and Immunoblots
2.10. Statistical Analysis
3. Results
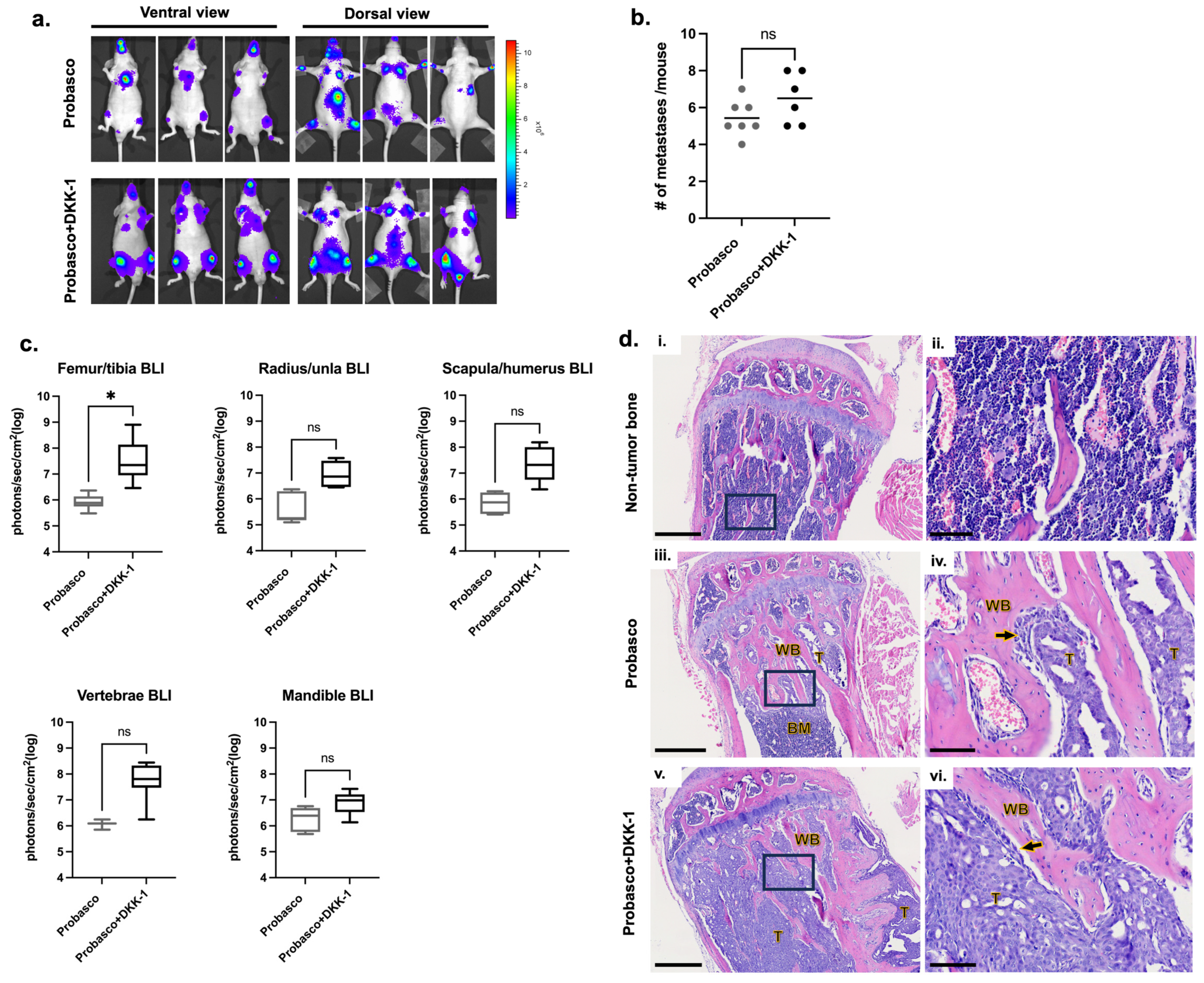
3.1. DKK-1 Did Not Affect the Incidence of Cancer Metastases but Induced Larger Tumors at Metastatic Bone Sites
3.2. DKK-1 Increased Cancer Growth and Decreased Cancer-Induced Bone Formation
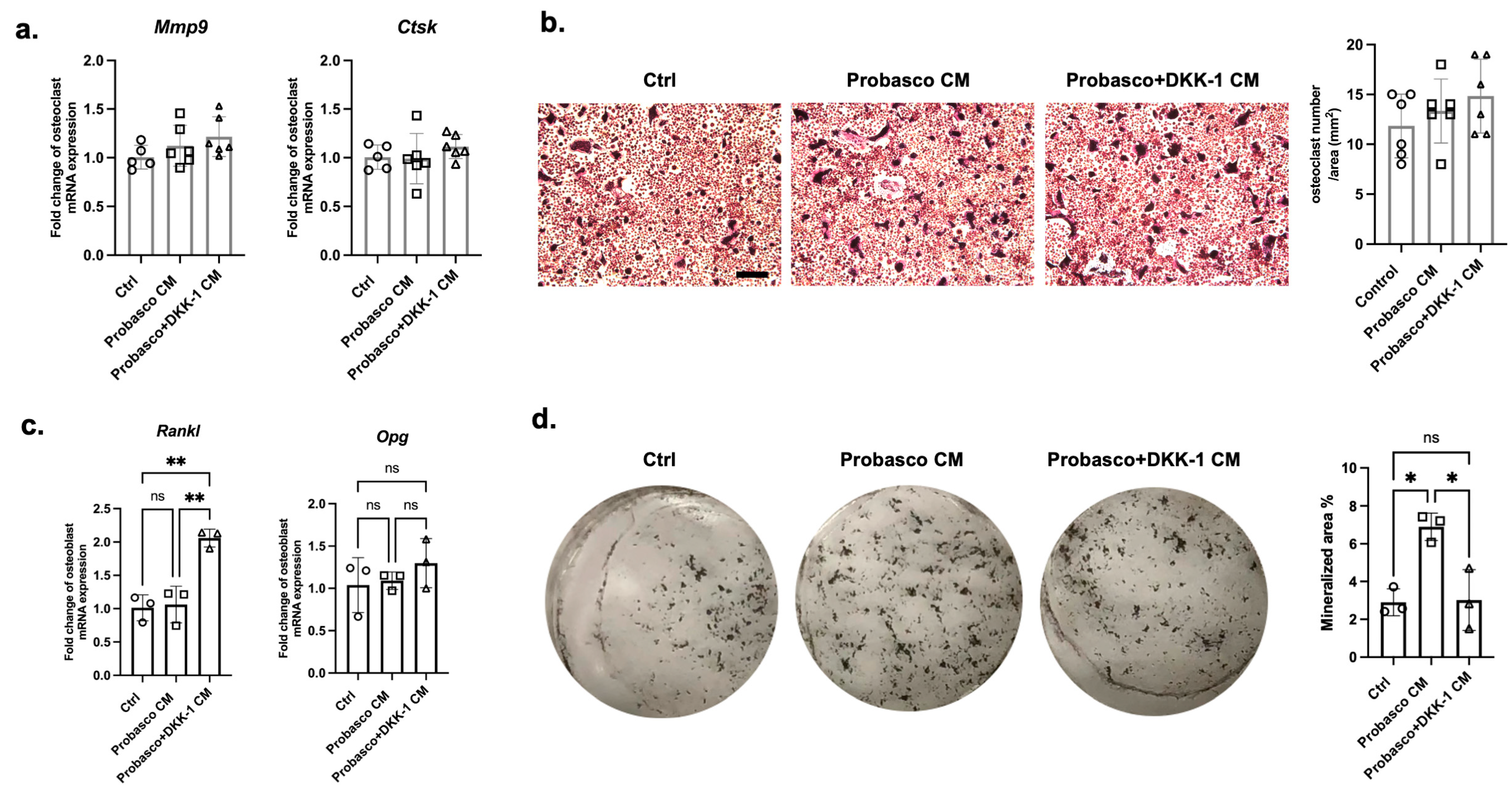
3.3. Effects of DKK-1 on Osteoblasts and Osteoclasts
3.4. DKK-1 Stimulated PCa Cell Proliferation and Migration and Induced EMT
3.5. The Cancer-Promoting Effect of DKK-1 Was Independent of the Canonical WNT Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, S.C.; Yu-Lee, L.Y.; Lin, S.H. Osteoblastic Factors in Prostate Cancer Bone Metastasis. Curr. Osteoporos. Rep. 2018, 16, 642–647. [Google Scholar] [CrossRef]
- Murillo-Garzon, V.; Kypta, R. WNT signalling in prostate cancer. Nat. Rev. Urol. 2017, 14, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Nishimori, H.; Ehata, S.; Suzuki, H.I.; Katsuno, Y.; Miyazono, K. Prostate cancer cells and bone stromal cells mutually interact with each other through bone morphogenetic protein-mediated signals. J. Biol. Chem. 2012, 287, 20037–20046. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Kitagawa, Y.; Zhang, J.; Yao, Z.; Mizokami, A.; Cheng, S.; Nor, J.; McCauley, L.K.; Taichman, R.S.; Keller, E.T. Vascular endothelial growth factor contributes to the prostate cancer-induced osteoblast differentiation mediated by bone morphogenetic protein. Cancer Res. 2004, 64, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Saber, S.H.; Ali, H.E.A.; Gaballa, R.; Gaballah, M.; Ali, H.I.; Zerfaoui, M.; Abd Elmageed, Z.Y. Exosomes are the Driving Force in Preparing the Soil for the Metastatic Seeds: Lessons from the Prostate Cancer. Cells 2020, 9, 564. [Google Scholar] [CrossRef]
- Scimeca, M.; Urbano, N.; Bonfiglio, R.; Mapelli, S.N.; Catapano, C.V.; Carbone, G.M.; Ciuffa, S.; Tavolozza, M.; Schillaci, O.; Mauriello, A.; et al. Prostate Osteoblast-Like Cells: A Reliable Prognostic Marker of Bone Metastasis in Prostate Cancer Patients. Contrast Media Mol. Imaging 2018, 2018, 9840962. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Song, J.; Chen, W.; Yuan, D.; Wang, W.; Chen, X.; Liu, H.; Su, H.; Zhu, J. Expression and Role of Dickkopf-1 (Dkk1) in Tumors: From the Cells to the Patients. Cancer Manag. Res. 2021, 13, 659–675. [Google Scholar] [CrossRef] [PubMed]
- Pinzone, J.J.; Hall, B.M.; Thudi, N.K.; Vonau, M.; Qiang, Y.W.; Rosol, T.J.; Shaughnessy, J.D., Jr. The role of Dickkopf-1 in bone development, homeostasis, and disease. Blood 2009, 113, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Yang, K.E.; Hwang, I.H.; Huh, Y.H.; Kim, D.J.; Yoo, H.S.; Park, S.J.; Jang, I.S. Cordycepin inhibits human ovarian cancer by inducing autophagy and apoptosis through Dickkopf-related protein 1/beta-catenin signaling. Am. J. Transl. Res. 2019, 11, 6890–6906. [Google Scholar]
- Liu, Z.; Sun, B.; Qi, L.; Li, Y.; Zhao, X.; Zhang, D.; Zhang, Y. Dickkopf-1 expression is down-regulated during the colorectal adenoma-carcinoma sequence and correlates with reduced microvessel density and VEGF expression. Histopathology 2015, 67, 158–166. [Google Scholar] [CrossRef]
- Krause, U.; Ryan, D.M.; Clough, B.H.; Gregory, C.A. An unexpected role for a Wnt-inhibitor: Dickkopf-1 triggers a novel cancer survival mechanism through modulation of aldehyde-dehydrogenase-1 activity. Cell Death Dis. 2014, 5, e1093. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.H.; Liu, Z.B.; Yang, C.; Qin, W.; Shao, Z.M. Expression of dickkopf-1 and beta-catenin related to the prognosis of breast cancer patients with triple negative phenotype. PLoS ONE 2012, 7, e37624. [Google Scholar] [CrossRef] [PubMed]
- Thudi, N.K.; Martin, C.K.; Murahari, S.; Shu, S.T.; Lanigan, L.G.; Werbeck, J.L.; Keller, E.T.; McCauley, L.K.; Pinzone, J.J.; Rosol, T.J. Dickkopf-1 (DKK-1) stimulated prostate cancer growth and metastasis and inhibited bone formation in osteoblastic bone metastases. Prostate 2011, 71, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, A.; Matsumoto, S.; Sada, R. Dickkopf signaling, beyond Wnt-mediated biology. Semin. Cell Dev. Biol. 2022, 125, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Fumoto, K.; Shojima, K.; Nojima, S.; Osugi, Y.; Tomihara, H.; Eguchi, H.; Shintani, Y.; Endo, H.; Inoue, M.; et al. CKAP4 is a Dickkopf1 receptor and is involved in tumor progression. J. Clin. Investig. 2016, 126, 2689–2705. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Hu, J. The role of JNK in prostate cancer progression and therapeutic strategies. Biomed. Pharmacother. 2020, 121, 109679. [Google Scholar] [CrossRef]
- Holl, M.; Koziel, R.; Schafer, G.; Pircher, H.; Pauck, A.; Hermann, M.; Klocker, H.; Jansen-Durr, P.; Sampson, N. ROS signaling by NADPH oxidase 5 modulates the proliferation and survival of prostate carcinoma cells. Mol. Carcinog. 2016, 55, 27–39. [Google Scholar] [CrossRef]
- Supsavhad, W.; Hassan, B.B.; Simmons, J.K.; Dirksen, W.P.; Elshafae, S.M.; Kohart, N.A.; Demirer, A.A.; Rosol, T.J. Effect of Dickkopf-1 (Dkk-1) and SP600125, a JNK Inhibitor, on Wnt Signaling in Canine Prostate Cancer Growth and Bone Metastases. Vet. Sci. 2021, 8, 153. [Google Scholar] [CrossRef]
- Xie, H.; Liu, T.; Chen, J.; Yang, Z.; Xu, S.; Fan, Y.; Zeng, J.; Chen, Y.; Ma, Z.; Gao, Y.; et al. Activation of PSGR with beta-ionone suppresses prostate cancer progression by blocking androgen receptor nuclear translocation. Cancer Lett. 2019, 453, 193–205. [Google Scholar] [CrossRef]
- Verzella, D.; Pescatore, A.; Capece, D.; Vecchiotti, D.; Ursini, M.V.; Franzoso, G.; Alesse, E.; Zazzeroni, F. Life, death, and autophagy in cancer: NF-kappaB turns up everywhere. Cell Death Dis. 2020, 11, 210. [Google Scholar] [CrossRef]
- Grosset, A.A.; Ouellet, V.; Caron, C.; Fragoso, G.; Barres, V.; Delvoye, N.; Latour, M.; Aprikian, A.; Bergeron, A.; Chevalier, S.; et al. Validation of the prognostic value of NF-kappaB p65 in prostate cancer: A retrospective study using a large multi-institutional cohort of the Canadian Prostate Cancer Biomarker Network. PLoS Med. 2019, 16, e1002847. [Google Scholar] [CrossRef]
- Papa, S.; Bubici, C.; Zazzeroni, F.; Pham, C.G.; Kuntzen, C.; Knabb, J.R.; Dean, K.; Franzoso, G. The NF-kappaB-mediated control of the JNK cascade in the antagonism of programmed cell death in health and disease. Cell Death Differ. 2006, 13, 712–729. [Google Scholar] [CrossRef]
- Simmons, J.K.; Hildreth, B.E., 3rd; Supsavhad, W.; Elshafae, S.M.; Hassan, B.B.; Dirksen, W.P.; Toribio, R.E.; Rosol, T.J. Animal Models of Bone Metastasis. Vet. Pathol. 2015, 52, 827–841. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Kantake, N.; Hellmann, D.E.; Elshafae, S.M.; Rosol, T.J. Stimulation of in vitro bone formation by canine prostate cancer. J. Cancer Metastasis Treat. 2021, 7, 37. [Google Scholar] [CrossRef]
- Simmons, J.K.; Dirksen, W.P.; Hildreth, B.E., 3rd; Dorr, C.; Williams, C.; Thomas, R.; Breen, M.; Toribio, R.E.; Rosol, T.J. Canine prostate cancer cell line (Probasco) produces osteoblastic metastases in vivo. Prostate 2014, 74, 1251–1265. [Google Scholar] [CrossRef] [PubMed]
- Bakker, A.D.; Klein-Nulend, J. Osteoblast isolation from murine calvaria and long bones. Methods Mol. Biol. 2012, 816, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.L.; Bafico, A.; Dai, J.; Aaronson, S.A.; Keller, E.T. Prostate cancer cells promote osteoblastic bone metastases through Wnts. Cancer Res. 2005, 65, 7554–7560. [Google Scholar] [CrossRef]
- Rachner, T.D.; Thiele, S.; Gobel, A.; Browne, A.; Fuessel, S.; Erdmann, K.; Wirth, M.P.; Frohner, M.; Todenhofer, T.; Muders, M.H.; et al. High serum levels of Dickkopf-1 are associated with a poor prognosis in prostate cancer patients. BMC Cancer 2014, 14, 649. [Google Scholar] [CrossRef]
- Yamabuki, T.; Takano, A.; Hayama, S.; Ishikawa, N.; Kato, T.; Miyamoto, M.; Ito, T.; Ito, H.; Miyagi, Y.; Nakayama, H.; et al. Dikkopf-1 as a novel serologic and prognostic biomarker for lung and esophageal carcinomas. Cancer Res. 2007, 67, 2517–2525. [Google Scholar] [CrossRef]
- Kasoha, M.; Bohle, R.M.; Seibold, A.; Gerlinger, C.; Juhasz-Boss, I.; Solomayer, E.F. Dickkopf-1 (Dkk1) protein expression in breast cancer with special reference to bone metastases. Clin. Exp. Metastasis 2018, 35, 763–775. [Google Scholar] [CrossRef]
- Chu, H.Y.; Chen, Z.; Wang, L.; Zhang, Z.K.; Tan, X.; Liu, S.; Zhang, B.T.; Lu, A.; Yu, Y.; Zhang, G. Dickkopf-1: A Promising Target for Cancer Immunotherapy. Front. Immunol. 2021, 12, 658097. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Zhu, S.; Zheng, J.; Xie, Y.; Jiang, H.; Guo, J.; Wang, Y.; Peng, Z.; Wang, M.; et al. DKK1 activates noncanonical NF-kappaB signaling via IL-6-induced CKAP4 receptor in multiple myeloma. Blood Adv. 2021, 5, 3656–3667. [Google Scholar] [CrossRef] [PubMed]
- Menezes, M.E.; Devine, D.J.; Shevde, L.A.; Samant, R.S. Dickkopf1: A tumor suppressor or metastasis promoter? Int. J. Cancer 2012, 130, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.L.; Daignault, S.D.; Shah, R.B.; Pienta, K.J.; Keller, E.T. Dickkopf-1 expression increases early in prostate cancer development and decreases during progression from primary tumor to metastasis. Prostate 2008, 68, 1396–1404. [Google Scholar] [CrossRef] [PubMed]
- Clines, K.L.; Clines, G.A. DKK1 and Kremen Expression Predicts the Osteoblastic Response to Bone Metastasis. Transl. Oncol. 2018, 11, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Roudier, M.P.; Morrissey, C.; True, L.D.; Higano, C.S.; Vessella, R.L.; Ott, S.M. Histopathological assessment of prostate cancer bone osteoblastic metastases. J. Urol. 2008, 180, 1154–1160. [Google Scholar] [CrossRef]
- Bisseret, D.; Kaci, R.; Lafage-Proust, M.H.; Alison, M.; Parlier-Cuau, C.; Laredo, J.D.; Bousson, V. Periosteum: Characteristic imaging findings with emphasis on radiologic-pathologic comparisons. Skeletal Radiol. 2015, 44, 321–338. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, Z.; Yu, Y.; Chu, H.Y.; Yu, S.; Yao, S.; Zhang, G.; Zhang, B.T. Drug Discovery of DKK1 Inhibitors. Front. Pharmacol. 2022, 13, 847387. [Google Scholar] [CrossRef]
- Wise, D.R.; Schneider, J.A.; Armenia, J.; Febles, V.A.; McLaughlin, B.; Brennan, R.; Thoren, K.L.; Abida, W.; Sfanos, K.S.; De Marzo, A.M.; et al. Dickkopf-1 Can Lead to Immune Evasion in Metastatic Castration-Resistant Prostate Cancer. JCO Precis. Oncol. 2020, 4, 1167–1179. [Google Scholar] [CrossRef]
- Bell, F.W.; Klausner, J.S.; Hayden, D.W.; Feeney, D.A.; Johnston, S.D. Clinical and pathologic features of prostatic adenocarcinoma in sexually intact and castrated dogs: 31 cases (1970–1987). J. Am. Vet. Med. Assoc. 1991, 199, 1623–1630. [Google Scholar] [PubMed]
- Rosol, T.J.; Tannehill-Gregg, S.H.; LeRoy, B.E.; Mandl, S.; Contag, C.H. Animal models of bone metastasis. Cancer 2003, 97, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Bell, F.W.; Klausner, J.S.; Hayden, D.W.; Lund, E.M.; Liebenstein, B.B.; Feeney, D.A.; Johnston, S.D.; Shivers, J.L.; Ewing, C.M.; Isaacs, W.B. Evaluation of serum and seminal plasma markers in the diagnosis of canine prostatic disorders. J. Vet. Intern. Med. 1995, 9, 149–153. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Species | Forward Primer | Reverse Primer |
|---|---|---|---|
| Ubc | murine | CGTCGAGCCCAGTGTTACCACCAAGAAGG | CCCCCATCACACCCAAGAACAAGCACAAG |
| Rankl | ACACCTCACCATCAATGCT | CTTAACGTCATGTTAGAGATCTTGG | |
| Opg | AGCTGCTGAAGCTGTGGAA | TCGAGTGGCCGAGAT | |
| Mmp9 | CATTCGCGTGGATAAGGAGT | ATTTTGGAAACTCACACGCC | |
| Ctsk | CTTCCAATACGTGCAGCAGA | CCTCTGCATTTAGCTGCCTT | |
| GAPDH | canine | CCCACTCTTCCACCTTCGAC | AGCCAAATTCATTGTCATACCAGG |
| CDH1 | GCTGCTGACCTGCAAGGCGA | GGCCGGGGTATCGGGGACAT | |
| SLUG | GGCAAGGCGTTTTCCAGACCCT | GGGCAAGAAAAAGGCTTCTCCCCAG | |
| SNAIL | GTCTGTGGCACCTGCGGGAAG | GAAGGTTGGAGCGGTCGGCA | |
| TWIST1 | GGCAGGGCCGGAGACCTAGATG | TCCACGGGCCTGTCTCGCTT | |
| COX2 | AACATCCCCTTCCTGCGAAAT | TGGGTGTTGGACAGTCATCAG | |
| VEGFA | CGCAGACGTGTAAATGTTCCTG | TCCCGAAACCCTGAGGGAG | |
| DKK-1 | human | GACCATTGACAACTACCAGCC | GGGACTAGCGCAGTACTCATC |
| Femur/Tibia | Radius/Ulna | Scapula/Humerus | Vertebrae | Mandible | |
|---|---|---|---|---|---|
| Probasco (n = 7) | 7/7 | 3/7 | 3/7 | 2/7 | 6/7 |
| Probasco + DKK-1 (n = 6) | 6/6 | 5/6 | 5/6 | 4/6 | 6/6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, S.; Hoggard, N.K.; Kantake, N.; Hildreth, B.E., III; Rosol, T.J. Effects of Dickkopf-1 (DKK-1) on Prostate Cancer Growth and Bone Metastasis. Cells 2023, 12, 2695. https://doi.org/10.3390/cells12232695
Yuan S, Hoggard NK, Kantake N, Hildreth BE III, Rosol TJ. Effects of Dickkopf-1 (DKK-1) on Prostate Cancer Growth and Bone Metastasis. Cells. 2023; 12(23):2695. https://doi.org/10.3390/cells12232695
Chicago/Turabian StyleYuan, Shiyu, Nathan K. Hoggard, Noriko Kantake, Blake E. Hildreth, III, and Thomas J. Rosol. 2023. "Effects of Dickkopf-1 (DKK-1) on Prostate Cancer Growth and Bone Metastasis" Cells 12, no. 23: 2695. https://doi.org/10.3390/cells12232695
APA StyleYuan, S., Hoggard, N. K., Kantake, N., Hildreth, B. E., III, & Rosol, T. J. (2023). Effects of Dickkopf-1 (DKK-1) on Prostate Cancer Growth and Bone Metastasis. Cells, 12(23), 2695. https://doi.org/10.3390/cells12232695