
Chikungunya Virus and Its Envelope Protein E2 Induce Hyperalgesia in Mice: Inhibition by Anti-E2 Monoclonal Antibodies and by Targeting TRPV1

 , , , , , , ,
, , , , , , ,  , , and
, , and 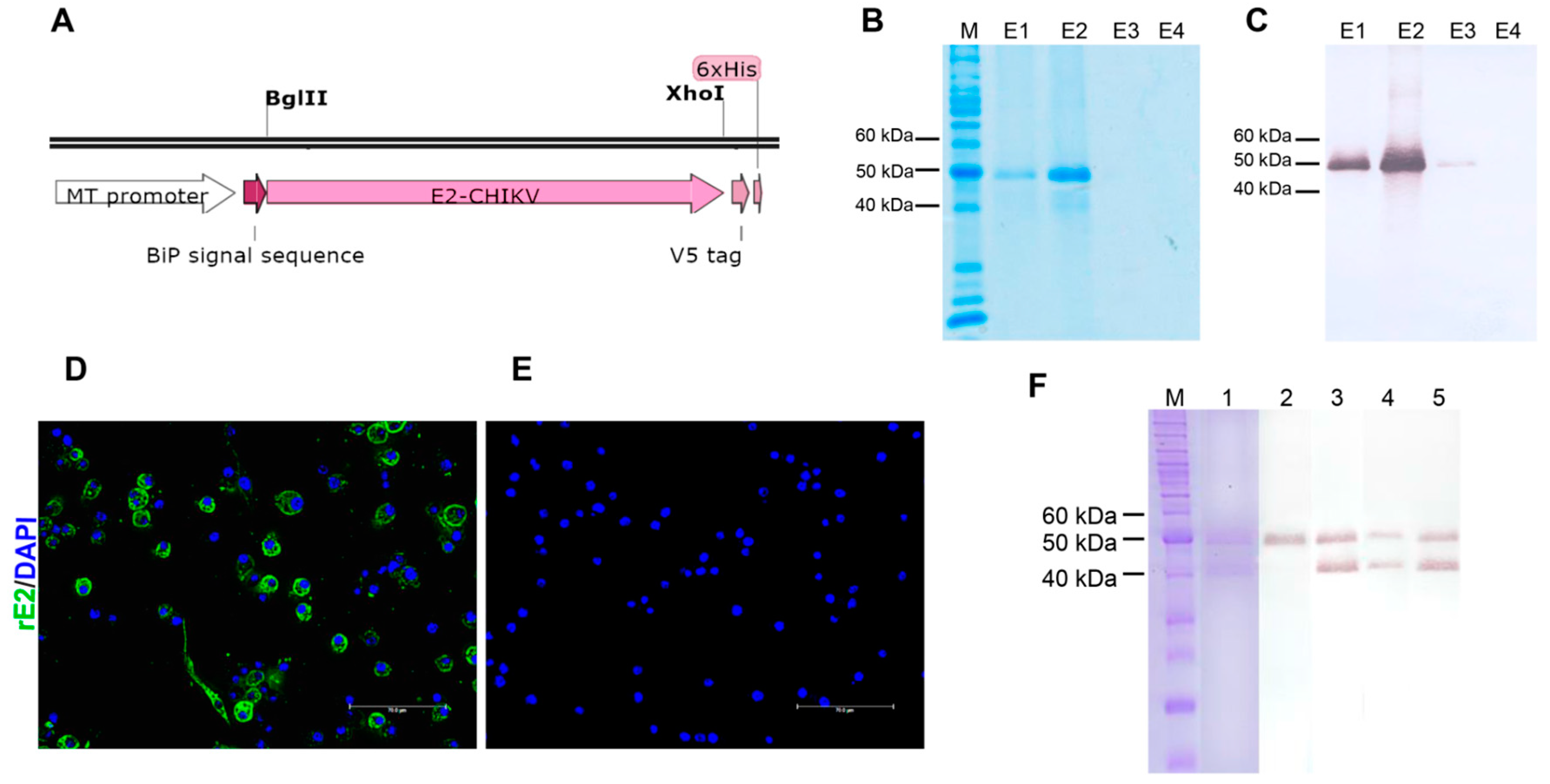{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Inactivation of Chikungunya Virus
2.3. Cloning, Expression, and Purification of CHIKV rE2 Protein
2.4. Anti-rE2 Antibodies Synthesis
2.5. Experimental Protocols
2.6. Mechanical Hyperalgesia
2.7. Thermal Hyperalgesia
2.8. Calcium Imaging
2.9. Statistical Analysis
3. Results
3.1. Expression and Purification of CHIKV rE2
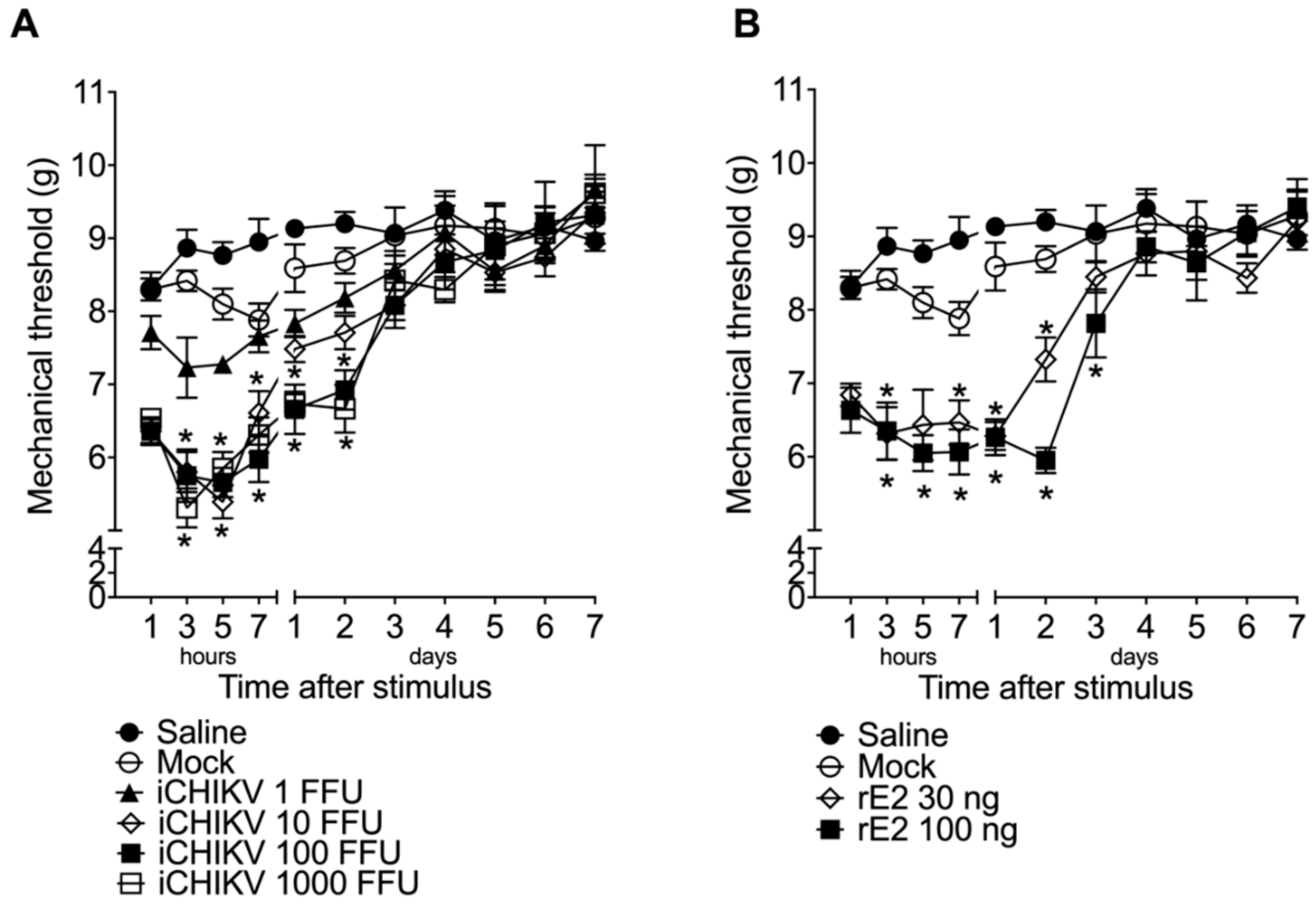
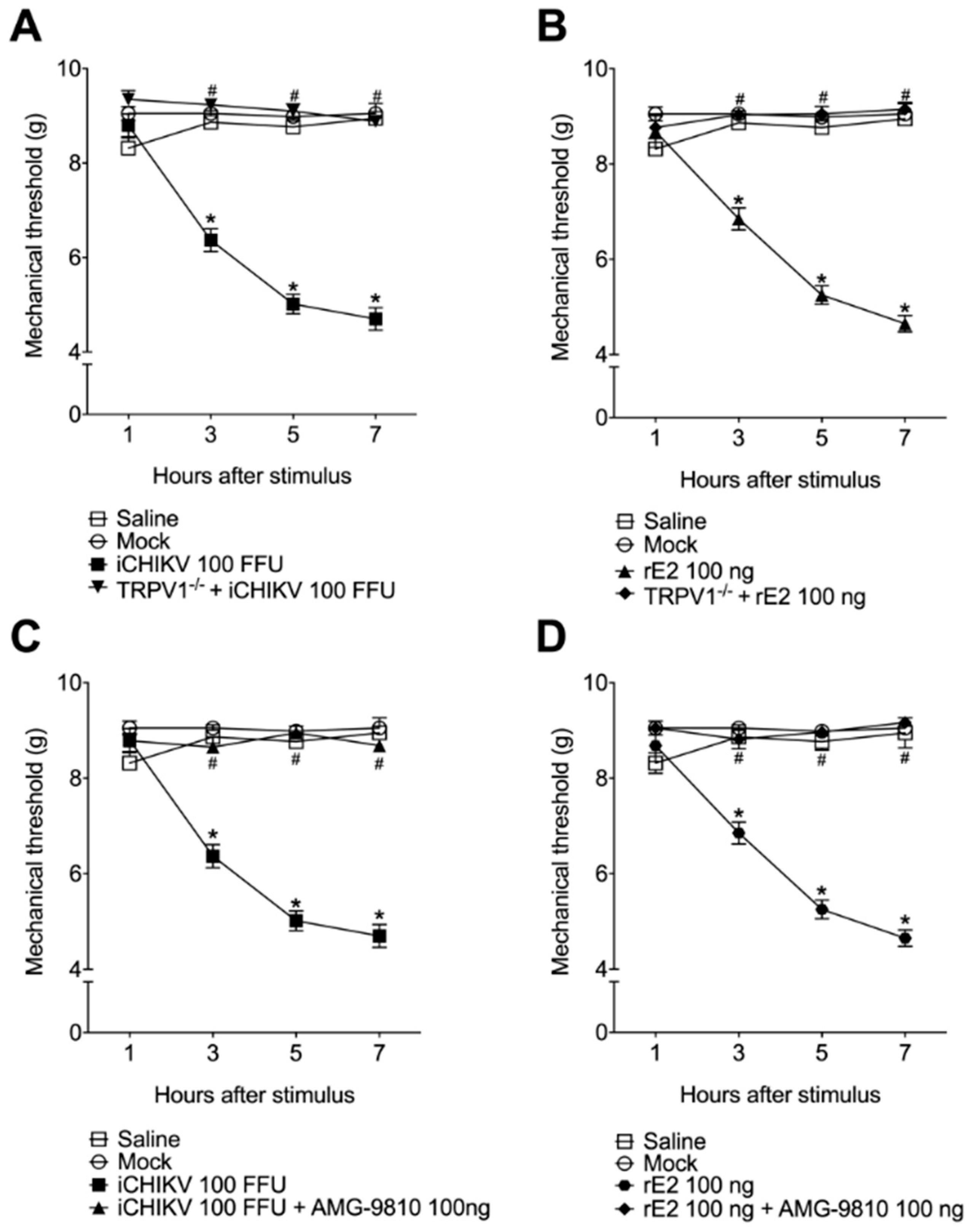
3.2. iCHIKV and rE2 Protein Induce Mechanical Hyperalgesia in Mice
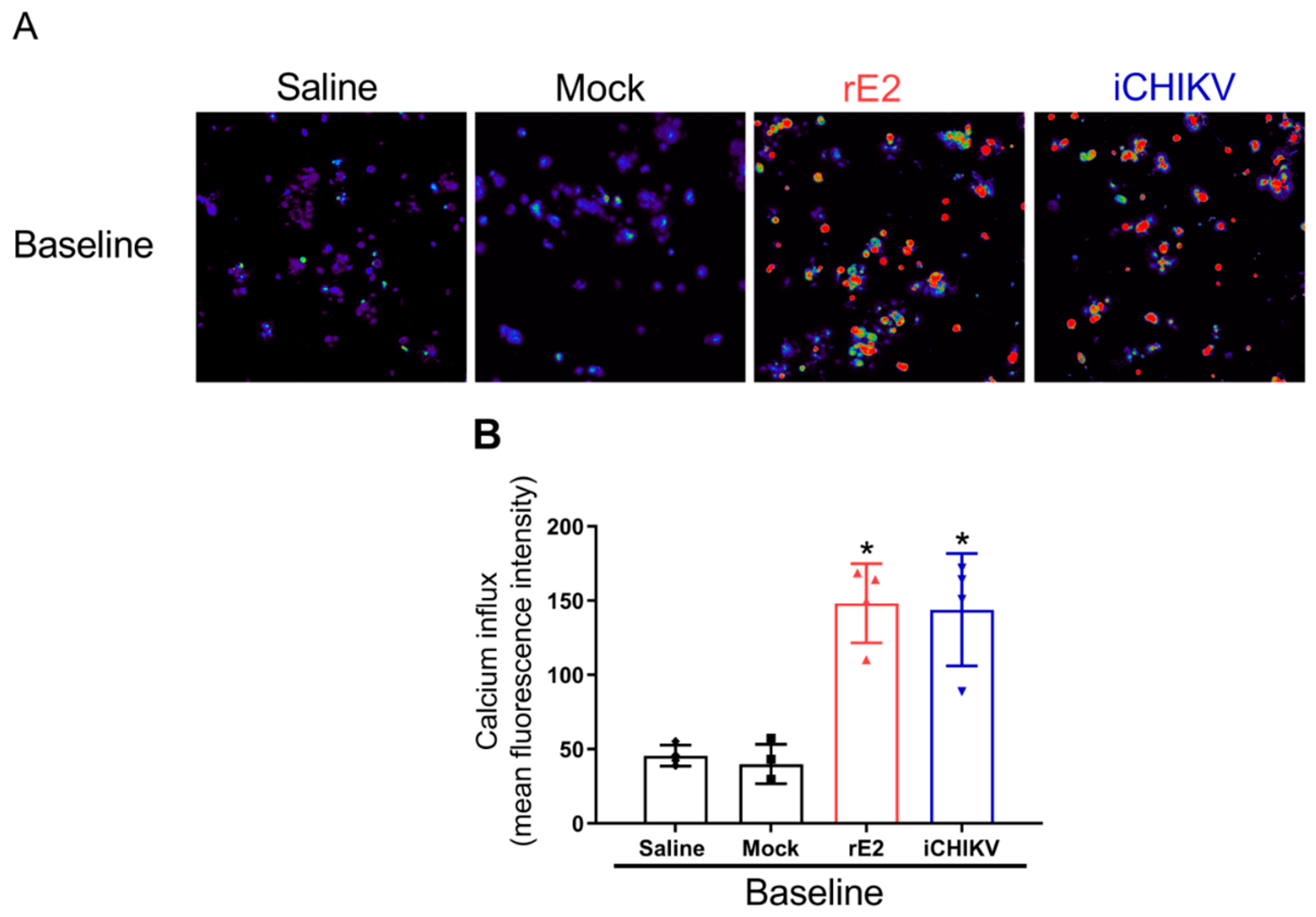
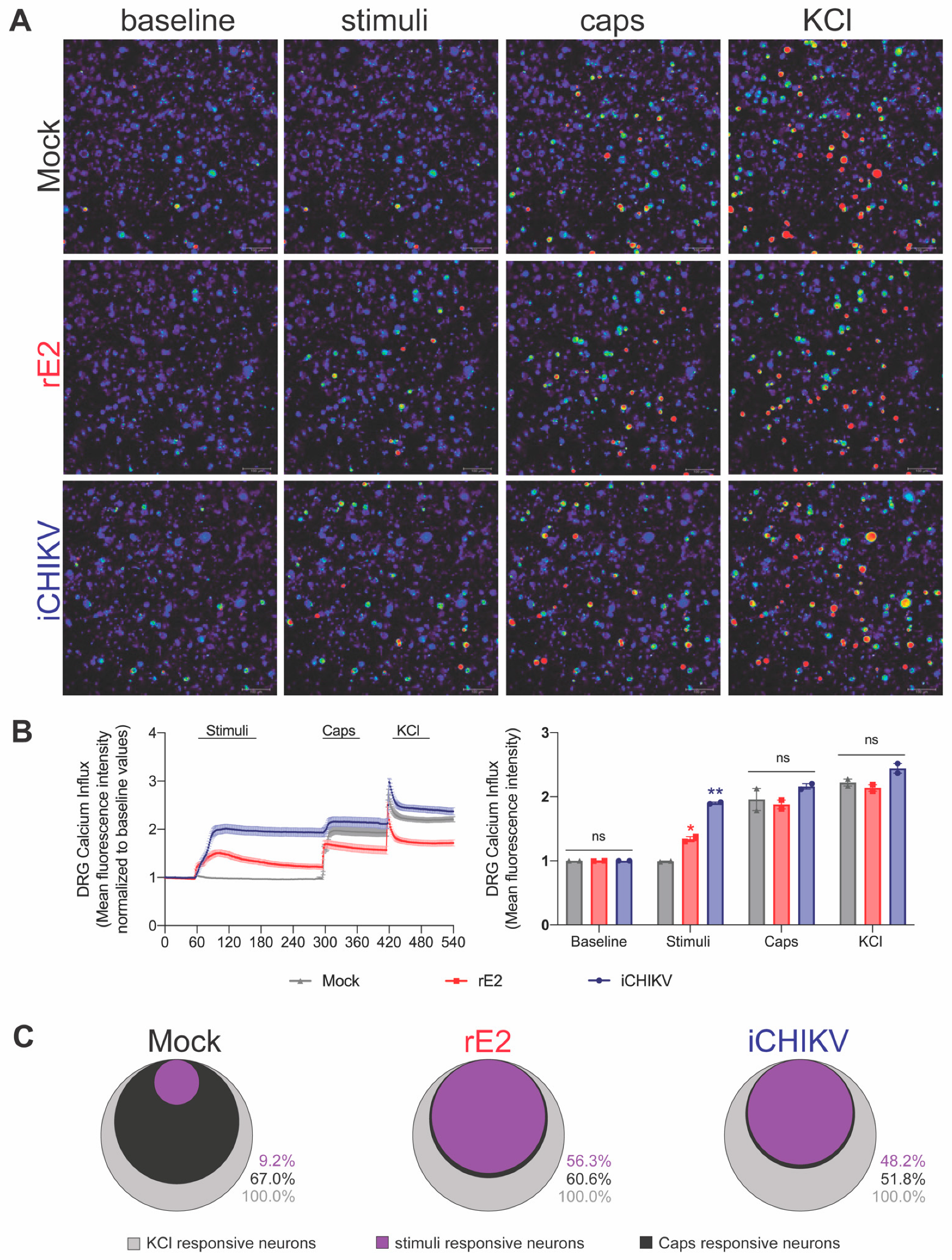
3.3. DRG Neurons from iCHIKV and rE2-Stimulated Mice Presented Higher Baseline Levels of Calcium
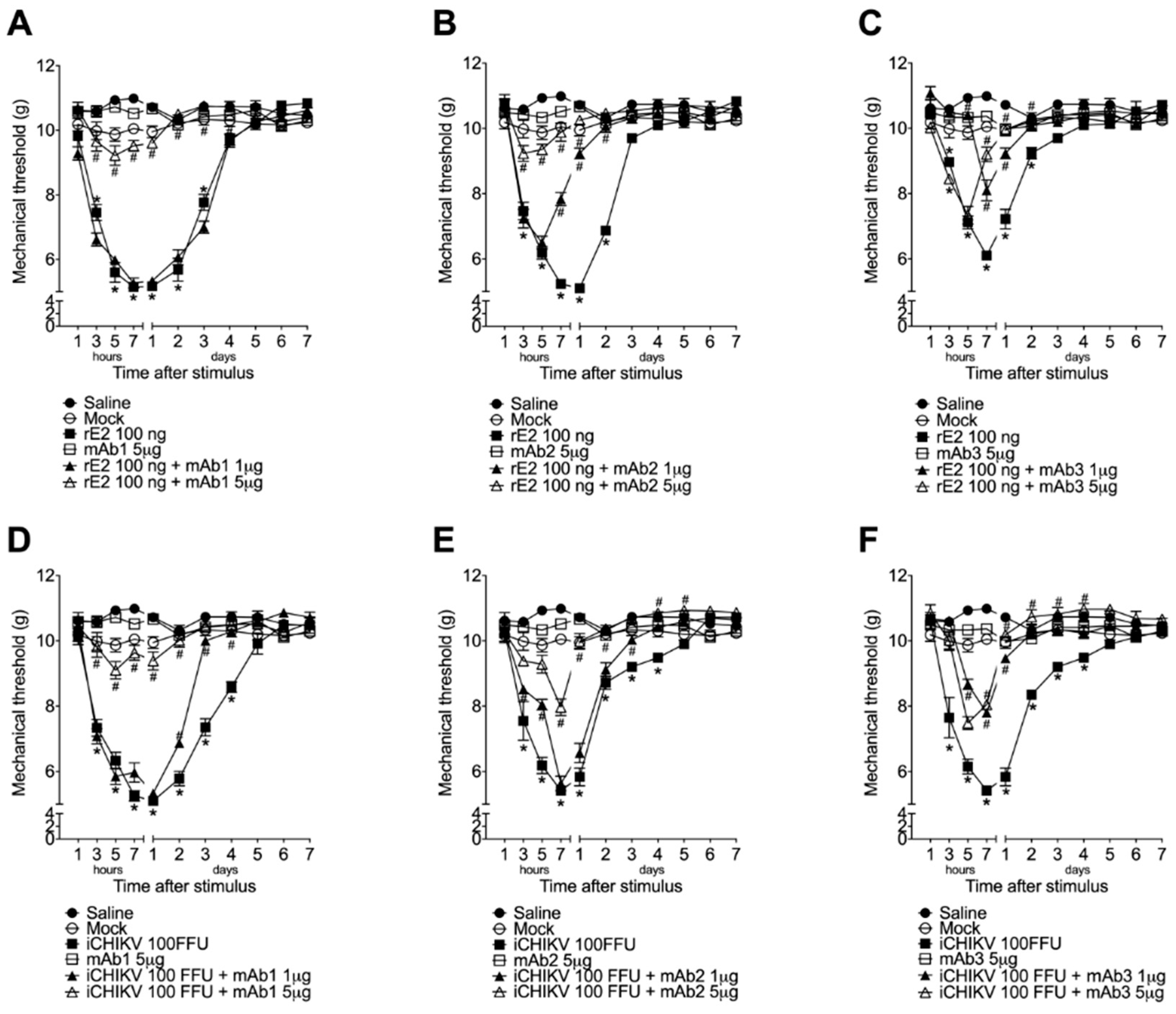
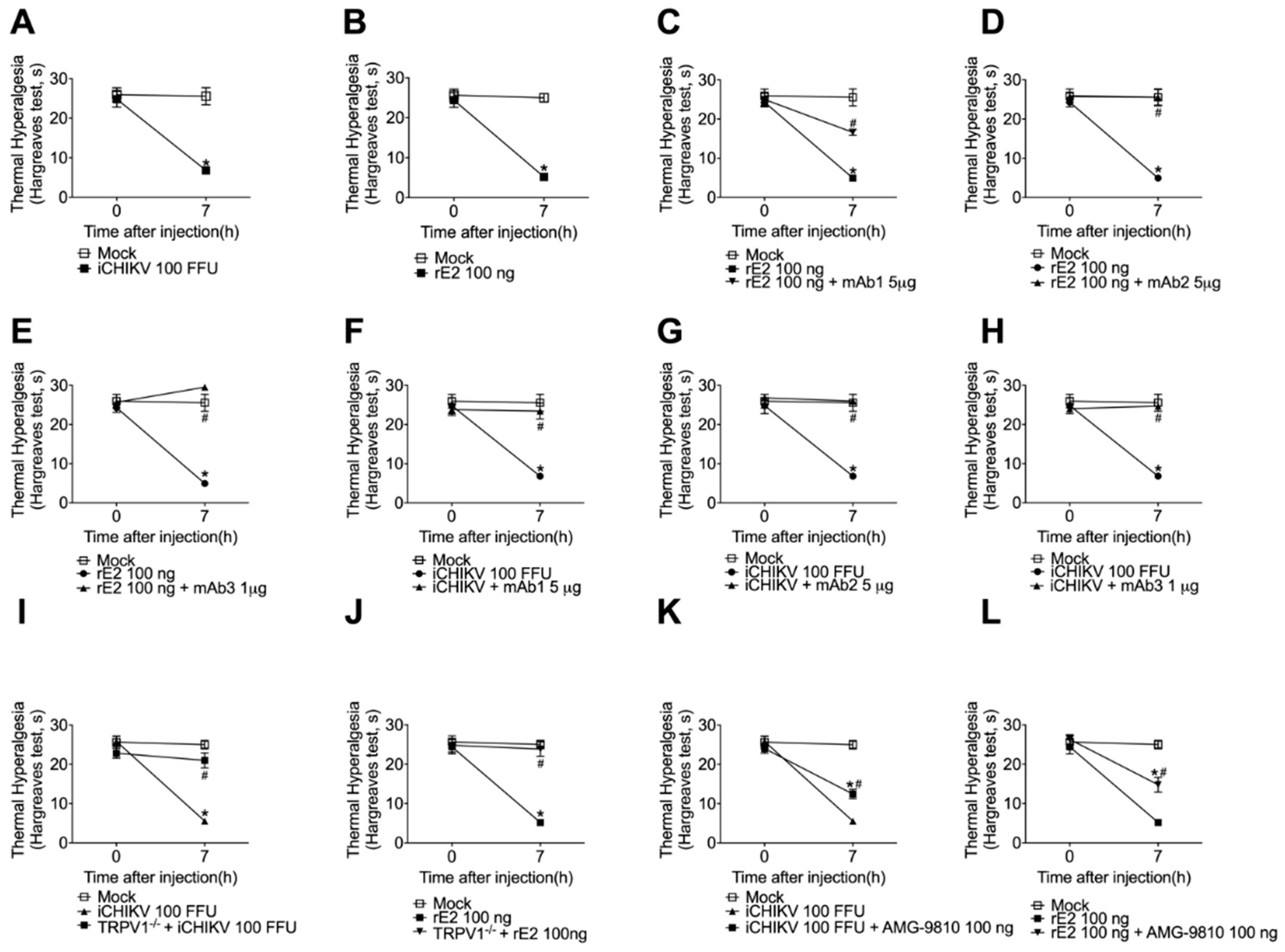
3.4. Monoclonal Antibodies Targeting rE2 Reduce rE2 and iCHIKV-Induced Mechanical Hyperalgesia
3.5. The Mechanical Hyperalgesia Caused by iCHIKV and rE2 Protein Was Inhibited by the Absence or Blockade of TRPV1 Receptor
3.6. iCHIKV and rE2 Protein Induce Thermal Hyperalgesia in Mice: Inhibition by Anti-rE2 Monoclonal Antibodies and by Targeting TRPV1 Receptor
3.7. iCHIKV and rE2 Protein Activate TRPV1+ DRG Nociceptive Neurons
4. Discussion
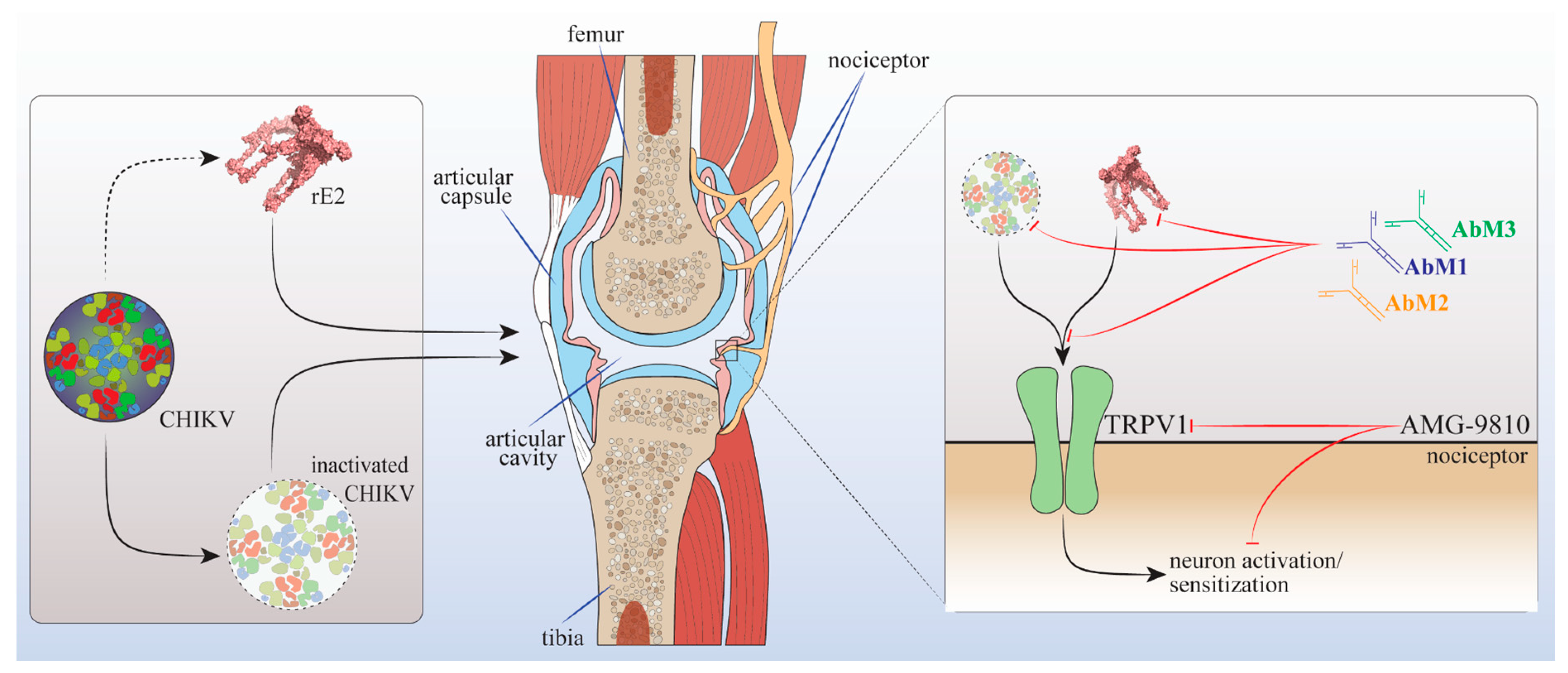
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caglioti, C.; Lalle, E.; Castilletti, C.; Carletti, F.; Capobianchi, M.R.; Bordi, L. Chikungunya virus infection: An overview. New Microbiol. 2013, 36, 211–227. [Google Scholar]
- Her, Z.; Kam, Y.W.; Lin, R.T.; Ng, L.F. Chikungunya: A bending reality. Microbes. Infect. 2009, 11, 1165–1176. [Google Scholar] [CrossRef]
- Robinson, M.C. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952–1953. I. Clinical features. Trans. R Soc. Trop. Med. Hyg. 1955, 49, 28–32. [Google Scholar] [CrossRef]
- Burt, F.J.; Rolph, M.S.; Rulli, N.E.; Mahalingam, S.; Heise, M.T. Chikungunya: A re-emerging virus. Lancet 2012, 379, 662–671. [Google Scholar] [CrossRef]
- Gerardin, P.; Fianu, A.; Malvy, D.; Mussard, C.; Boussaid, K.; Rollot, O.; Michault, A.; Gauzere, B.A.; Breart, G.; Favier, F. Perceived morbidity and community burden after a Chikungunya outbreak: The TELECHIK survey, a population-based cohort study. BMC Med. 2011, 9, 5. [Google Scholar] [CrossRef]
- Labeaud, A.D.; Bashir, F.; King, C.H. Measuring the burden of arboviral diseases: The spectrum of morbidity and mortality from four prevalent infections. Popul. Health Metr. 2011, 9, 1. [Google Scholar] [CrossRef]
- Weaver, S.C.; Osorio, J.E.; Livengood, J.A.; Chen, R.; Stinchcomb, D.T. Chikungunya virus and prospects for a vaccine. Expert Rev. Vaccines 2012, 11, 1087–1101. [Google Scholar] [CrossRef]
- Lam, S.K.; Chua, K.B.; Hooi, P.S.; Rahimah, M.A.; Kumari, S.; Tharmaratnam, M.; Chuah, S.K.; Smith, D.W.; Sampson, I.A. Chikungunya infection--an emerging disease in Malaysia. Southeast Asian J. Trop. Med. Public Health 2001, 32, 447–451. [Google Scholar]
- Borgherini, G.; Poubeau, P.; Jossaume, A.; Gouix, A.; Cotte, L.; Michault, A.; Arvin-Berod, C.; Paganin, F. Persistent arthralgia associated with chikungunya virus: A study of 88 adult patients on reunion island. Clin. Infect. Dis. 2008, 47, 469–475. [Google Scholar] [CrossRef]
- Ross, R.W. The Newala epidemic. III. The virus: Isolation, pathogenic properties and relationship to the epidemic. J. Hyg. 1956, 54, 177–191. [Google Scholar] [CrossRef]
- Staples, J.E.; Breiman, R.F.; Powers, A.M. Chikungunya fever: An epidemiological review of a re-emerging infectious disease. Clin. Infect. Dis. 2009, 49, 942–948. [Google Scholar] [CrossRef]
- Simon, F.; Javelle, E.; Oliver, M.; Leparc-Goffart, I.; Marimoutou, C. Chikungunya virus infection. Curr. Infect. Dis. Rep. 2011, 13, 218–228. [Google Scholar] [CrossRef] [Green Version]
- Pialoux, G.; Gauzere, B.A.; Jaureguiberry, S.; Strobel, M. Chikungunya, an epidemic arbovirosis. Lancet Infect. Dis. 2007, 7, 319–327. [Google Scholar] [CrossRef]
- Hoarau, J.J.; Jaffar Bandjee, M.C.; Krejbich Trotot, P.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T.; et al. Persistent chronic inflammation and infection by Chikungunya arthritogenic alphavirus in spite of a robust host immune response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef]
- Schilte, C.; Staikowsky, F.; Couderc, T.; Madec, Y.; Carpentier, F.; Kassab, S.; Albert, M.L.; Lecuit, M.; Michault, A. Chikungunya virus-associated long-term arthralgia: A 36-month prospective longitudinal study. PLoS Negl. Trop. Dis. 2013, 7, e2137. [Google Scholar] [CrossRef]
- Simon, F.; Parola, P.; Grandadam, M.; Fourcade, S.; Oliver, M.; Brouqui, P.; Hance, P.; Kraemer, P.; Mohamed, A.A.; de Lamballerie, X.; et al. Chikungunya infection: An emerging rheumatism among travelers returned from Indian Ocean islands. Report of 47 cases. Medicine 2007, 86, 123–137. [Google Scholar] [CrossRef]
- Sissoko, D.; Malvy, D.; Ezzedine, K.; Renault, P.; Moscetti, F.; Ledrans, M.; Pierre, V. Post-epidemic Chikungunya disease on Reunion Island: Course of rheumatic manifestations and associated factors over a 15-month period. PLoS Negl. Trop. Dis. 2009, 3, e389. [Google Scholar] [CrossRef]
- Staikowsky, F.; Le Roux, K.; Schuffenecker, I.; Laurent, P.; Grivard, P.; Develay, A.; Michault, A. Retrospective survey of Chikungunya disease in Reunion Island hospital staff. Epidemiol. Infect. 2008, 136, 196–206. [Google Scholar] [CrossRef]
- Selvarajah, S.; Sexton, N.R.; Kahle, K.M.; Fong, R.H.; Mattia, K.A.; Gardner, J.; Lu, K.; Liss, N.M.; Salvador, B.; Tucker, D.F.; et al. A neutralizing monoclonal antibody targeting the acid-sensitive region in chikungunya virus E2 protects from disease. PLoS Negl. Trop. Dis. 2013, 7, e2423. [Google Scholar] [CrossRef]
- Broeckel, R.; Fox, J.M.; Haese, N.; Kreklywich, C.N.; Sukulpovi-Petty, S.; Legasse, A.; Smith, P.P.; Denton, M.; Corvey, C.; Krishnan, S.; et al. Therapeutic administration of a recombinant human monoclonal antibody reduces the severity of chikungunya virus disease in rhesus macaques. PLoS Negl. Trop. Dis. 2017, 11, e0005637. [Google Scholar] [CrossRef]
- Edelman, R.; Tacket, C.O.; Wasserman, S.S.; Bodison, S.A.; Perry, J.G.; Mangiafico, J.A. Phase II safety and immunogenicity study of live chikungunya virus vaccine TSI-GSD-218. Am. J. Trop. Med. Hyg. 2000, 62, 681–685. [Google Scholar] [CrossRef]
- Parashar, D.; Cherian, S. Antiviral perspectives for chikungunya virus. BioMed Res. Int. 2014, 2014, 631642. [Google Scholar] [CrossRef] [Green Version]
- Partidos, C.D.; Weger, J.; Brewoo, J.; Seymour, R.; Borland, E.M.; Ledermann, J.P.; Powers, A.M.; Weaver, S.C.; Stinchcomb, D.T.; Osorio, J.E. Probing the attenuation and protective efficacy of a candidate chikungunya virus vaccine in mice with compromised interferon (IFN) signaling. Vaccine 2011, 29, 3067–3073. [Google Scholar] [CrossRef]
- van den Doel, P.; Volz, A.; Roose, J.M.; Sewbalaksing, V.D.; Pijlman, G.P.; van Middelkoop, I.; Duiverman, V.; van de Wetering, E.; Sutter, G.; Osterhaus, A.D.; et al. Recombinant modified vaccinia virus Ankara expressing glycoprotein E2 of Chikungunya virus protects AG129 mice against lethal challenge. PLoS Negl. Trop. Dis. 2014, 8, e3101. [Google Scholar] [CrossRef]
- Teo, T.H.; Chan, Y.H.; Lee, W.W.; Lum, F.M.; Amrun, S.N.; Her, Z.; Rajarethinam, R.; Merits, A.; Rotzschke, O.; Renia, L.; et al. Fingolimod treatment abrogates chikungunya virus-induced arthralgia. Sci. Transl. Med. 2017, 9, eaal1333. [Google Scholar] [CrossRef]
- Warter, L.; Lee, C.Y.; Thiagarajan, R.; Grandadam, M.; Lebecque, S.; Lin, R.T.; Bertin-Maghit, S.; Ng, L.F.; Abastado, J.P.; Despres, P.; et al. Chikungunya virus envelope-specific human monoclonal antibodies with broad neutralization potency. J. Immunol. 2011, 186, 3258–3264. [Google Scholar] [CrossRef]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Despres, P.; Frenkiel, M.P.; Deubel, V. Differences between cell membrane fusion activities of two dengue type-1 isolates reflect modifications of viral structure. Virology 1993, 196, 209–219. [Google Scholar] [CrossRef]
- Cho, B.; Jeon, B.Y.; Kim, J.; Noh, J.; Kim, J.; Park, M.; Park, S. Expression and evaluation of Chikungunya virus E1 and E2 envelope proteins for serodiagnosis of Chikungunya virus infection. Yonsei Med. J. 2008, 49, 828–835. [Google Scholar] [CrossRef]
- Zanluca, C.; Mazzarotto, G.A.; Bordignon, J.; Duarte Dos Santos, C.N. Development, characterization and application of monoclonal antibodies against Brazilian Dengue virus isolates. PLoS ONE 2014, 9, e110620. [Google Scholar] [CrossRef]
- Guerrero, A.T.; Verri, W.A., Jr.; Cunha, T.M.; Silva, T.A.; Rocha, F.A.; Ferreira, S.H.; Cunha, F.Q.; Parada, C.A. Hypernociception elicited by tibio-tarsal joint flexion in mice: A novel experimental arthritis model for pharmacological screening. Pharmacol. Biochem. Behav. 2006, 84, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Manchope, M.F.; Artero, N.A.; Fattori, V.; Mizokami, S.S.; Pitol, D.L.; Issa, J.P.M.; Fukada, S.Y.; Cunha, T.M.; Alves-Filho, J.C.; Cunha, F.Q.; et al. Naringenin mitigates titanium dioxide (TiO2)-induced chronic arthritis in mice: Role of oxidative stress, cytokines, and NFkappaB. Inflamm. Res. 2018, 67, 997–1012. [Google Scholar] [CrossRef]
- Hargreaves, K.; Dubner, R.; Brown, F.; Flores, C.; Joris, J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain 1988, 32, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Fattori, V.; Pinho-Ribeiro, F.A.; Staurengo-Ferrari, L.; Borghi, S.M.; Rossaneis, A.C.; Casagrande, R.; Verri, W.A., Jr. The specialised pro-resolving lipid mediator maresin 1 reduces inflammatory pain with a long-lasting analgesic effect. Br. J. Pharmacol. 2019, 176, 1728–1744. [Google Scholar] [CrossRef] [PubMed]
- Blake, K.J.; Baral, P.; Voisin, T.; Lubkin, A.; Pinho-Ribeiro, F.A.; Adams, K.L.; Roberson, D.P.; Ma, Y.C.; Otto, M.; Woolf, C.J.; et al. Staphylococcus aureus produces pain through pore-forming toxins and neuronal TRPV1 that is silenced by QX-314. Nat. Commun. 2018, 9, 37. [Google Scholar] [CrossRef]
- Donate-Macian, P.; Jungfleisch, J.; Perez-Vilaro, G.; Rubio-Moscardo, F.; Peralvarez-Marin, A.; Diez, J.; Valverde, M.A. The TRPV4 channel links calcium influx to DDX3X activity and viral infectivity. Nat. Commun. 2018, 9, 2307. [Google Scholar] [CrossRef]
- Karlas, A.; Berre, S.; Couderc, T.; Varjak, M.; Braun, P.; Meyer, M.; Gangneux, N.; Karo-Astover, L.; Weege, F.; Raftery, M.; et al. A human genome-wide loss-of-function screen identifies effective chikungunya antiviral drugs. Nat. Commun. 2016, 7, 11320. [Google Scholar] [CrossRef]
- Goh, L.Y.; Hobson-Peters, J.; Prow, N.A.; Gardner, J.; Bielefeldt-Ohmann, H.; Pyke, A.T.; Suhrbier, A.; Hall, R.A. Neutralizing monoclonal antibodies to the E2 protein of chikungunya virus protects against disease in a mouse model. Clin. Immunol. 2013, 149, 487–497. [Google Scholar] [CrossRef]
- Suhrbier, A.; Jaffar-Bandjee, M.C.; Gasque, P. Arthritogenic alphaviruses--An overview. Nat. Rev. Rheumatol. 2012, 8, 420–429. [Google Scholar] [CrossRef]
- Kielian, M.; Chanel-Vos, C.; Liao, M. Alphavirus Entry and Membrane Fusion. Viruses 2010, 2, 796–825. [Google Scholar] [CrossRef]
- Berta, T.; Qadri, Y.; Tan, P.H.; Ji, R.R. Targeting dorsal root ganglia and primary sensory neurons for the treatment of chronic pain. Expert Opin. Ther. Targets 2017, 21, 695–703. [Google Scholar] [CrossRef]
- Choi, S.I.; Lim, J.Y.; Yoo, S.; Kim, H.; Hwang, S.W. Emerging Role of Spinal Cord TRPV1 in Pain Exacerbation. Neural Plast. 2016, 2016, 5954890. [Google Scholar] [CrossRef] [Green Version]
- Tsagareli, M.G.; Nozadze, I.; Tsiklauri, N.; Carstens, M.I.; Gurtskaia, G.; Carstens, E. Thermal Hyperalgesia and Mechanical Allodynia Elicited by Histamine and Non-histaminergic Itch Mediators: Respective Involvement of TRPV1 and TRPA1. Neuroscience 2020, 449, 35–45. [Google Scholar] [CrossRef]
- Warwick, C.A.; Shutov, L.P.; Shepherd, A.J.; Mohapatra, D.P.; Usachev, Y.M. Mechanisms underlying mechanical sensitization induced by complement C5a: The roles of macrophages, TRPV1, and calcitonin gene-related peptide receptors. Pain 2019, 160, 702–711. [Google Scholar] [CrossRef]
- Barton, N.J.; McQueen, D.S.; Thomson, D.; Gauldie, S.D.; Wilson, A.W.; Salter, D.M.; Chessell, I.P. Attenuation of experimental arthritis in TRPV1R knockout mice. Exp. Mol. Pathol. 2006, 81, 166–170. [Google Scholar] [CrossRef]
- Caterina, M.J.; Rosen, T.A.; Tominaga, M.; Brake, A.J.; Julius, D. A capsaicin-receptor homologue with a high threshold for noxious heat. Nature 1999, 398, 436–441. [Google Scholar] [CrossRef]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhe, V.; Plee-Gautier, E.; Carre, J.L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in cutaneous neurogenic and chronic inflammation: Pro-inflammatory response induced by their activation and their sensitization. Protein Cell 2017, 8, 644–661. [Google Scholar] [CrossRef]
- Gavva, N.R.; Treanor, J.J.; Garami, A.; Fang, L.; Surapaneni, S.; Akrami, A.; Alvarez, F.; Bak, A.; Darling, M.; Gore, A.; et al. Pharmacological blockade of the vanilloid receptor TRPV1 elicits marked hyperthermia in humans. Pain 2008, 136, 202–210. [Google Scholar] [CrossRef]
- Liao, H.Y.; Hsieh, C.L.; Huang, C.P.; Lin, Y.W. Electroacupuncture Attenuates CFA-induced Inflammatory Pain by suppressing Nav1.8 through S100B, TRPV1, Opioid, and Adenosine Pathways in Mice. Sci. Rep. 2017, 7, 42531. [Google Scholar] [CrossRef]
- Tekus, V.; Bolcskei, K.; Kis-Varga, A.; Dezsi, L.; Szentirmay, E.; Visegrady, A.; Horvath, C.; Szolcsanyi, J.; Petho, G. Effect of transient receptor potential vanilloid 1 (TRPV1) receptor antagonist compounds SB705498, BCTC and AMG9810 in rat models of thermal hyperalgesia measured with an increasing-temperature water bath. Eur. J. Pharmacol. 2010, 641, 135–141. [Google Scholar] [CrossRef]
- Moreira, T.P.; de Sousa, C.D.F.; Costa, V.R.M.; Queiroz-Junior, C.M.; Santos, F.M.; Bonilha, C.S.; Esper, L.M.; Nogueira, M.L.; Cunha, T.M.; Teixeira, M.M.; et al. Tumor necrosis factor plays a deleterious role in the pathogenesis of chikungunya virus infection. Immunology, 2022; Early View. [Google Scholar] [CrossRef] [PubMed]
- Verri, W.A., Jr.; Cunha, T.M.; Parada, C.A.; Poole, S.; Cunha, F.Q.; Ferreira, S.H. Hypernociceptive role of cytokines and chemokines: Targets for analgesic drug development? Pharmacol. Ther. 2006, 112, 116–138. [Google Scholar] [CrossRef] [PubMed]
- Hibl, B.M.; Dailey Garnes, N.J.M.; Kneubehl, A.R.; Vogt, M.B.; Spencer Clinton, J.L.; Rico-Hesse, R.R. Mosquito-bite infection of humanized mice with chikungunya virus produces systemic disease with long-term effects. PLoS Negl. Trop. Dis. 2021, 15, e0009427. [Google Scholar] [CrossRef]
- Schrenk-Siemens, K.; Pohle, J.; Rostock, C.; Abd El Hay, M.; Lam, R.M.; Szczot, M.; Lu, S.; Chesler, A.T.; Siemens, J. Human Stem Cell-Derived TRPV1-Positive Sensory Neurons: A New Tool to Study Mechanisms of Sensitization. Cells 2022, 11, 2905. [Google Scholar] [CrossRef] [PubMed]
- Katz, B.; Zaguri, R.; Edvardson, S.; Maayan, C.; Elpeleg, O.; Lev, S.; Davidson, E.; Peters, M.; Kfir-Erenfeld, S.; Berger, E.; et al. Nociception and pain in humans lacking functional TRPV1 channel. J. Clin. Investig. 2022, 133, e153558. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segato-Vendrameto, C.Z.; Zanluca, C.; Zucoloto, A.Z.; Zaninelli, T.H.; Bertozzi, M.M.; Saraiva-Santos, T.; Ferraz, C.R.; Staurengo-Ferrari, L.; Badaro-Garcia, S.; Manchope, M.F.; et al. Chikungunya Virus and Its Envelope Protein E2 Induce Hyperalgesia in Mice: Inhibition by Anti-E2 Monoclonal Antibodies and by Targeting TRPV1. Cells 2023, 12, 556. https://doi.org/10.3390/cells12040556
Segato-Vendrameto CZ, Zanluca C, Zucoloto AZ, Zaninelli TH, Bertozzi MM, Saraiva-Santos T, Ferraz CR, Staurengo-Ferrari L, Badaro-Garcia S, Manchope MF, et al. Chikungunya Virus and Its Envelope Protein E2 Induce Hyperalgesia in Mice: Inhibition by Anti-E2 Monoclonal Antibodies and by Targeting TRPV1. Cells. 2023; 12(4):556. https://doi.org/10.3390/cells12040556
Chicago/Turabian StyleSegato-Vendrameto, Carina Z., Camila Zanluca, Amanda Z. Zucoloto, Tiago H. Zaninelli, Mariana M. Bertozzi, Telma Saraiva-Santos, Camila R. Ferraz, Larissa Staurengo-Ferrari, Stephanie Badaro-Garcia, Marília F. Manchope, and et al. 2023. "Chikungunya Virus and Its Envelope Protein E2 Induce Hyperalgesia in Mice: Inhibition by Anti-E2 Monoclonal Antibodies and by Targeting TRPV1" Cells 12, no. 4: 556. https://doi.org/10.3390/cells12040556
APA StyleSegato-Vendrameto, C. Z., Zanluca, C., Zucoloto, A. Z., Zaninelli, T. H., Bertozzi, M. M., Saraiva-Santos, T., Ferraz, C. R., Staurengo-Ferrari, L., Badaro-Garcia, S., Manchope, M. F., Dionisio, A. M., Pinho-Ribeiro, F. A., Borghi, S. M., Mosimann, A. L. P., Casagrande, R., Bordignon, J., Fattori, V., Santos, C. N. D. d., & Verri, W. A. (2023). Chikungunya Virus and Its Envelope Protein E2 Induce Hyperalgesia in Mice: Inhibition by Anti-E2 Monoclonal Antibodies and by Targeting TRPV1. Cells, 12(4), 556. https://doi.org/10.3390/cells12040556