



Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer



Abstract
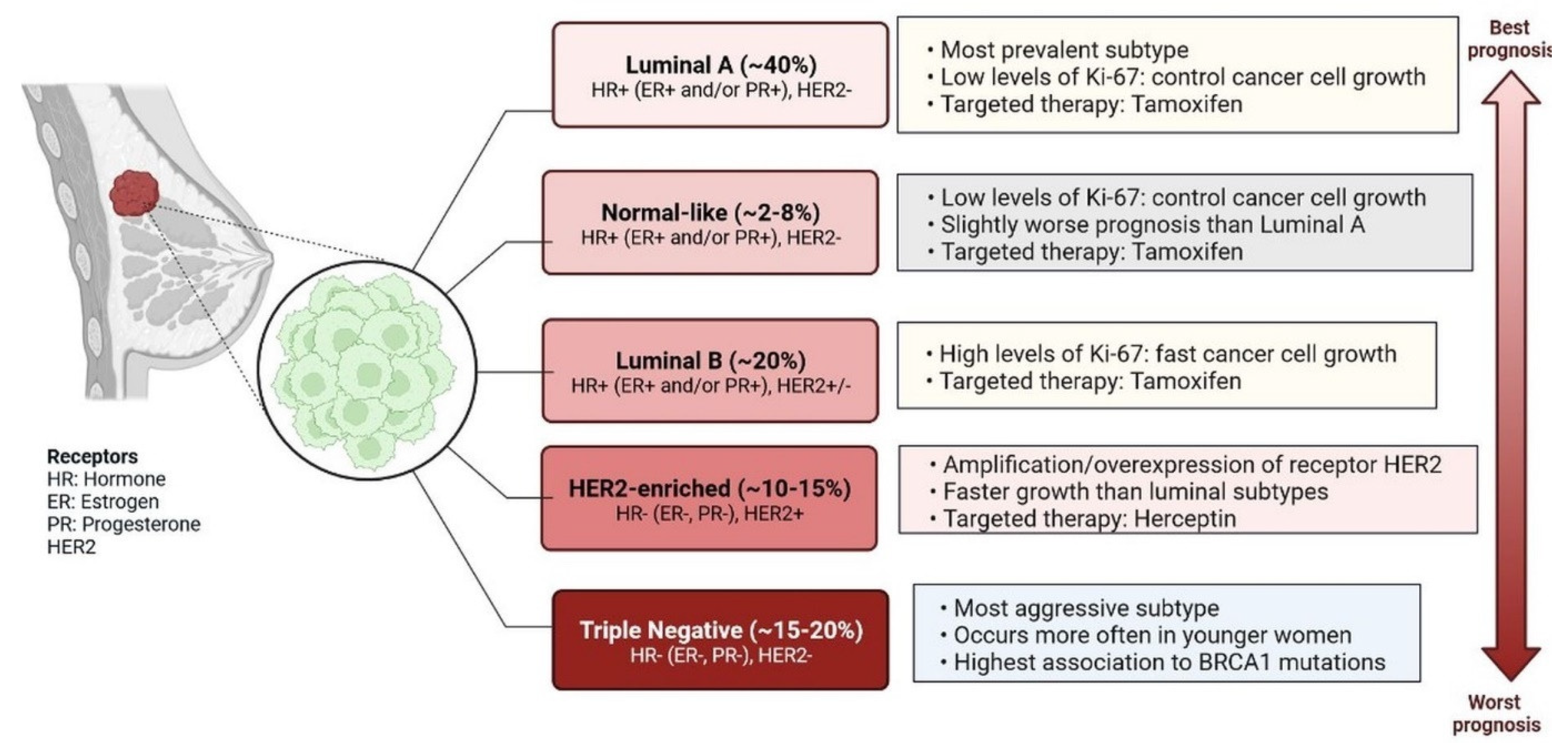
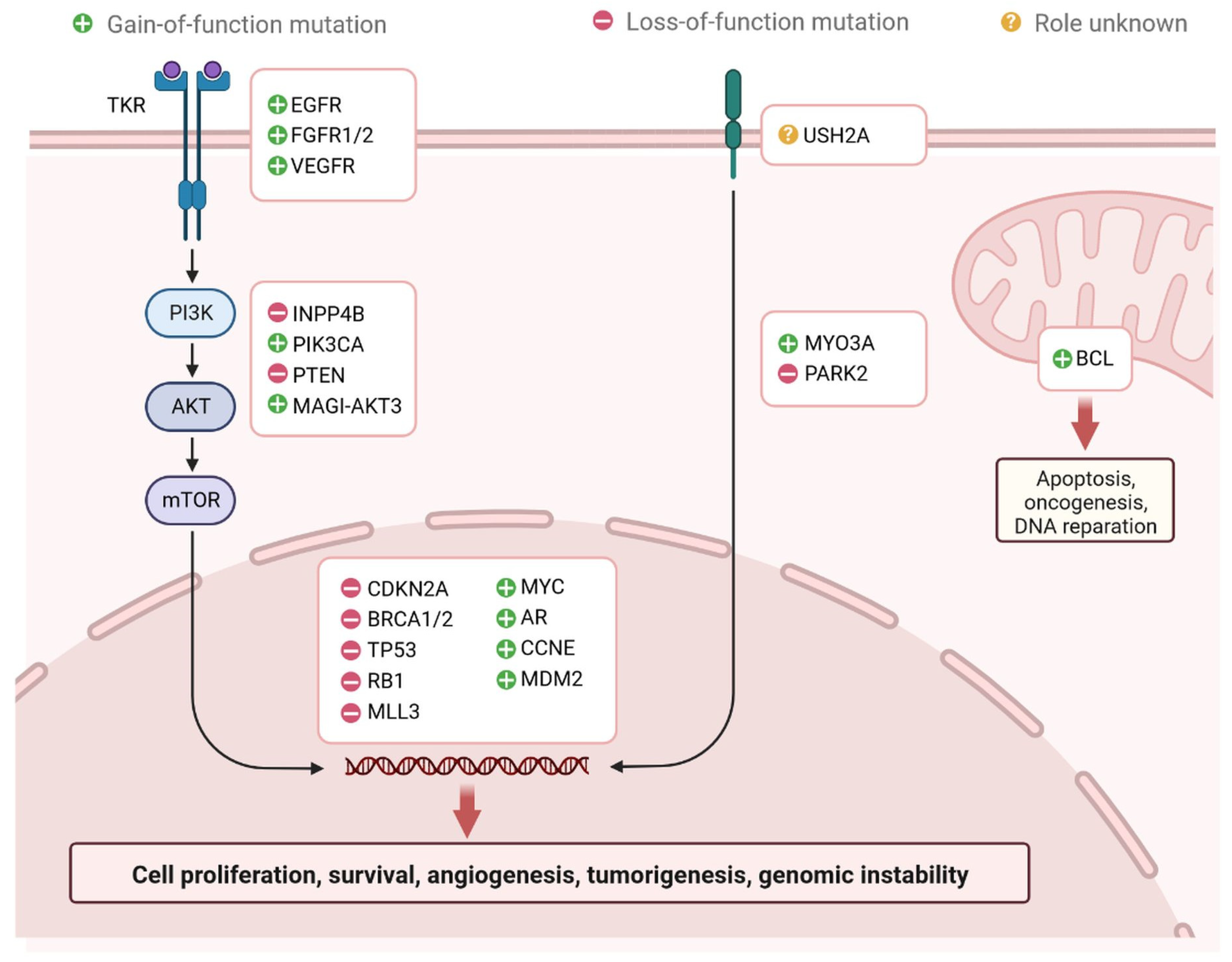
:1. Introduction
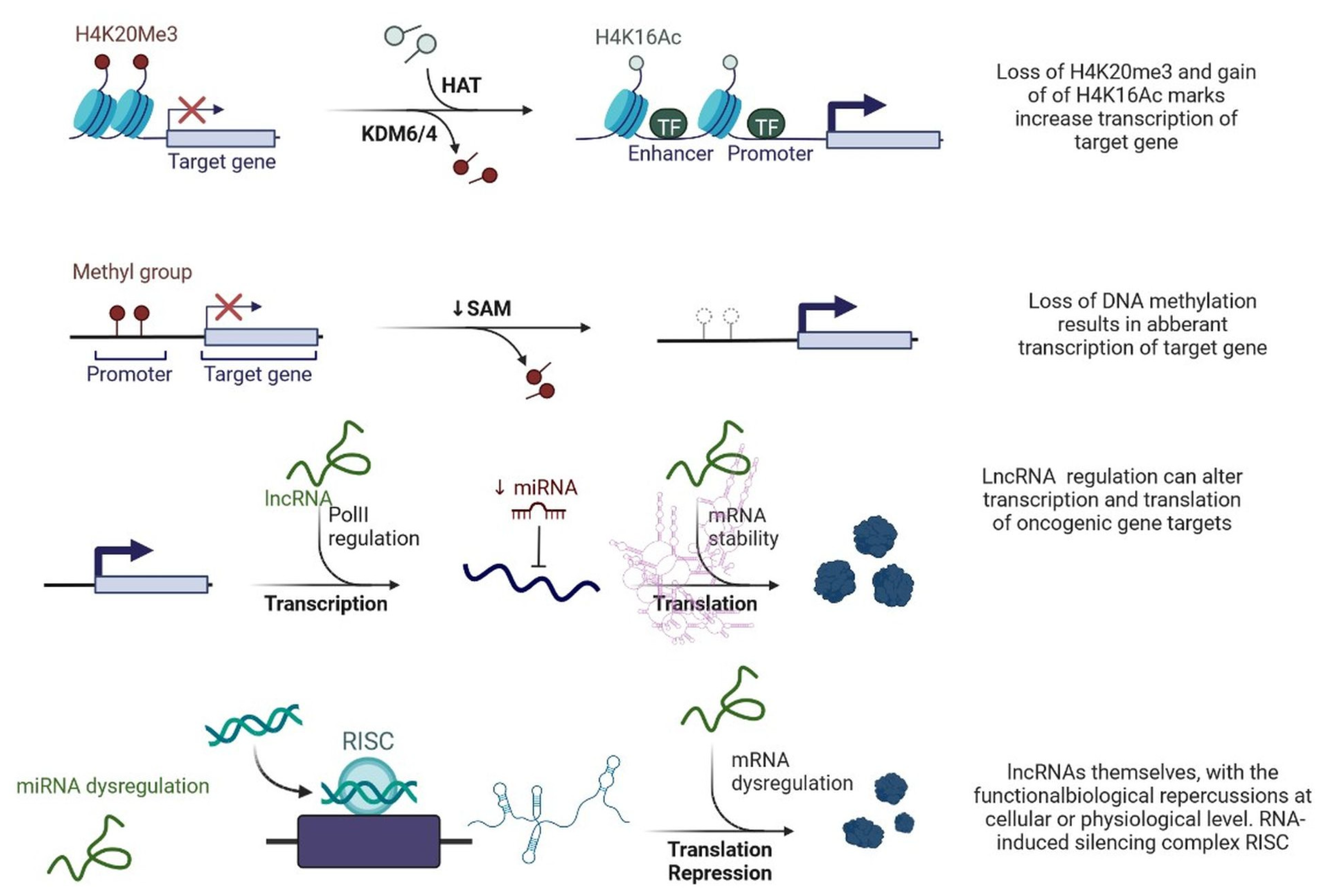
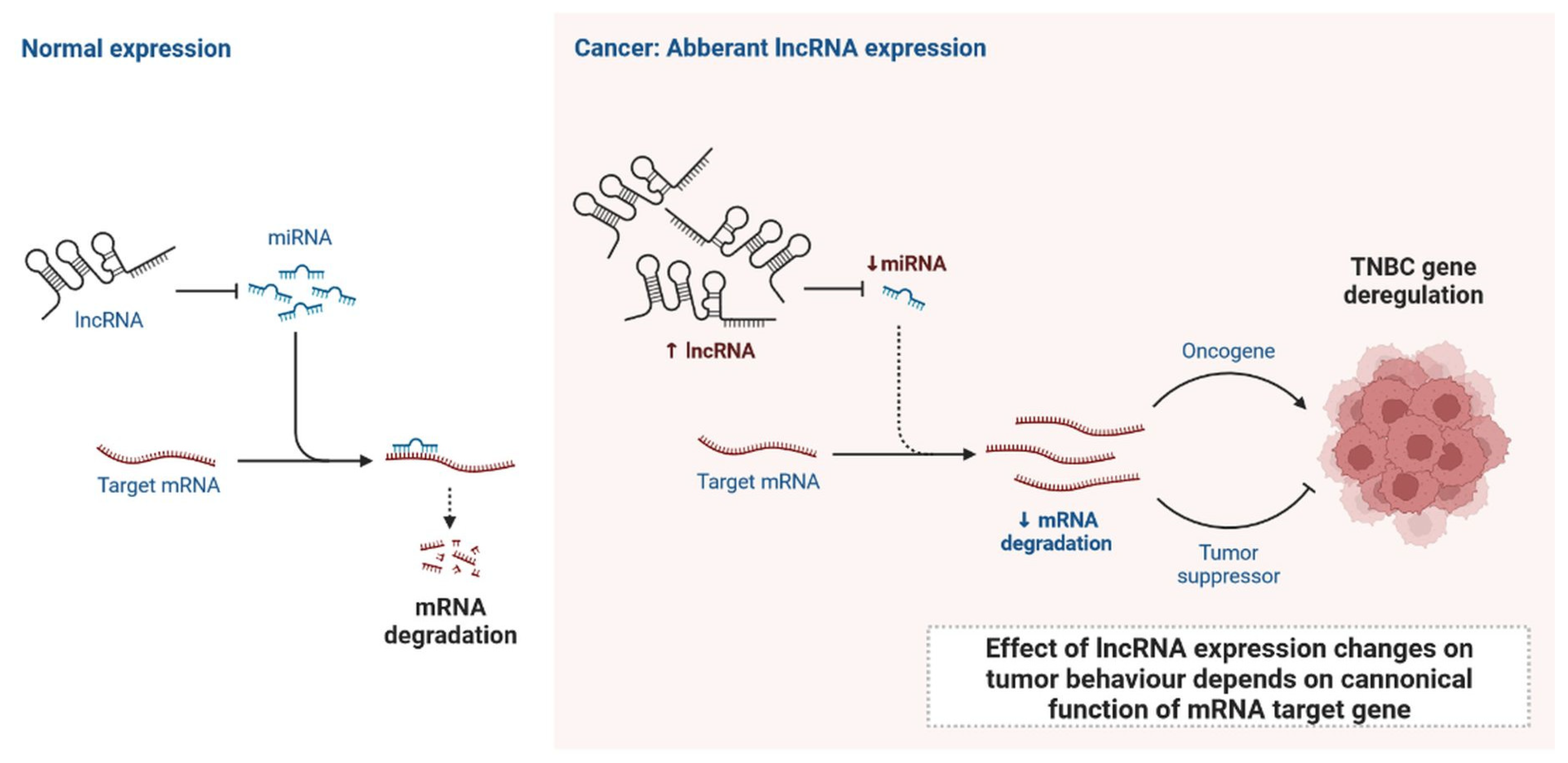
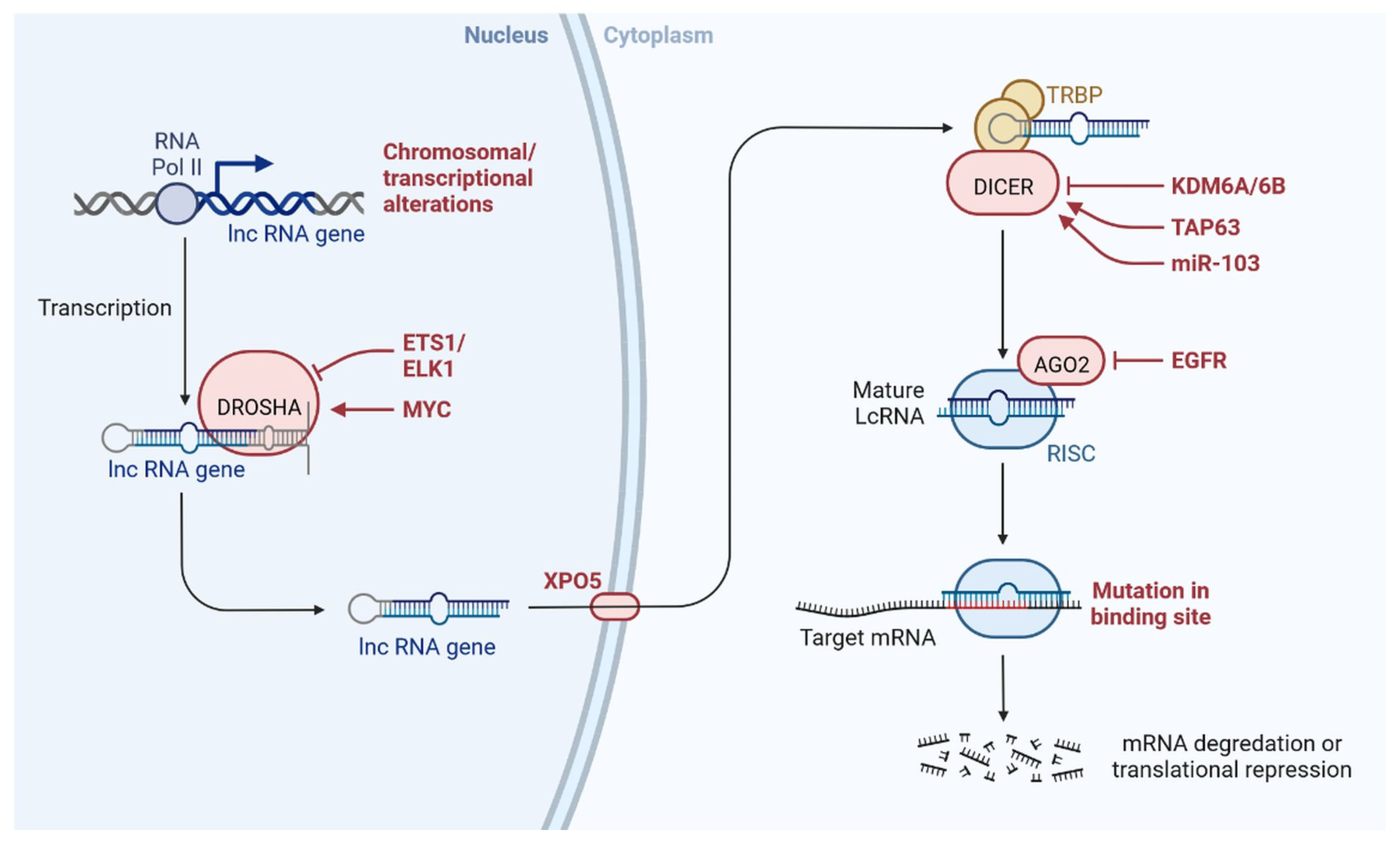
2. LncRNAs
3. Clinical Updates on lncRNAs in TNBC
3.1. Importance of lncRNAs in Tumour Invasiveness and Metastasis
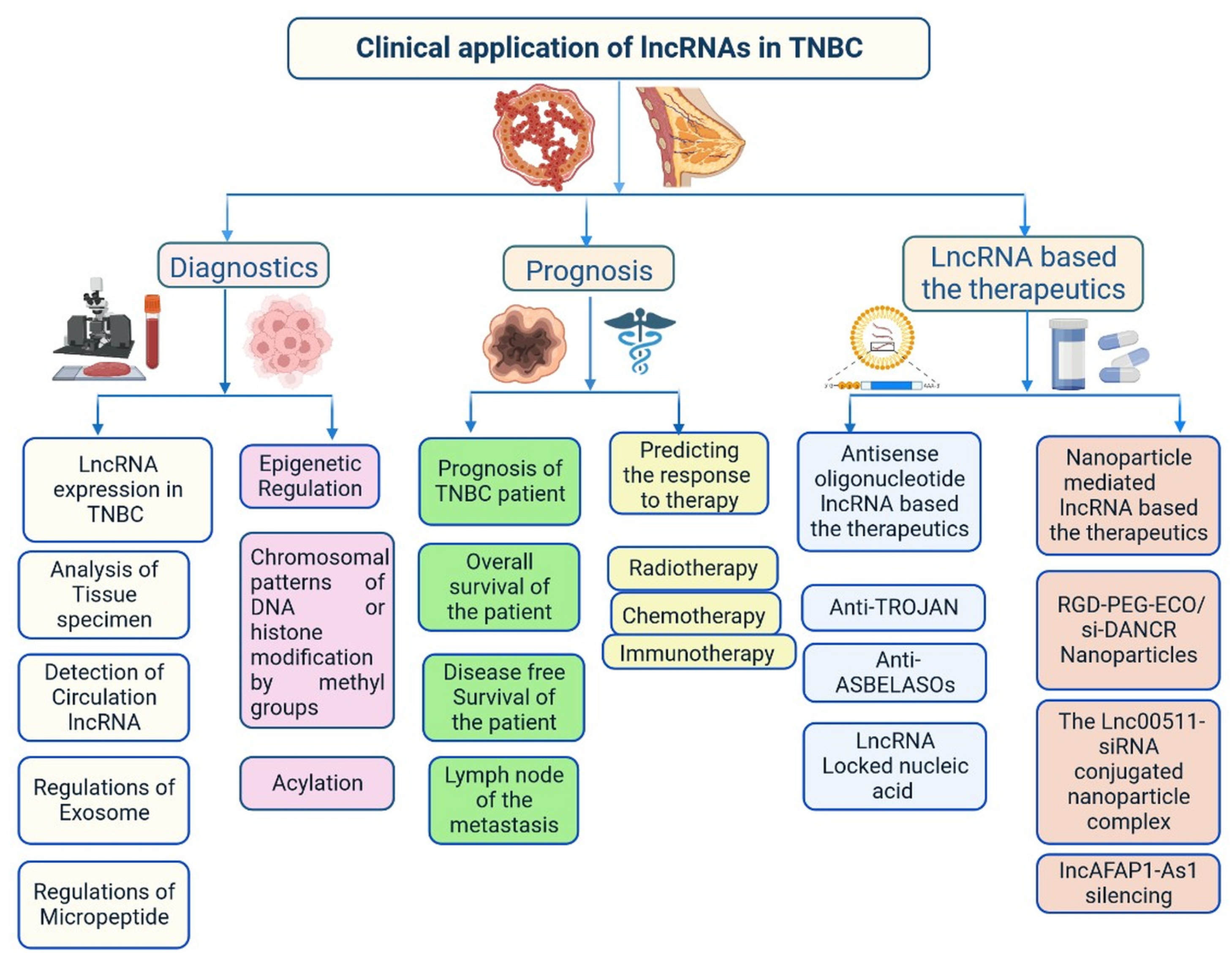
3.2. Importance of lncRNAs in Clinical Diagnosis
3.3. Importance of lncRNAs in Treatment
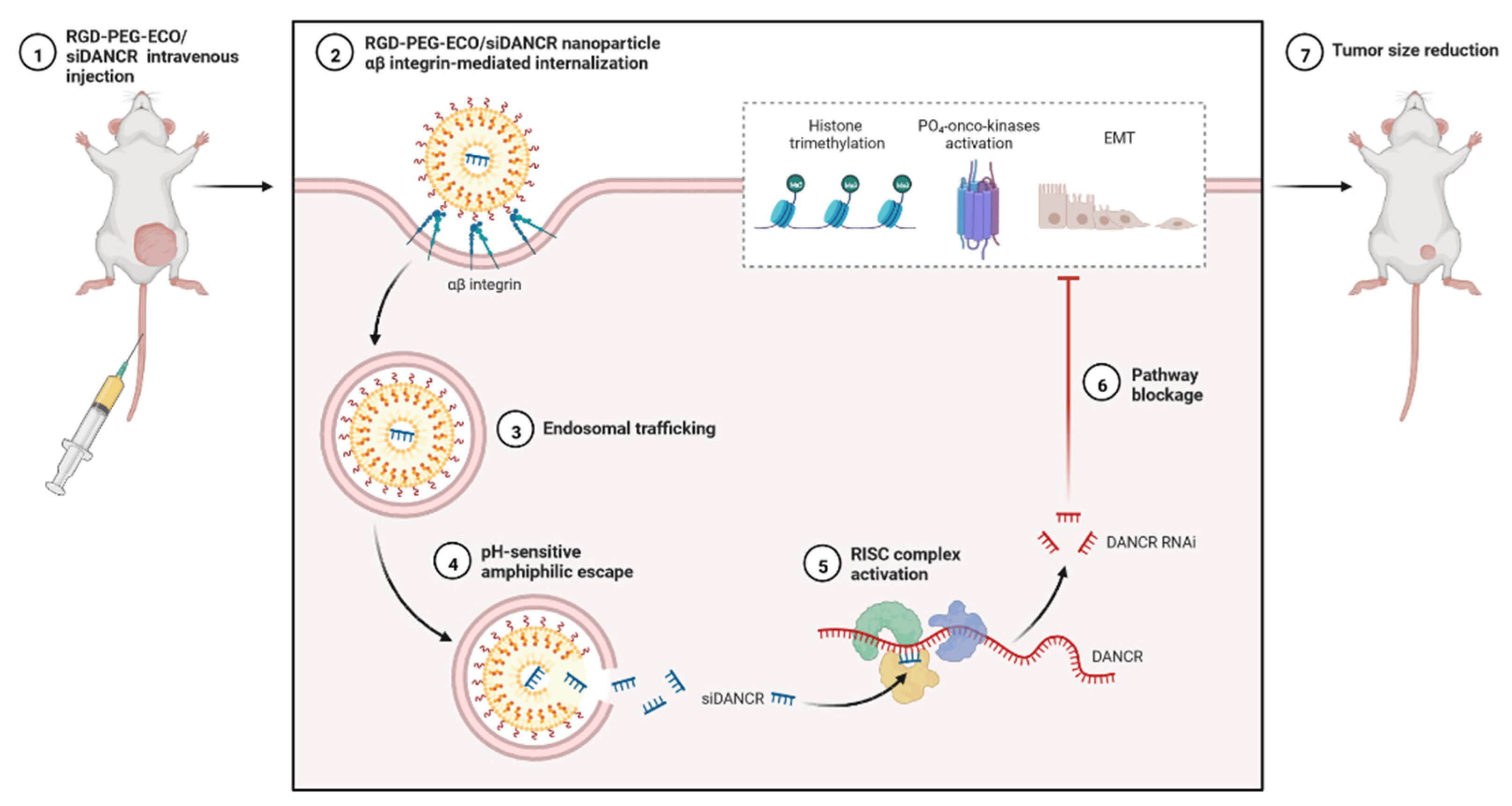
4. Nanoparticle-Based Targeted Therapy with ncRNAs for TNBC
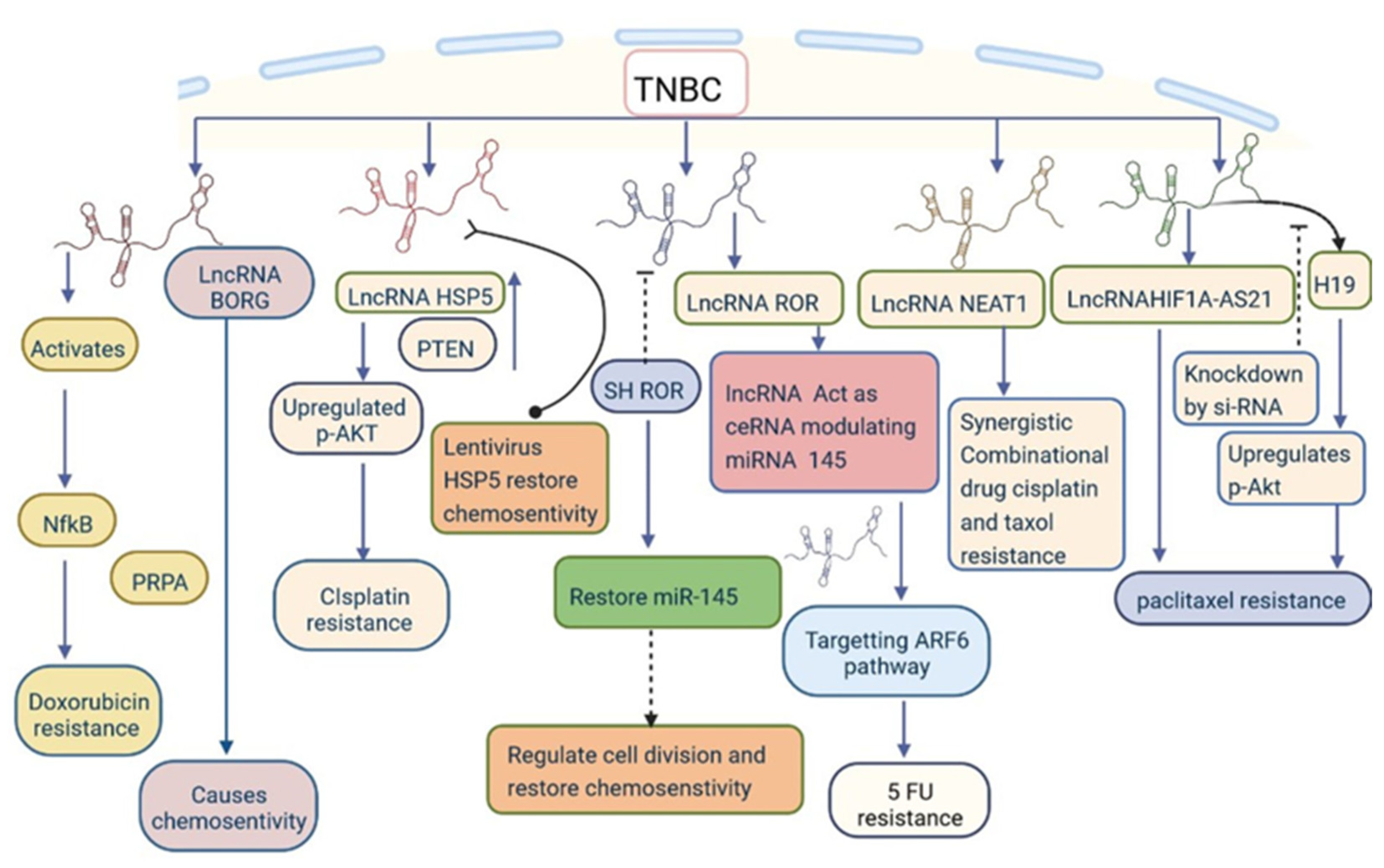
5. LncRNAs Involved in The Regulation of Drug Resistance
6. Future Perspectives of lncRNAs as Potential Diagnosis and Pharmacological Tools/Targets
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onitilo, A.A.; Engel, J.M.; Greenlee, R.T.; Mukesh, B.N. Breasreastct Cancer Subtypes Based on ER/PR and Her2 Expression: Comparison of Clinicopathologic Features and Survival. Clin. Med. Res. 2009, 7, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Marra, A.; Trapani, D.; Viale, G.; Criscitiello, C.; Curigliano, G. Practical Classification of Triple-Negative Breast Cancer: Intratumoral Heterogeneity, Mechanisms of Drug Resistance, and Novel Therapies. NPJ Breast Cancer 2020, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Aine, M.; Boyaci, C.; Hartman, J.; Häkkinen, J.; Mitra, S.; Campos, A.B.; Nimeus, E.; Ehinger, A.; Vallon-Christersson, J.; Borg, Å.; et al. Molecular Analyses of Triple-Negative Breast Cancer in the Young and Elderly. Breast Cancer Res. 2021, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Zagami, P.; Carey, L.A. Triple negative breast cancer: Pitfalls and progress. NPJ Breast Cancer 2022, 8, 95. [Google Scholar] [CrossRef]
- Bissanum, R.; Chaichulee, S.; Kamolphiwong, R.; Navakanitworakul, R.; Kanokwiroon, K. Molecular Classification Models for Triple Negative Breast Cancer Subtype Using Machine Learning. J. Pers. Med. 2021, 11, 881. [Google Scholar] [CrossRef] [PubMed]
- Volovat, S.R.; Volovat, C.; Hordila, I.; Hordila, D.-A.; Mirestean, C.C.; Miron, O.T.; Lungulescu, C.; Scripcariu, D.V.; Stolniceanu, C.R.; Konsoulova-Kirova, A.A.; et al. MiRNA and LncRNA as Potential Biomarkers in Triple-Negative Breast Cancer: A Review. Front. Oncol. 2020, 10, 526850. [Google Scholar] [CrossRef]
- Lee, J.; Park, H.Y.; Kim, W.W.; Lee, S.J.; Jeong, J.H.; Kang, S.H.; Jung, J.H.; Chae, Y.S. Biological function of long noncoding RNA snaR in HER2-positive breast cancer cells. Tumour Biol. 2017, 39, 1010428317707374. [Google Scholar] [CrossRef] [Green Version]
- Melone, V.; Salvati, A.; Brusco, N.; Alexandrova, E.; D’Agostino, Y.; Palumbo, D.; Palo, L.; Terenzi, I.; Nassa, G.; Rizzo, F.; et al. Functional Relationships between Long Non-Coding RNAs and Estrogen Receptor Alpha: A New Frontier in Hormone-Responsive Breast Cancer Management. Int. J. Mol. Sci. 2023, 24, 1145. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, Q.; Xing, W.; Wang, W. Role of MicroRNA/LncRNA Intertwined with the Wnt/β-Catenin Axis in Regulating the Pathogenesis of Triple-Negative Breast Cancer. Front. Pharmacol. 2022, 13, 814971. [Google Scholar] [CrossRef]
- Mou, E.; Wang, H. LncRNA LUCAT1 Facilitates Tumorigenesis and Metastasis of Triple-Negative Breast Cancer through Modulating MiR-5702. Biosci. Rep. 2019, 39, BSR20190489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, L.; Xiao, X.; Zhao, Y.; Yin, L.; Fu, M.; Zhang, X.; Jiang, P. The Functional Roles of Long Noncoding RNA DANCR in Human Cancers. J. Cancer 2020, 11, 6970–6981. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Liu, L.; Wang, Z. Long Non-coding RNA Heart and Neural Crest Derivatives Expressed 2-antisense RNA 1 Overexpression Inhibits the Proliferation of Cancer Cells by Reducing RUNX2 Expression in Triple-negative Breast Cancer. Oncol. Lett. 2019, 18, 6775–6780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, N.; Liu, Y.; Su, P.; Liang, Y.; Li, Y.; Wang, X.; Chen, T.; Song, X.; Sang, Y.; et al. Epigenetic Regulation of NAMPT by NAMPT-AS Drives Metastatic Progression in Triple-Negative Breast Cancer. Cancer Res. 2019, 79, 3347–3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, S.; Bai, R.; Lu, D.; Chen, C.; Ma, T.; Zou, Z.; Zhang, Z. Long Non-Coding RNA MAFG-AS1 Promotes Proliferation and Metastasis of Breast Cancer by Modulating STC2 Pathway. Cell Death Discov. 2022, 8, 249. [Google Scholar] [CrossRef]
- Du, T.; Shi, Y.; Xu, S.; Wan, X.; Sun, H.; Liu, B. Long Non-Coding RNAs in Drug Resistance of Breast Cancer. OncoTargets Ther. 2020, 13, 7075–7087. [Google Scholar] [CrossRef]
- Mei, J.; Hao, L.; Wang, H.; Xu, R.; Liu, Y.; Zhu, Y.; Liu, C. Systematic Characterization of Non-coding RNAs in Triple-negative Breast Cancer. Cell Prolif. 2020, 53, e12801. [Google Scholar] [CrossRef] [Green Version]
- Richard, J.L.C.; Eichhorn, P.J.A. Deciphering the Roles of LncRNAs in Breast Development and Disease. Oncotarget 2018, 9, 20179–20212. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Ke, H.; Zhang, H.; Ma, Y.; Ao, L.; Zou, L.; Yang, Q.; Zhu, H.; Nie, J.; Wu, C.; et al. LncRNA MIR100HG Promotes Cell Proliferation in Triple-Negative Breast Cancer through Triplex Formation with P27 Loci. Cell Death Discov. 2018, 9, 805. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wang, Z.; Yu, S.; Liu, D.; Sun, L. LncmiRHG-MIR100HG: A New Budding Star in Cancer. Front. Oncol. 2022, 12, 997532. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, N.; Song, P.; Fu, Y.; Ren, Y.; Li, Z.; Wang, J. LncRNA GATA3-AS1 Facilitates Tumour Progression and Immune Escape in Triple-negative Breast Cancer through Destabilization of GATA3 but Stabilization of PD-L1. Cell Prolif. 2020, 53, e12855. [Google Scholar] [CrossRef]
- Qi, F.; Qin, W.; Zang, Y. Molecular Mechanism of Triple-negative Breast Cancer-associated BRCA1 and the Identification of Signaling Pathways. Oncol. Lett. 2019, 17, 2905–2914. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tan, Y.; Wang, H.; Xu, M.; Xu, L. Long Non-Coding RNA Plasmacytoma Variant Translocation 1 (PVT1) Enhances Proliferation, Migration, and Epithelial-Mesenchymal Transition (EMT) of Pituitary Adenoma Cells by Activating β-Catenin, c-Myc, and Cyclin D1 Expression. Med. Sci. Monit. 2019, 25, 7652–7659. [Google Scholar] [CrossRef]
- Tang, J.; Li, Y.; Sang, Y.; Yu, B.; Lv, D.; Zhang, W.; Feng, H. LncRNA PVT1 Regulates Triple-Negative Breast Cancer through KLF5/Beta-Catenin Signaling. Oncogene 2018, 37, 4723–4734. [Google Scholar] [CrossRef]
- Zhang, W.; Guan, X.; Tang, J. The Long Non-coding RNA Landscape in Triple-negative Breast Cancer. Cell Prolif. 2021, 54, e12966. [Google Scholar] [CrossRef]
- López-Urrutia, E.; Bustamante Montes, L.P.; Ladrón de Guevara Cervantes, D.; Pérez-Plasencia, C.; Campos-Parra, A.D. Crosstalk between Long Non-Coding RNAs, Micro-RNAs and MRNAs: Deciphering Molecular Mechanisms of Master Regulators in Cancer. Front. Oncol. 2019, 9, 669. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Song, G.; Li, M.; Hao, X.; Huang, Y.; Lan, J.; Yang, S.; Zhang, Z.; Zhang, G.; Mu, J. Construction and Comprehensive Analysis of a CeRNA Network to Reveal Potential Novel Biomarkers for Triple-Negative Breast Cancer. Cancer Manag. Res. 2020, 12, 7061–7075. [Google Scholar] [CrossRef] [PubMed]
- Le, K.; Guo, H.; Zhang, Q.; Huang, X.; Xu, M.; Huang, Z.; Yi, P. Gene and LncRNA Co-Expression Network Analysis Reveals Novel CeRNA Network for Triple-Negative Breast Cancer. Sci. Rep. 2019, 9, 15122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Mi, M.; Li, X.; Zheng, X.; Wu, G.; Zhang, L. LncRNA OSTN-AS1 May Represent a Novel Immune-Related Prognostic Marker for Triple-Negative Breast Cancer Based on Integrated Analysis of a CeRNA Network. Front. Genet. 2019, 10, 850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, G.; Han, J. HIF1A-AS2 Predicts Poor Prognosis and Regulates Cell Migration and Invasion in Triple-negative Breast Cancer. J. Cell. Biochem. 2019, 120, 10513–10518. [Google Scholar] [CrossRef]
- Zhao, W.; Geng, D.; Li, S.; Chen, Z.; Sun, M. LncRNA HOTAIR Influences Cell Growth, Migration, Invasion, and Apoptosis via the MiR-20a-5p/HMGA2 Axis in Breast Cancer. Cancer Med. 2018, 7, 842–855. [Google Scholar] [CrossRef]
- Yu, F.; Wang, L.; Zhang, B. Long Non-coding RNA DRHC Inhibits the Proliferation of Cancer Cells in Triple Negative Breast Cancer by Downregulating Long Non-coding RNA HOTAIR. Oncol. Lett. 2019, 18, 3817–3822. [Google Scholar] [CrossRef] [Green Version]
- Tu, Z.; Schmöllerl, J.; Cuiffo, B.G.; Karnoub, A.E. Microenvironmental Regulation of Long Noncoding RNA LINC01133 Promotes Cancer Stem Cell-Like Phenotypic Traits in Triple-Negative Breast Cancers. Stem Cells 2019, 37, 1281–1292. [Google Scholar] [CrossRef]
- Zhao, M.; Zhang, M.; Tao, Z.; Cao, J.; Wang, L.; Hu, X. MiR-331-3p Suppresses Cell Proliferation in TNBC Cells by Downregulating NRP2. Technol. Cancer Res. Treat. 2020, 19, 153303382090582. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Zhou, B.; Lv, Y.; Qian, Q. LncRNA HEIH Regulates Cell Proliferation and Apoptosis through MiR-4458/SOCS1 Axis in Triple-Negative Breast Cancer. Human Cell 2019, 32, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Shen, Y.; Zhang, W.; Jin, J.; Huang, D.; Fang, H.; Ji, W.; Shi, Y.; Tang, L.; Chen, W.; et al. An Androgen Receptor Negatively Induced Long Non-Coding RNA ARNILA Binding to MiR-204 Promotes the Invasion and Metastasis of Triple-Negative Breast Cancer. Cell Death Differ. 2018, 25, 2209–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tariq, A.; Hao, Q.; Sun, Q.; Singh, D.K.; Jadaliha, M.; Zhang, Y.; Chetlangia, N.; Ma, J.; Holton, S.E.; Bhargava, R.; et al. LncRNA-Mediated Regulation of SOX9 Expression in Basal Subtype Breast Cancer Cells. RNA 2020, 26, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xi, C.; Yang, X.; Lu, X.; Yu, K.; Zhang, Y.; Gao, R. LncRNA WT1-AS Inhibits Triple-Negative Breast Cancer Cell Migration and Invasion by Downregulating Transforming Growth Factor Β1. Cancer Biother. Radiopharm. 2019, 34, 671–675. [Google Scholar] [CrossRef]
- Filippova, E.A.; Fridman, M.V.; Burdennyy, A.M.; Loginov, V.I.; Pronina, I.V.; Lukina, S.S.; Dmitriev, A.A.; Braga, E.A. Long Noncoding RNA GAS5 in Breast Cancer: Epigenetic Mechanisms and Biological Functions. Int. J. Mol. Sci. 2021, 22, 6810. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Li, X.; Fan, Q.; Liu, G.; Yin, J. CCAT1 Promotes Triple-Negative Breast Cancer Progression by Suppressing MiR-218/ZFX Signaling. Aging 2019, 11, 4858–4875. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, S.; Zhu, X.; Zhang, L.; Deng, J.; Li, F.; Guo, B.; Zhang, S.; Wu, R.; Zhang, Z.; et al. LncRNA-Encoded Polypeptide ASRPS Inhibits Triple-Negative Breast Cancer Angiogenesis. J. Exp. Med. 2020, 217, e20190950. [Google Scholar] [CrossRef]
- Tang, M.; Liu, Y.; Zhang, Q.; Zhang, P.; Wu, J.; Wang, J.; Ruan, Y.; Huang, Y. Antitumor Efficacy of the Runx2-dendritic Cell Vaccine in Triple-negative Breast Cancer In Vitro. Oncol. Lett. 2018, 16, 2813–2822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Meng, X.; Yu, Y.; Pan, L.; Zheng, Q.; Lin, W. LncRNA POU3F3 Promotes Proliferation and Inhibits Apoptosis of Cancer Cells in Triple-Negative Breast Cancer by Inactivating Caspase 9. Biosci. Biotechnol. Biochem. 2019, 83, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, X.; Liang, Y.; Ye, G.; Che, Y.; Wu, X.; Zhu, X.; Fan, H.; Fan, X.; Xu, J. MiR-155 Increases Stemness and Decitabine Resistance in Triple-negative Breast Cancer Cells by Inhibiting TSPAN5. Mol. Carcinog. 2020, 59, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Deng, L.; Wang, Y.-D. Roles and Mechanisms of Long Non-Coding RNAs in Breast Cancer. Int. J. Mol. Sci. 2023, 24, 89. [Google Scholar] [CrossRef]
- Tian, Y.; Xia, S.; Ma, M.; Zuo, Y. LINC00096 Promotes the Proliferation and Invasion by Sponging MiR-383-5p and Regulating RBM3 Expression in Triple-Negative Breast Cancer. OncoTargets Ther. 2019, 12, 10569–10578. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Zhao, W.; Zhang, H.; Chu, Z.; Liu, H.; Fang, X.; Tang, D. Long Non-Coding RNA ANRIL Promotes Chemoresistance in Triple-Negative Breast Cancer via Enhancing Aerobic Glycolysis. Life Sci. 2022, 306, 120810. [Google Scholar] [CrossRef]
- Tamang, S.; Acharya, V.; Roy, D.; Sharma, R.; Aryaa, A.; Sharma, U.; Khandelwal, A.; Prakash, H.; Vasquez, K.M.; Jain, A. SNHG12: An LncRNA as a Potential Therapeutic Target and Biomarker for Human Cancer. Front. Oncol. 2019, 9, 901. [Google Scholar] [CrossRef]
- Xia, L.; Wang, H. LncRNA LUCAT1/ELAVL1/LIN28B/SOX2 Positive Feedback Loop Promotes Cell Stemness in Triple-Negative Breast Cancer. Breast J. 2022, 2022, 1–12. [Google Scholar] [CrossRef]
- Shi, R.; Wu, P.; Liu, M.; Chen, B.; Cong, L. Knockdown of LncRNA PCAT6 Enhances Radiosensitivity in Triple-Negative Breast Cancer Cells by Regulating MiR-185-5p/TPD52 Axis. OncoTargets Ther. 2020, 13, 3025–3037. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.-X.; Chen, S.; Huang, L.; Shao, Z.-M. Predictive and Prognostic Value of Matrix Metalloproteinase (MMP)-9 in Neoadjuvant Chemotherapy for Triple-Negative Breast Cancer Patients. BMC Cancer 2018, 18, 909. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Katsuta, E.; Oshi, M.; Sporn, J.C.; Yan, L.; Le, L.; Matsuhashi, N.; Futamura, M.; Akao, Y.; Yoshida, K.; et al. High Expression of miR-34a Associated with Less Aggressive Cancer Biology but Not with Survival in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 3045. [Google Scholar] [CrossRef]
- Wang, L.; Luan, T.; Zhou, S.; Lin, J.; Yang, Y.; Liu, W.; Tong, X.; Jiang, W. LncRNA HCP5 Promotes Triple Negative Breast Cancer Progression as a CeRNA to Regulate BIRC3 by Sponging MiR-219a-5p. Cancer Med. 2019, 8, 4389–4403. [Google Scholar] [CrossRef]
- Han, C.; Fu, Y.; Zeng, N.; Yin, J.; Li, Q. LncRNA FAM83H-AS1 Promotes Triple-Negative Breast Cancer Progression by Regulating the MiR-136-5p/Metadherin Axis. Aging 2020, 12, 3594–3616. [Google Scholar] [CrossRef]
- Niu, L.; Fan, Q.; Yan, M.; Wang, L. LncRNA NRON Down-Regulates LncRNA SnaR and Inhibits Cancer Cell Proliferation in TNBC. Biosci. Rep. 2019, 39, BSR20190468. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Qu, H.; Gong, W.; Xiang, J.; Yang, M.; Zhang, W. LncRNA AWPPH and MiRNA-21 Regulates Cancer Cell Proliferation and Chemosensitivity in Triple-negative Breast Cancer by Interacting with Each Other. J. Cell. Biochem. 2019, 120, 14860–14866. [Google Scholar] [CrossRef]
- Youness, R.A.; Hafez, H.M.; Khallaf, E.; Assal, R.A.; Abdel Motaal, A.; Gad, M.Z. The Long Noncoding RNA SONE Represses Triple-negative Breast Cancer Aggressiveness through Inducing the Expression of MiR-34a, MiR-15a, MiR-16, and Let-7a. J. Cell. Physiol. 2019, 234, 20286–20297. [Google Scholar] [CrossRef]
- Jin, S.-J.; Jin, M.-Z.; Xia, B.-R.; Jin, W.-L. Long Non-Coding RNA DANCR as an Emerging Therapeutic Target in Human Cancers. Front. Oncol. 2019, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Li, C.; Xing, Z.; Hu, Q.; Liang, K.; Han, L.; Wang, C.; Hawke, D.H.; Wang, S.; Zhang, Y.; et al. The LINK-A LncRNA Activates Normoxic HIF1α Signalling in Triple-Negative Breast Cancer. Nat. Cell. Biol. 2016, 18, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Dong, G.; Shi, H.; Zhang, J.; Ning, Z.; Bao, X.; Liu, C.; Hu, J.; Liu, M.; Xiong, B. LncRNA MIR503HG Inhibits Cell Migration and Invasion via MiR-103/OLFM4 Axis in Triple Negative Breast Cancer. J. Cell. Mol. Med. 2019, 23, 4738–4745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kansara, S.; Pandey, V.; Lobie, P.E.; Sethi, G.; Garg, M.; Pandey, A.K. Mechanistic Involvement of Long Non-Coding RNAs in Oncotherapeutics Resistance in Triple-Negative Breast Cancer. Cells 2020, 9, 1511. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hou, M.; Zhan, Y.; Sheng, X. LncRNA PTCSC3 Inhibits Triple-negative Breast Cancer Cell Proliferation by Downregulating LncRNA H19. J. Cell. Biochem. 2019, 120, 15083–15088. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Xiong, Z.; Li, R.; Liang, H.; Jia, C.; Deng, M. Long Non-Coding RNA NRON Is Downregulated in HCC and Suppresses Tumour Cell Proliferation and Metastasis. Biomed. Pharmacother. 2018, 104, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Goh, C.Y.; Wyse, C.; Ho, M.; O’Beirne, E.; Howard, J.; Lindsay, S.; Kelly, P.; Higgins, M.; McCann, A. Exosomes in Triple Negative Breast Cancer: Garbage Disposals or Trojan Horses? Cancer Lett. 2020, 473, 90–97. [Google Scholar] [CrossRef]
- Tracy, K.M.; Tye, C.E.; Ghule, P.N.; Malaby, H.L.H.; Stumpff, J.; Stein, J.L.; Stein, G.S.; Lian, J.B. Mitotically-Associated LncRNA (MANCR) Affects Genomic Stability and Cell Division in Aggressive Breast Cancer. Mol. Cancer Res. 2018, 16, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, D.; Wu, X.; Zeng, Y.; Li, L.; Hou, Y.; Li, W.; Liu, Z. Long Non-coding RNA (LncRNA) RMST in Triple-negative Breast Cancer (TNBC): Expression Analysis and Biological Roles Research. J. Cell. Physiol. 2018, 233, 6603–6612. [Google Scholar] [CrossRef]
- Ferraro, D.A.; Patella, F.; Zanivan, S.; Donato, C.; Aceto, N.; Giannotta, M.; Dejana, E.; Diepenbruck, M.; Christofori, G.; Buess, M. Endothelial Cell-Derived Nidogen-1 Inhibits Migration of SK-BR-3 Breast Cancer Cells. BMC Cancer 2019, 19, 312. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Kim, Y.-J.; Ji, Z.; Kang, J.M.; Wirianto, M.; Paudel, K.R.; Smith, J.A.; Ono, K.; Kim, J.-A.; Eckel-Mahan, K.; et al. ROR Activation by Nobiletin Enhances Antitumor Efficacy via Suppression of IκB/NF-ΚB Signaling in Triple-Negative Breast Cancer. Cell Death Dis. 2022, 13, 374. [Google Scholar] [CrossRef]
- Merikhian, P.; Eisavand, M.R.; Farahmand, L. Triple-Negative Breast Cancer: Understanding Wnt Signaling in Drug Resistance. Cancer Cell Int. 2021, 21, 419. [Google Scholar] [CrossRef]
- Wang, P.-S.; Chou, C.-H.; Lin, C.-H.; Yao, Y.-C.; Cheng, H.-C.; Li, H.-Y.; Chuang, Y.-C.; Yang, C.-N.; Ger, L.-P.; Chen, Y.-C.; et al. A Novel Long Non-Coding RNA Linc-ZNF469-3 Promotes Lung Metastasis through MiR-574-5p-ZEB1 Axis in Triple Negative Breast Cancer. Oncogene 2018, 37, 4662–4678. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Z.; Zhang, L.; Sun, S. LncRNA PDCD4-AS1 Alleviates Triple Negative Breast Cancer by Increasing Expression of IQGAP2 via MiR-10b-5p. Transl. Oncol. 2021, 14, 100958. [Google Scholar] [CrossRef] [PubMed]
- Hua, K.; Deng, X.; Hu, J.; Ji, C.; Yu, Y.; Li, J.; Wang, X.; Fang, L. Long Noncoding RNA HOST2, Working as a Competitive Endogenous RNA, Promotes STAT3-Mediated Cell Proliferation and Migration via Decoying of Let-7b in Triple-Negative Breast Cancer. J. Exp. Clin. Cancer Res. 2020, 39, 58. [Google Scholar] [CrossRef] [PubMed]
- Jahangiri, L.; Ishola, T. Dormancy in Breast Cancer, the Role of Autophagy, lncRNAs, miRNAs and Exosomes. Int. J. Mol. Sci. 2022, 23, 5271. [Google Scholar] [CrossRef]
- Tao, W.; Sun, W.; Zhu, H.; Zhang, J. Knockdown of Long Non-Coding RNA TP73-AS1 Suppresses Triple Negative Breast Cancer Cell Vasculogenic Mimicry by Targeting MiR-490-3p/TWIST1 Axis. Biochem. Biophys. Res. Commun. 2018, 504, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Sun, L.; Wan, F. Molecular Mechanisms of TUG1 in the Proliferation, Apoptosis, Migration and Invasion of Cancer Cells (Review). Oncol. Lett. 2019, 18, 4393–4402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.-Y.; Zhou, Z.-Y.; Zhang, K.-J.; Pang, J.; Wang, S.-M. Long Non-Coding RNA MIR100HG Promotes the Migration, Invasion and Proliferation of Triple-Negative Breast Cancer Cells by Targeting the MiR-5590-3p/OTX1 Axis. Cancer Cell Int. 2020, 20, 508. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Tang, H.; Ling, L.; Li, N.; Jia, X.; Zhang, Z.; Wang, X.; Shi, L.; Yin, J.; Qiu, N.; et al. LINC01638 LncRNA Activates MTDH-Twist1 Signaling by Preventing SPOP-Mediated c-Myc Degradation in Triple-Negative Breast Cancer. Oncogene 2018, 37, 6166–6179. [Google Scholar] [CrossRef]
- Al-Mahmood, S.; Sapiezynski, J.; Garbuzenko, O.B.; Minko, T. Metastatic and Triple-Negative Breast Cancer: Challenges and Treatment Options. Drug Deliv. Transl. Res. 2018, 8, 1483–1507. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhou, J.; Wang, Z.; Wang, P.; Gao, X.; Wang, Y. Long Noncoding RNA GAS5 Suppresses Triple Negative Breast Cancer Progression through Inhibition of Proliferation and Invasion by Competitively Binding MiR-196a-5p. Biomed. Pharmacother. 2018, 104, 451–457. [Google Scholar] [CrossRef]
- Jin, X.; Xu, X.-E.; Jiang, Y.-Z.; Liu, Y.-R.; Sun, W.; Guo, Y.-J.; Ren, Y.-X.; Zuo, W.-J.; Hu, X.; Huang, S.-L.; et al. The Endogenous Retrovirus-Derived Long Noncoding RNA TROJAN Promotes Triple-Negative Breast Cancer Progression via ZMYND8 Degradation. Sci. Adv. 2019, 5, eaat9820. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Liu, P.; Tang, H.; Xie, X.; Kong, Y.; Song, C.; Qiu, X.; Xiao, X. AFAP1-AS1 Promotes Epithelial-Mesenchymal Transition and Tumorigenesis Through Wnt/β-Catenin Signaling Pathway in Triple-Negative Breast Cancer. Front. Pharmacol. 2018, 9, 1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imani, S.; Wei, C.; Cheng, J.; Khan, A.; Fu, S.; Yang, L.; Tania, M.; Zhang, X.; Xiao, X.; Zhang, X.; et al. MicroRNA-34a Targets Epithelial to Mesenchymal Transition-Inducing Transcription Factors (EMT-TFs) and Inhibits Breast Cancer Cell Migration and Invasion. Oncotarget 2017, 8, 21362–21379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Geng, C.; Dong, Q. LncRNA PAPAS May Promote Triple-Negative Breast Cancer by Downregulating MiR-34a. J. Int. Med. Res. 2019, 47, 3709–3718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, H.; Sun, R.; Li, P.; Yang, Z.; Liu, Y.; Wang, Z.; Yang, Y.; Yin, C. Long Non-coding RNA ZEB2-AS1 Promotes the Proliferation, Metastasis and Epithelial Mesenchymal Transition in Triple-negative Breast Cancer by Epigenetically Activating ZEB2. J. Cell. Mol. Med. 2019, 23, 3271–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Y.; Li, Y.; Zhou, Z.; Ma, M.; Fu, K. Long Non-Coding RNA MALAT1 Promotes Proliferation and Invasion via Targeting MiR-129-5p in Triple-Negative Breast Cancer. Biomed. Pharmacother. 2017, 95, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Mofed, D.; Omran, J.I.; Sabet, S.; Baiomy, A.A.; Emara, M.; Salem, T.Z. The Regulatory Role of Long Non-Coding RNAs as a Novel Controller of Immune Response against Cancer Cells. Mol. Biol. Rep. 2022, 49, 11775–11793. [Google Scholar] [CrossRef]
- Zuo, K.; Yuan, X.; Liang, X.; Sun, X.; Liu, S.; Connell, P.P.; Li, X.; Yang, W. QRT-PCR-Based DNA Homologous Recombination-Associated 4-Gene Score Predicts Pathologic Complete Response to Platinum-Based Neoadjuvant Chemotherapy in Triple-Negative Breast Cancer. Breast Cancer Res. Treat. 2022, 191, 335–344. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, S.; Chen, D.; Yuwen, D.; Zhang, J.; Wei, X.; Han, X.; Guan, X. SOX2-OT Induced by PAI-1 Promotes Triple-Negative Breast Cancer Cells Metastasis by Sponging MiR-942-5p and Activating PI3K/Akt Signaling. Cell. Mol. Life Sci. 2022, 79, 59. [Google Scholar] [CrossRef]
- Zhou, Y.; Meng, X.; Chen, S.; Li, W.; Li, D.; Singer, R.; Gu, W. IMP1 Regulates UCA1-Mediated Cell Invasion through Facilitating UCA1 Decay and Decreasing the Sponge Effect of UCA1 for MiR-122-5p. Breast Cancer Res. 2018, 20, 32. [Google Scholar] [CrossRef]
- Manoochehri, M.; Jones, M.; Tomczyk, K.; Fletcher, O.; Schoemaker, M.J.; Swerdlow, A.J.; Borhani, N.; Hamann, U. DNA Methylation of the Long Intergenic Noncoding RNA 299 Gene in Triple-Negative Breast Cancer: Results from a Prospective Study. Sci. Rep. 2020, 10, 11762. [Google Scholar] [CrossRef]
- Zheng, S.; Li, M.; Miao, K.; Xu, H. LncRNA GAS5-promoted Apoptosis in Triple-negative Breast Cancer by Targeting MiR-378a-5p/SUFU Signaling. J. Cell. Biochem. 2020, 121, 2225–2235. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Zhu, Q.; Lu, H.; Chang, A.; Gao, C.; Zhou, Q.; Luo, K. HEXIM1 Controls P-TEFb Processing and Regulates Drug Sensitivity in Triple-Negative Breast Cancer. Mol. Biol. Cell 2020, 31, 1867–1878. [Google Scholar] [CrossRef] [PubMed]
- Aranza-Martínez, A.; Sánchez-Pérez, J.; Brito-Elias, L.; López-Camarillo, C.; Cantú de León, D.; Pérez-Plasencia, C.; López-Urrutia, E. Non-Coding RNAs Associated With Radioresistance in Triple-Negative Breast Cancer. Front. Oncol. 2021, 11, 752270. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Mangala, L.S.; Rodriguez-Aguayo, C.; Kong, X.; Lopez-Berestein, G.; Sood, A.K. RNA Interference-Based Therapy and Its Delivery Systems. Cancer Metastasis. Rev. 2018, 37, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Hattab, D.; Bakhtiar, A. Bioengineered SiRNA-Based Nanoplatforms Targeting Molecular Signaling Pathways for the Treatment of Triple Negative Breast Cancer: Preclinical and Clinical Advancements. Pharmaceutics 2020, 12, 929. [Google Scholar] [CrossRef]
- Vaidya, A.M.; Sun, Z.; Ayat, N.; Schilb, A.; Liu, X.; Jiang, H.; Sun, D.; Scheidt, J.; Qian, V.; He, S.; et al. Systemic Delivery of Tumor-Targeting SiRNA Nanoparticles against an Oncogenic LncRNA Facilitates Effective Triple-Negative Breast Cancer Therapy. Bioconjugate Chem. 2019, 30, 907–919. [Google Scholar] [CrossRef]
- Wu, B.; Yuan, Y.; Han, X.; Wang, Q.; Shang, H.; Liang, X.; Jing, H.; Cheng, W. Structure of LINC00511-siRNA-conjugated Nanobubbles and Improvement of Cisplatin Sensitivity on Triple Negative Breast Cancer. FASEB J. 2020, 34, 9713–9726. [Google Scholar] [CrossRef]
- Bi, Z.; Li, Q.; Dinglin, X.; Xu, Y.; You, K.; Hong, H.; Hu, Q.; Zhang, W.; Li, C.; Tan, Y.; et al. Nanoparticles (NPs)-Meditated LncRNA AFAP1-AS1 Silencing to Block Wnt/β -Catenin Signaling Pathway for Synergistic Reversal of Radioresistance and Effective Cancer Radiotherapy. Adv. Sci. 2020, 7, 2000915. [Google Scholar] [CrossRef]
- Jordan, N.J.; Dutkowski, C.M.; Barrow, D.; Mottram, H.J.; Hutcheson, I.R.; Nicholson, R.I.; Guichard, S.M.; Gee, J.M. Impact of Dual MTORC1/2 MTOR Kinase Inhibitor AZD8055 on Acquired Endocrine Resistance in Breast Cancer in Vitro. Breast Cancer Res. 2014, 16, R12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-L.; Overstreet, A.-M.; Chen, M.-S.; Wang, J.; Zhao, H.-J.; Ho, P.-C.; Smith, M.; Wang, S.-C. Combined Inhibition of EGFR and C-ABL Suppresses the Growth of Triple-Negative Breast Cancer Growth through Inhibition of HOTAIR. Oncotarget 2015, 6, 11150–11161. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Hu, Q.; Li, C.; Xing, Z.; Ma, G.; Wang, C.; Li, J.; Ye, Y.; Yao, J.; Liang, K.; et al. The LINK-A LncRNA Interacts with PtdIns(3,4,5)P3 to Hyperactivate AKT and Confer Resistance to AKT Inhibitors. Nat. Cell Biol. 2017, 19, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Ye, Y.; Chan, L.-C.; Li, Y.; Liang, K.; Lin, A.; Egranov, S.D.; Zhang, Y.; Xia, W.; Gong, J.; et al. Oncogenic LncRNA Downregulates Cancer Cell Antigen Presentation and Intrinsic Tumor Suppression. Nat. Immunol. 2019, 20, 835–851. [Google Scholar] [CrossRef] [PubMed]
- Shin, V.Y.; Chen, J.; Cheuk, I.W.-Y.; Siu, M.-T.; Ho, C.-W.; Wang, X.; Jin, H.; Kwong, A. Long Non-Coding RNA NEAT1 Confers Oncogenic Role in Triple-Negative Breast Cancer through Modulating Chemoresistance and Cancer Stemness. Cell Death Dis. 2019, 10, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooding, A.J.; Parker, K.A.; Valadkhan, S.; Schiemann, W.P. The LncRNA BORG: A Novel Inducer of TNBC Metastasis, Chemoresistance, and Disease Recurrence. J. Cancer Metastasis Treat. 2019, 5, 41. [Google Scholar] [CrossRef]
- Zhang, Y.; He, Q.; Hu, Z.; Feng, Y.; Fan, L.; Tang, Z.; Yuan, J.; Shan, W.; Li, C.; Hu, X.; et al. Long Noncoding RNA LINP1 Regulates Repair of DNA Double-Strand Breaks in Triple-Negative Breast Cancer. Nat. Struct. Mol. Biol. 2016, 23, 522–530. [Google Scholar] [CrossRef]
- Eades, G.; Wolfson, B.; Zhang, Y.; Li, Q.; Yao, Y.; Zhou, Q. LincRNA-RoR and MiR-145 Regulate Invasion in Triple-Negative Breast Cancer via Targeting ARF6. Mol. Cancer Res. 2015, 13, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Mitobe, Y.; Ikeda, K.; Suzuki, T.; Takagi, K.; Kawabata, H.; Horie-Inoue, K.; Inoue, S. ESR1-Stabilizing Long Noncoding RNA TMPO-AS1 Promotes Hormone-Refractory Breast Cancer Progression. Mol. Cell. Biol. 2019, 39, e00261-19. [Google Scholar] [CrossRef]
- Tang, L.; Chen, Y.; Chen, H.; Jiang, P.; Yan, L.; Mo, D.; Tang, X.; Yan, F. DCST1-AS1 Promotes TGF-β-Induced Epithelial–Mesenchymal Transition and Enhances Chemoresistance in Triple-Negative Breast Cancer Cells via ANXA1. Front. Oncol. 2020, 10, 280. [Google Scholar] [CrossRef]
- Wang, X.; Li, S.; Xiao, H.; Deng, X. Serum LncRNA TINCR Serve as a Novel Biomarker for Predicting the Prognosis in Triple-Negative Breast Cancer. Technol. Cancer Res. Treat. 2020, 19, 153303382096557. [Google Scholar] [CrossRef]
- Wu, M.; Wen, L.; Zhou, Y.; Wu, W. Role of LncRNA AGAP2-AS1 in Breast Cancer Cell Resistance to Apoptosis by the Regulation of MTA1 Promoter Activity. Technol. Cancer Res. Treat. 2022, 21, 153303382210853. [Google Scholar] [CrossRef]
- Hsu, M.-Y.; Hsieh, C.-H.; Huang, Y.-T.; Chu, S.-Y.; Chen, C.-M.; Lee, W.-J.; Liu, S.-J. Enhanced Paclitaxel Efficacy to Suppress Triple-Negative Breast Cancer Progression Using Metronomic Chemotherapy with a Controlled Release System of Electrospun Poly-d-l-Lactide-Co-Glycolide (PLGA) Nanofibers. Cancers 2021, 13, 3350. [Google Scholar] [CrossRef]
- Narui, K.; Ishikawa, T.; Shimizu, D.; Yamada, A.; Tanabe, M.; Sasaki, T.; Oba, M.S.; Morita, S.; Nawata, S.; Kida, K.; et al. Anthracycline Could Be Essential for Triple-Negative Breast Cancer: A Randomised Phase II Study by the Kanagawa Breast Oncology Group (KBOG) 1101. Breast 2019, 47, 1–9. [Google Scholar] [CrossRef]
- Xu, J.; Wu, K.; Jia, Q.; Ding, X. Roles of MiRNA and IncRNA in Triple-Negative Breast Cancer. J. Zhejiang Univ. Sci. B 2020, 21, 673–689. [Google Scholar] [CrossRef]
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative Approaches for Cancer Treatment: Current Perspectives and New Challenges. Ecancermedicalscience 2019, 13, 961. [Google Scholar] [CrossRef]
- Huang, L.; Liang, G.; Zhang, Q.; Zhao, W. The Role of Long Noncoding RNAs in Antiestrogen Resistance in Breast Cancer: An Overview and Update. J. Breast Cancer 2020, 23, 129. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.D.; Verma, R.; Tripathi, S.K.; Sahu, R.; Trivedi, P.; Yadav, D.K. Breast Cancer Transcriptional Regulatory Network Reprogramming by using the CRISPR/Cas9 System: An Oncogenetics Perspective. Curr. Top Med. Chem. 2021, 21, 2800–2813. [Google Scholar] [CrossRef]
- Singh, D.D.; Hawkins, R.D.; Lahesmaa, R.; Tripathi, S.K. CRISPR/Cas9 guided genome and epigenome engineering and its therapeutic applications in immune mediated diseases. Semin. Cell Dev. Biol. 2019, 96, 32–43. [Google Scholar] [CrossRef]
- Chen, B.; Dragomir, M.P.; Yang, C.; Li, Q.; Horst, D.; Calin, G.A. Targeting Non-Coding RNAs to Overcome Cancer Therapy Resistance. Sig. Transduct. Target. Ther. 2022, 7, 121. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. N. | lncRNAs | Regulation of Expression | Clinical Importance | Potential Targets | Reference |
|---|---|---|---|---|---|
| 1 | HOTAIR | Upregulation | Increase cell invasion and migration | LEF1/TCF4 | [31] |
| 2 | DRHC | Downregulation | Reduce cell proliferation | HOTAIR | [32] |
| 3 | LINC01133 | Upregulation | Promote phenotypic features like cell stem cells (CSCs) | KLF4 | [33] |
| 4 | LINC01096 | Upregulation | Encourage cell invasion | miR-3130-3p | [34] |
| 5 | HEIH | Upregulation | Increase cell proliferation and prevent cell death | miR-4458/SOCS1 | [35] |
| 6 | ARNILA | Downregulation | Invasion and metastasis | miR-204/SOX4 | [36] |
| 7 | LINC02095 | Upregulation | Promote cell proliferation | SOX9 | [37] |
| 8 | WT1-AS | Downregulation | Inhibit cell migration and invasion | TGF-β1 | [38] |
| 9 | GAS5 | Downregulation | Promote cell apoptosis | miR-378a-5p/SUFU | [39] |
| 10 | CCAT1 | Upregulation | Encourage cell division | miR-218/ZFX | [40] |
| 11 | ASRPS | Downregulation | Inhibit angiogenesis and cell proliferation | STAT3 | [41] |
| 12 | AND2-AS1 | Downregulation | Inhibit angiogenesis inhibit cell division | RUNX2 | [42] |
| 13 | POU3F3 | Upregulation | Promote cell proliferation and inhibit cell apoptosis | Caspase-9 | [43] |
| 14 | NEF | Downregulation | Inhibit cell migration and invasion | miR-155 | [44] |
| 15 | ZEB2-AS1 | Upregulation | Promote cell proliferation, metastasis, and EMT | ZEB2 | [45] |
| 16 | LINC0009 | Upregulation | Increase cell proliferation and invasion | miR-383-5p/RBM3 | [46] |
| 17 | ANRIL | Upregulation | Increase cell proliferation and apoptosis | miR-448/KDM5B | [47] |
| 18 | SNHG12 | Upregulation | Induce cell proliferation, migration, and apoptosis | MMP13 | [48] |
| 19 | LUCAT1 | Upregulation | Encourage cell division, movement, and invasion | miR-5702 | [49] |
| 20 | PCAT6 | Upregulation | Radiotherapy resistance | miR-185-5p/TPD52 | [50] |
| 22 | HULC | Upregulation | Promote metastasis | MMP-2, MMP-9 | [51] |
| 23 | PAPAS | Upregulation | Induce cell migration and invasion | miR-34a | [52] |
| 24 | HCP5 | Upregulation | Increase cell proliferation; reduce cell apoptosis | miR-219a-5p/BIRC3 | [53] |
| 25 | NRAD1 | Upregulation | Stimulate cell proliferation and CSC-like phenotypic traits | miR-219a-5p/BIRC3 | [54] |
| 26 | SNAR | Upregulation | Stimulate cell division | [55] | |
| 27 | AWPPH | Upregulation | Activate cell proliferation | miR-21; FZD7 | [56] |
| 28 | sONE | Downregulation | Prevent cell proliferation | TP53/c-Myc | [57] |
| 29 | DANCR | Upregulation | Promote cell proliferation and invasion | miR-216a-5p | [58] |
| 30 | LINK-A | Upregulation | Increase resistance to immunotherapy, AKT inhibitors, and glycolysis reprogramming | PI3K/GPCR | [59] |
| 31 | MIR503HG | Downregulation | Reduce cell migration and invasion | miR-103/OLFM4 | [60] |
| 32 | NEAT1 | Upregulation | Increase cell apoptosis | [61] | |
| 33 | PTCSC3 | Downregulation | Prevent cell proliferation | H19 | [62] |
| 34 | NRON | Downregulation | Inhibit cell proliferation | snaR | [63] |
| 35 | TROJAN | Upregulation | Promote cell proliferation and invasion | ZMYND8 | [64] |
| 36 | NAMPT-AS | Upregulation | Increase cell metastasis | miR-548b-3p/NAMPT | [14] |
| 37 | MANCR | Upregulation | Promote cell proliferation; inhibit DNA damage | [65] | |
| 38 | RMST | Downregulation | Prevent cell proliferation | [66] | |
| 39 | SK AI1BC | Upregulation | Increase cell migration and invasion | K AI1 | [67] |
| 40 | ROR | Upregulation | Promote cell invasion and metastasis | miR-145/ARF6 | [68] |
| 41 | AIRN | Downregulation | Inhibit cell migration and invasion | Wnt/β-catenin/mTOR/PI3K | [69] |
| 42 | LINC-ZNF469-3 | Upregulation | Promote cell invasion | miR-574-5p/ZEB1 | [70] |
| 43 | PDCD4-AS1 | Downregulation | Inhibit cell proliferation and migration | PDCD4 | [71] |
| 44 | HOST2 | Downregulation | Inhibit cell proliferation | et-7 b/CDK6 | [72] |
| 45 | BORG | Upregulation | Promote doxorubicin resistance | RPA1 | [73] |
| 46 | PVT1 | Upregulation | Promote cell proliferation and migration, and EMT | p21, KLF5/β-catenin | [24] |
| 47 | H19 | Upregulation | Promote paclitaxel resistance and CSC-like phenotypic traits | Akt | [62] |
| 48 | TP73-AS1 | Downregulation | Promote cell vasculogenic mimicry | miR-490-3p/TWIST1 | [74] |
| 49 | TUG1 | Downregulation | Enhance cisplatin sensitivity | miR-197/NLK | [75] |
| 50 | MIR100HG | Upregulation | Promote cell proliferation | p27 | [76] |
| 51 | LINC01638 | Upregulation | Promote cell proliferation | c-Myc | [77] |
| S.N. | lncRNA | Expression Patterns | Pathway/Target | Expression Pattern Drugs | Reference |
|---|---|---|---|---|---|
| 1 | H19 | Upregulation | H19/Let-7/LIN28 axis | Anthracycline, paclitaxel, fulverstrant, doxorubicin tamoxifen | [19] |
| 2 | NEAT1 | Upregulation | miR-211/HMGA2 | Paclitaxel, 5-FU, cisplatin | [61] |
| 3 | GAS5 | Downregulation | miR-21/mTOR/ | Adriamycin, trastuzumab, tamoxifen, paclitaxel | [91] |
| 4 | LINK-A | Upregulation | PIP3/GPCR/cAMP/PKA/ TRIM71/PLC | Immune checkpoint blockers | [101] |
| 5 | UCA1 | Upregulation | Wnt/b-catenin signalling | Trastuzumab, tamoxifen | [89] |
| 6 | LINP1 | Upregulation | Caspase-9/Bax | Doxorubicin, 5-FU, tamoxifen | [105] |
| 7 | Linc-ROR | Upregulation | miR-194-3p/MECP2 | Tamoxifen, paclitaxel, 5-FU | [106] |
| 8 | TMPO-AS1 | Upregulation | ER | Endocrine therapy | [107] |
| 9 | DCST1-AS1 | Upregulation | ANXA1 | Doxorubicin, paclitaxel | [108] |
| 10 | TINCR | Upregulation | miR-125b/ERBB2 | Trastuzumab | [109] |
| 11 | HOTAIR | Upregulation | ER | Tamoxifen, doxorubicin, trastuzumab, | [100] |
| 12 | AFAP1-AS1 | Upregulation | AUF1/ERBB2 | Trastuzumab | [98] |
| 13 | AGAP2-AS1 | Upregulation | CBP/ MyD88/H3K27/NF-kB | Trastuzumab | [110] |
| 14 | AK124454 | Upregulation | Paclitaxel | [111] | |
| 15 | NONHSAT057282 | Upregulation | ELF1 and E2F1 | Anthracycline | [112] |
| 16 | NONHSAG023333 | Upregulation | ELF1 and E2F1 | Anthracycline | [112] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, D.D.; Lee, H.-J.; Yadav, D.K. Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer. Cells 2023, 12, 674. https://doi.org/10.3390/cells12040674
Singh DD, Lee H-J, Yadav DK. Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer. Cells. 2023; 12(4):674. https://doi.org/10.3390/cells12040674
Chicago/Turabian StyleSingh, Desh Deepak, Hae-Jeung Lee, and Dharmendra Kumar Yadav. 2023. "Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer" Cells 12, no. 4: 674. https://doi.org/10.3390/cells12040674
APA StyleSingh, D. D., Lee, H. -J., & Yadav, D. K. (2023). Recent Clinical Advances on Long Non-Coding RNAs in Triple-Negative Breast Cancer. Cells, 12(4), 674. https://doi.org/10.3390/cells12040674