
Decoy Receptors Regulation by Resveratrol in Lipopolysaccharide-Activated Microglia


 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Cytotoxicity Assay
2.3. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) and End-Point PCR
2.4. Electrophoresis
2.5. Western Blotting
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. NO Production
2.8. PGE2 Assay
2.9. Statistical Analysis
3. Results and Discussion
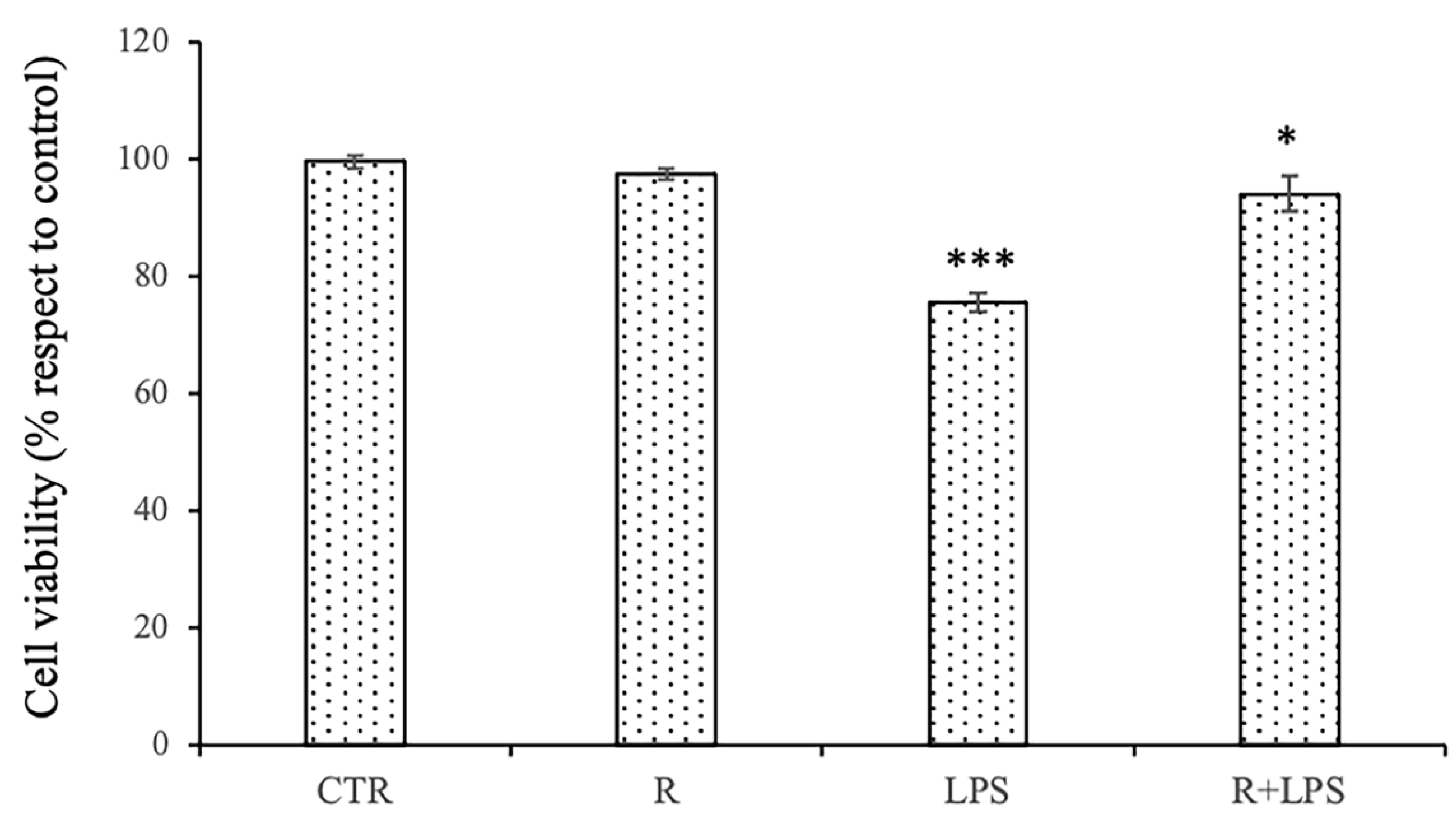
3.1. Effects of Resveratrol and Prolonged LPS Treatment on Cell Viability of N13 Microglial Cells
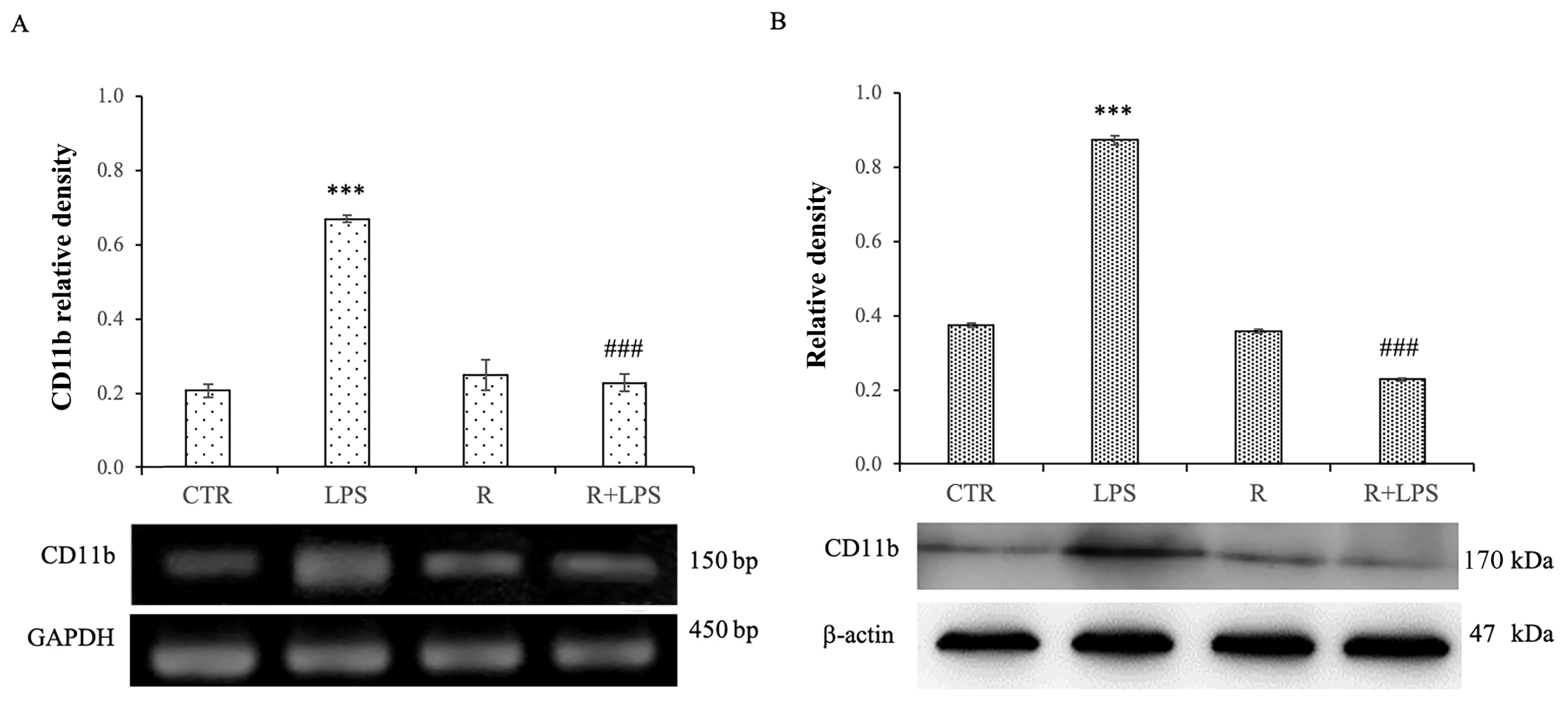
3.2. Resveratrol Modulates CD11b Expression Levels in LPS-Treated N13 Microglial Cells
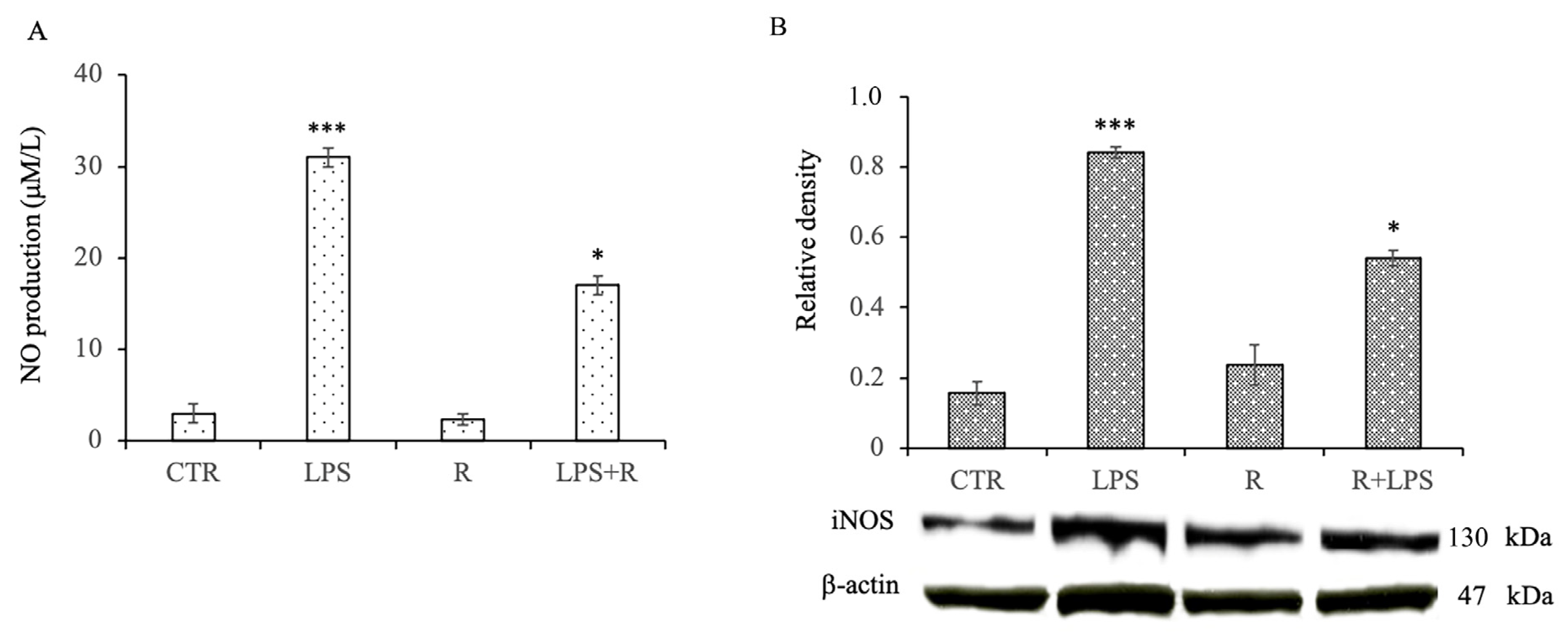
3.3. Effects of Resveratrol on Nitric Oxide Production and Inducible Nitric Oxide Synthase Protein Expression Levels in LPS-Treated N13 Microglial Cells
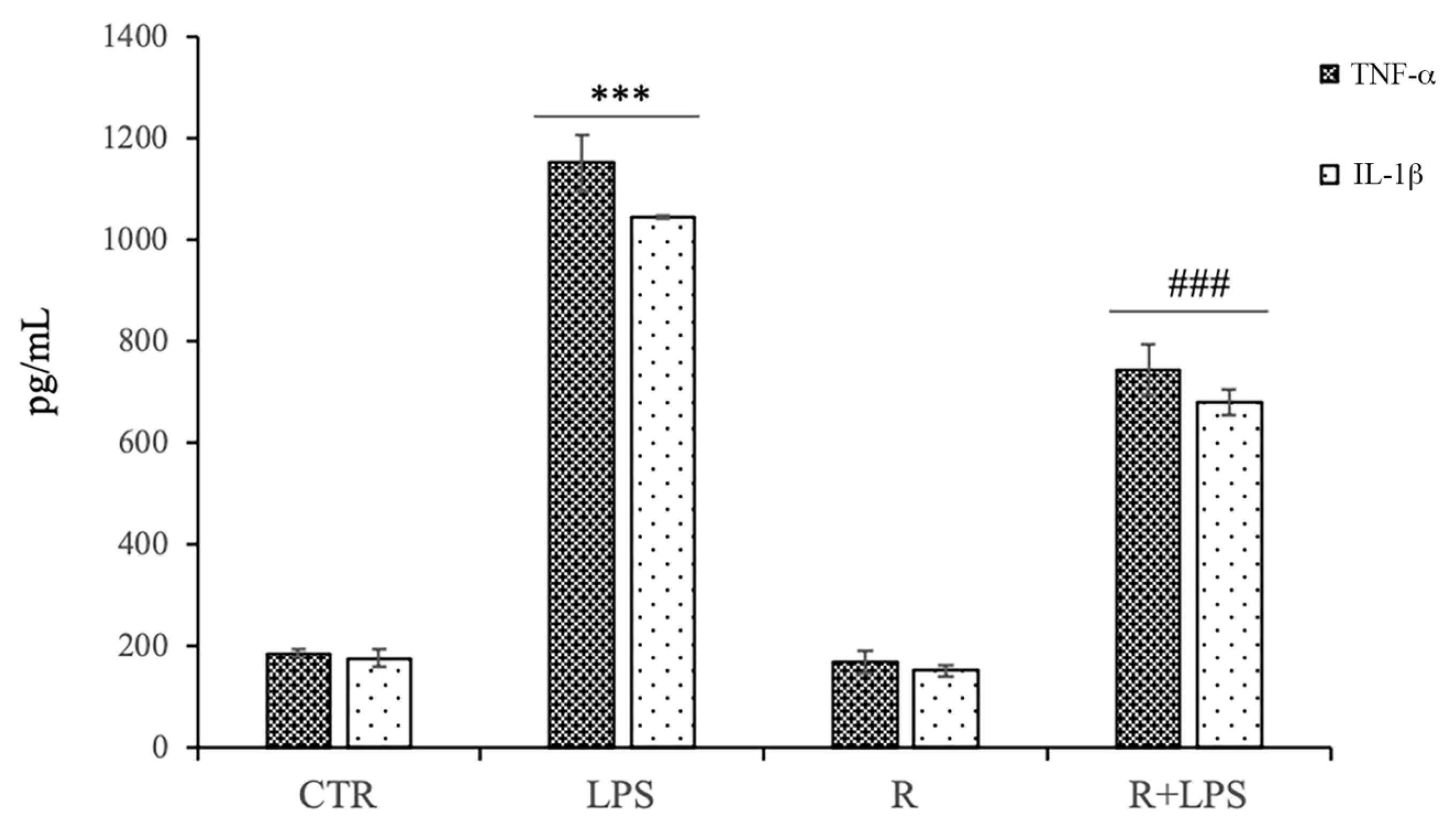
3.4. Effects of Resveratrol on Pro-Inflammatory Cytokine Production in LPS-Treated N13 Microglial Cells
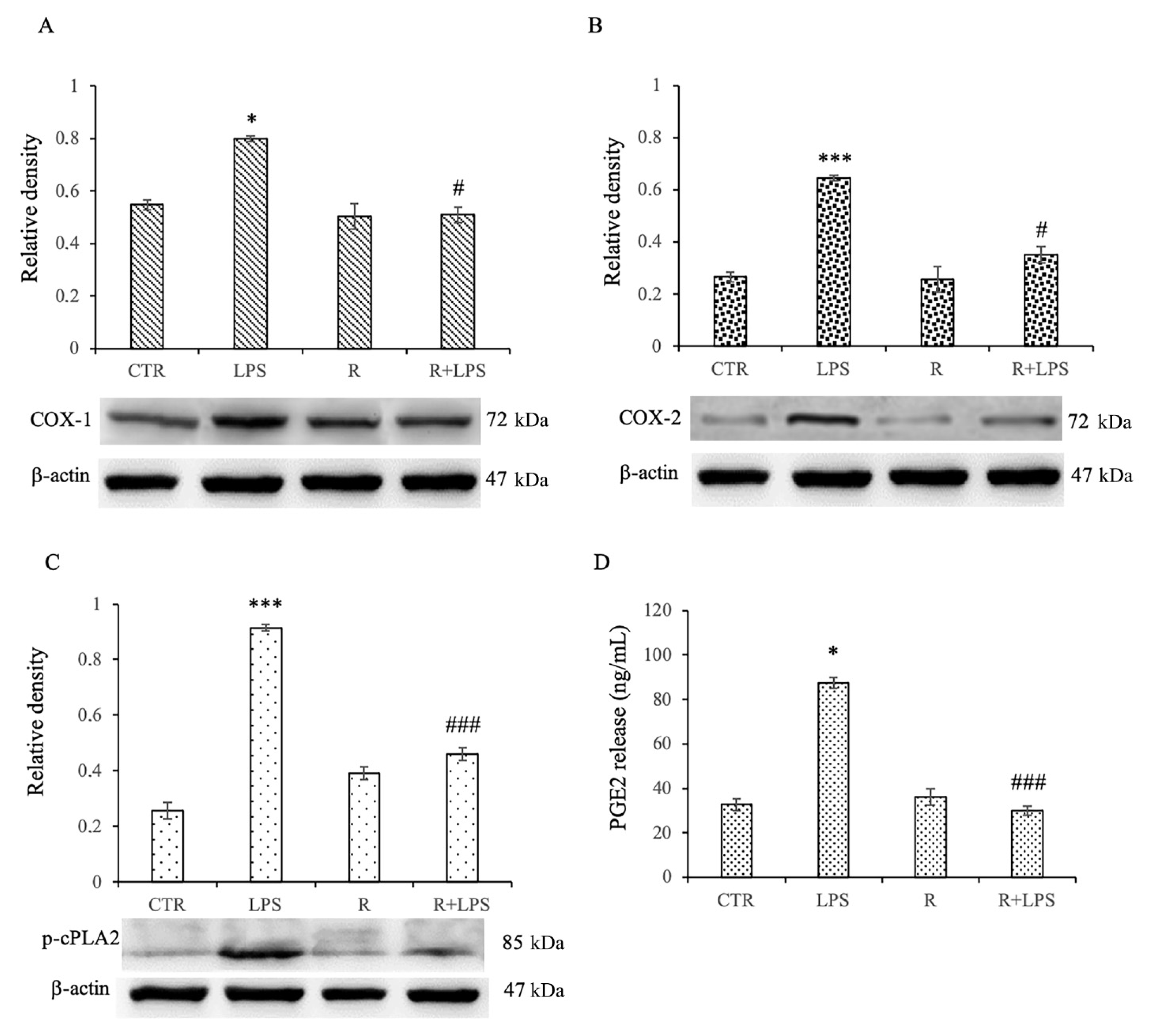
3.5. Effects of Resveratrol on Arachidonic Acid (AA) Pathway in LPS-Treated Microglia
3.6. Effects of Resveratrol on NF-kB Pathway in LPS-Treated N13 Microglial Cells
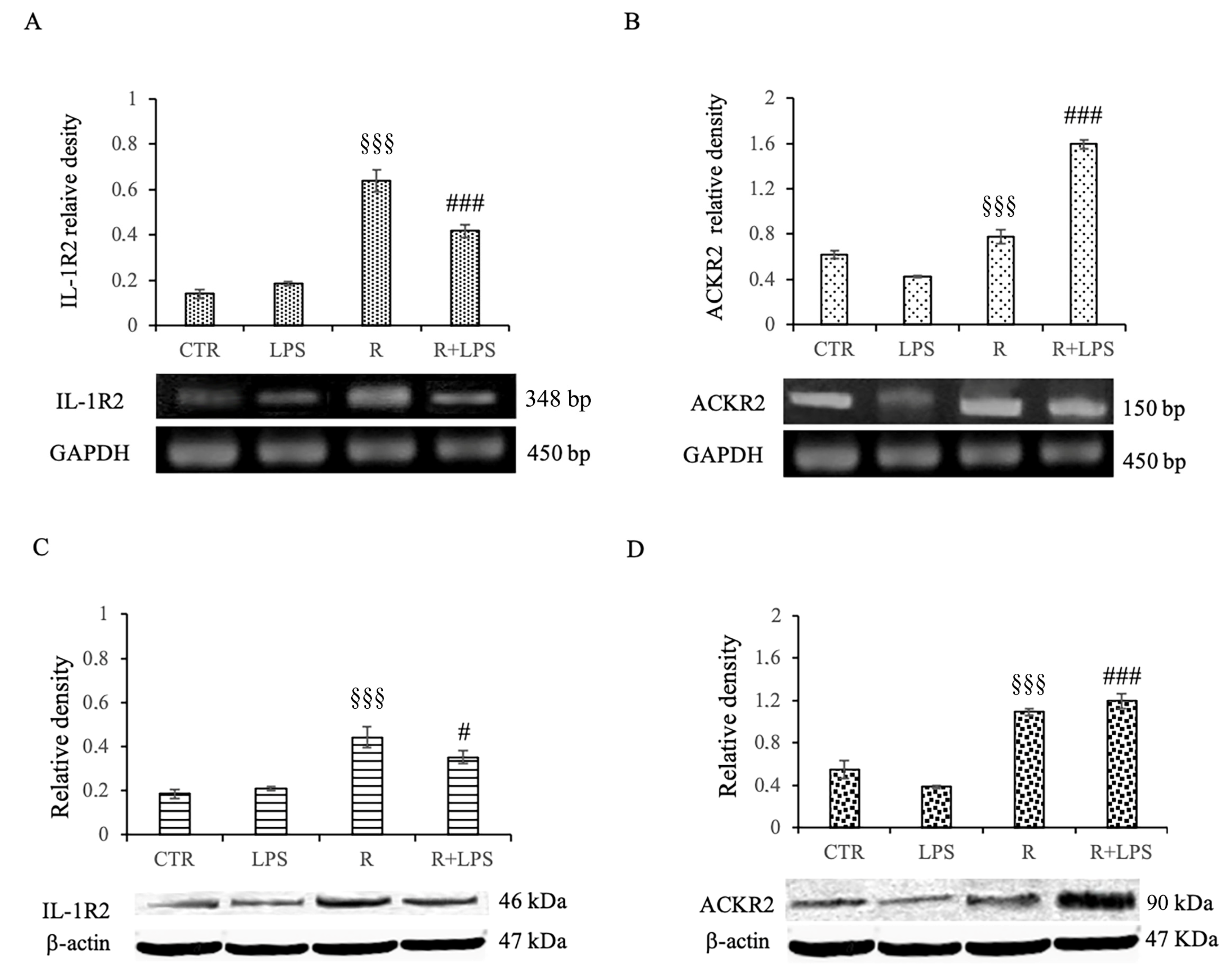
3.7. Effects of Resveratrol on IL1-R2 and ACKR2 Decoy Receptor Expression in LPS-Treated Microglia
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, L.X.; Li, C.X.; Kakar, M.U.; Khan, M.S.; Wu, P.F.; Amir, R.M.; Dai, D.F.; Naveed, M.; Li, Q.Y.; Saeed, M.; et al. Resveratrol (RV): A pharmacological review and call for further research. Biomed Pharm. 2021, 143, 112164. [Google Scholar] [CrossRef] [PubMed]
- Orallo, F. Comparative studies of the antioxidant effects of cis- and trans-resveratrol. Curr. Med. Chem. 2006, 13, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Porro, C.; Cianciulli, A.; Calvello, R.; Panaro, M.A. Reviewing the Role of Resveratrol as a Natural Modulator of Microglial Activities. Curr. Pharm. Des. 2015, 21, 5277–5529. [Google Scholar] [CrossRef] [PubMed]
- Shih, M.K.; Tain, Y.L.; Chen, Y.W.; Hsu, W.H.; Yeh, Y.T.; Chang, S.K.C.; Liao, J.X.; Hou, C.Y. Resveratrol Butyrate Esters Inhibit Obesity Caused by Perinatal Exposure to Bisphenol A in Female Offspring Rats. Molecules 2021, 26, 4010. [Google Scholar] [CrossRef]
- Su, M.; Zhao, W.; Xu, S.; Weng, J. Resveratrol in Treating Diabetes and Its Cardiovascular Complications: A Review of Its Mechanisms of Action. Antioxidants 2022, 11, 1085. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Y.; Jin, S.; Pang, Q.; Shan, A.; Feng, X. Dietary resveratrol alleviated lipopolysaccharide-induced ileitis through Nrf2 and NF-kappaB signaling pathways in ducks (Anas platyrhynchos). J. Anim. Physiol. Anim. Nutr. 2022, 106, 1306–1320. [Google Scholar] [CrossRef]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef]
- Kiskova, T.; Kubatka, P.; Büsselberg, D.; Kassayova, M. The Plant-Derived Compound Resveratrol in Brain Cancer: A Review. Biomolecules 2020, 10, 161. [Google Scholar] [CrossRef] [Green Version]
- Wąsik, A.; Antkiewicz-Michaluk, L. The mechanism of neuroprotective action of natural compounds. Pharmacol. Rep. 2017, 69, 851–860. [Google Scholar] [CrossRef]
- Rahman, M.H.; Bajgai, J.; Fadriquela, A.; Sharma, S.; Thi, T.T.; Akter, R.; Goh, S.H.; Kim, C.S.; Lee, K.J. Redox effects of molecular hydrogen and its therapeutic efficacy in the treatment of neurodegenerative diseases. Processes 2021, 9, 308. [Google Scholar] [CrossRef]
- Cione, E.; La Torre, C.; Cannataro, R.; Caroleo, M.C.; Plastina, P.; Gallelli, L. Quercetin, Epigallocatechin Gallate, Curcumin, and Resveratrol: From Dietary Sources to Human MicroRNA Modulation. Molecules 2019, 25, 63. [Google Scholar] [CrossRef] [Green Version]
- Viegas, F.P.D.; Gontijo, V.S.; de Freitas Silva, M.; Ortiz, C.J.C.; Dos Reis Rosa Franco, G.; Ernesto, J.T.; Damasio, C.M.; Silva, I.M.F.; Campos, T.G.; Viegas, C. Curcumin, Resveratrol and Cannabidiol as Natural Key Prototypes in Drug Design for Neuroprotective Agents. Curr. Neuropharmacol. 2022, 20, 1297–1328. [Google Scholar] [CrossRef] [PubMed]
- Benameur, T.; Giacomucci, G.; Panaro, M.A.; Ruggiero, M.; Trotta, T.; Monda, V.; Pizzolorusso, I.; Lofrumento, D.D.; Porro, C.; Messina, G. New Promising Therapeutic Avenues of Curcumin in Brain Diseases. Molecules. 2021, 27, 236. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Surh, Y.J. Protective effect of resveratrol on β-amyloid-induced oxidative PC12 cell death. Free Radic. Biol. Med. 2003, 34, 1100–1110. [Google Scholar] [CrossRef]
- Lofrumento, D.D.; Nicolardi, G.; Cianciulli, A.; De Nuccio, F.; La Pesa, V.; Carofiglio, V.; Dragone, T.; Calvello, R.; Panaro, M.A. Neuroprotective effects of resveratrol in an MPTP mouse model of Parkinson’s-like disease: Possible role of SOCS-1 in reducing pro-inflammatory responses. Innate. Immun. 2014, 20, 249–260. [Google Scholar] [CrossRef]
- Supino, D.; Minute, L.; Mariancini, A.; Riva, F.; Magrini, E.; Garlanda, C. Negative Regulation of the IL-1 System by IL-1R2 and IL-1R8: Relevance in Pathophysiology and Disease. Front. Immunol. 2022, 13, 804641. [Google Scholar] [CrossRef] [PubMed]
- Graham, G.J.; Locati, M.; Mantovani, A.; Rot, A.; Telen, M. The biochemistry and biology of the atypical chemokine receptors. Immunol. Lett. 2012, 145, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Bonecchi, R.; Graham, G.J. Atypical Chemokine Receptors and Their Roles in the Resolution of the Inflammatory Response. Front. Immunol. 2016, 7, 224. [Google Scholar] [CrossRef] [Green Version]
- Peters, V.A.; Joesting, J.J.; Freund, G.G. IL-1 receptor 2 (IL-1R2) and its role in immune regulation. Brain Behav. Immun. 2013, 32, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shabgah, A.G.; Jadidi-Niaragh, F.; Mohammadi, H.; Ebrahimzadeh, F.; Oveisee, M.; Jahanara, A.; Navashenaq, J.G. The Role of Atypical Chemokine Receptor D6 (ACKR2) in Physiological and Pathological Conditions; Friend, Foe, or Both? Front. Immunol. 2022, 13, 3861931. [Google Scholar]
- Cianciulli, A.; Dragone, T.; Calvello, R.; Porro, C.; Trotta, T.; Lofrumento, D.D.; Panaro, M.A. IL-10 plays a pivotal role in anti-inflammatory effects of resveratrol in activated microglia cells. Int. Immunopharmacol. 2015, 24, 369–376. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dragone, T.; Cianciulli, A.; Calvello, R.; Porro, C.; Trotta, T.; Panaro, M.A. Resveratrol counteracts lipopolysaccharide-mediated microglial inflammation by modulating a SOCS-1 dependent signaling pathway. Toxicol. Vitr. 2014, 28, 1126–1135. [Google Scholar] [CrossRef]
- Bartels, A.L.; Leenders, K.L. Cyclooxygenase and neuroinflammation in Parkinson’s disease neurodegeneration. Curr. Neuropharmacol. 2010, 8, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Aïd, S.; Bosetti, F. Targeting cyclooxygenases-1 and -2 in neuroinflammation: Therapeutic implications. Biochimie 2011, 93, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [Green Version]
- Capiralla, H.; Vingtdeux, V.; Zhao, H.; Sankowski, R.; Al-Abed, Y.; Davies, P.; Marambaud, P. Resveratrol mitigates lipopolysaccharide- and Aβ-mediated microglial inflammation by inhibiting the TLR4/NF-κB/STAT signaling cascade. J. Neurochem. 2012, 120, 461–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truong, V.L.; Jun, M.; Jeong, W.S. Role of resveratrol in regulation of cellular defense systems against oxidative stress. Biofactors 2018, 44, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, R.; Chatterjee, S.; Hembram, K.C.; Sethy, C.; Mandal, M.; Kundu, C.N. Nano formulated Resveratrol inhibits metastasis and angiogenesis by reducing inflammatory cytokines in oral cancer cells by targeting tumor associated macrophages. J. Nutr. Biochem. 2021, 92, 108624. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.L.; DeWitt, D.L.; Garavito, R.M. Cyclooxygenases: Structural, cellular, and molecular biology. Annu. Rev. Biochem. 2000, 69, 145–182. [Google Scholar] [CrossRef] [Green Version]
- Calvello, R.; Panaro, M.A.; Carbone, M.L.; Cianciulli, A.; Perrone, M.G.; Vitale, P.; Malerba, P.; Scilimati, A. Novel selective COX-1 inhibitors suppress neuroinflammatory mediators in LPS-stimulated N13 microglial cells. Pharmacol. Res. 2012, 65, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Xu, J.; Jensen, M.D.; Simonyi, A. Phospholipase A2 in the central nervous system: Implications for neurodegenerative diseases. J. Lipid. Res. 2004, 45, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, D.Y.; Simonyi, A.; Kotzbauer, P.T.; Gu, Z.; Sun, G.Y. Cytosolic phospholipase A2 plays a crucial role in ROS/NO signaling during microglial activation through the lipoxygenase pathway. J. Neuroinflammation 2015, 12, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyurin, V.A.; Tyurina, Y.Y.; Borisenko, G.G.; Sokolova, T.V.; Ritov, V.B.; Quinn, P.J.; Rose, M.; Kochanek, P.; Graham, S.H.; Kagan, V.E. Oxidative stress following traumatic brain injury in rats: Quantitation of biomarkers and detection of free radical intermediates. J. Neurochem. 2000, 75, 2178–2189. [Google Scholar] [CrossRef] [Green Version]
- Akundi, R.S.; Candelario-Jalil, E.; Hess, S.; Hüll, M.; Lieb, K.; Gebicke-Haerter, P.J.; Fiebich, B.L. Signal transduction pathways regulating cyclooxygenase-2 in lipopolysaccharide-activated primary rat microglia. Glia 2005, 51, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.A.; Kim, G.Y.; Park, K.Y.; Choi, Y.H. Resveratrol inhibits nitric oxide and prostaglandin E2 production by lipopolysaccharide-activated C6 microglia. J. Med. Food. 2007, 10, 218–224. [Google Scholar] [CrossRef]
- Pinteaux, E.; Parker, L.C.; Rothwell, N.J.; Luheshi, G.N. Expression of Interleukin1 Receptors and Their Role in Interleukin-1 Actions in Murine Microglial Cells. J. Neurochem. 2002, 83, 754–763. [Google Scholar] [CrossRef]
- McNamee, E.N.; Ryan, K.M.; Kilroy, D.; Connor, T.J. Noradrenaline Induces IL1ra and IL-1 Type II Receptor Expression in Primary Glial Cells and Protects Against IL-1beta-Induced Neurotoxicity. Eur. J. Pharmacol. 2010, 626, 219–228. [Google Scholar] [CrossRef]
- Docagne, F.; Campbell, S.J.; Bristow, A.F.; Poole, S.; Vigues, S.; Guaza, C.; Perry, V.H.; Anthony, D.C. Differential Regulation of Type I and Type II Interleukin-1 Receptors in Focal Brain Inflammation. Eur. J. Neurosci. 2005, 21, 1205–1214. [Google Scholar] [CrossRef]
- Bachelerie, F.; Graham, G.J.; Locati, M.; Mantovani, A.; Murphy, P.M.; Nibbs, R.; Rot, A.; Sozzani, S.; Thelen, M. New nomenclature for atypical chemokine receptors. Nat. Immunol. 2014, 15, 207–208. [Google Scholar] [CrossRef]
- Boraschi, D.; Italiani, P.; Weil, S.; Martin, M.U. The family of the interleukin-1 receptors. Immunol. Rev. 2018, 281, 197–232. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Sequence References | Product |
|---|---|---|---|
| CD11b forward | 5′-GACTCAGTGAGCCCCATCAT-3′ | EF101557.1 | 150 bp |
| CD11b reverse | 5′-AGATCGTCTTGGCAGATGCT-3′ | ||
| ACKR2 forward | 5′-CCAGCCAAGCCCAGCACGAA-3′ | NM_001276719.1 | 150 bp |
| ACKR2 reverse | 5′-TGCCTCTCACCACCGTCGGT-3′ | ||
| IL-1R2 forward | 5′-CTCTTGGATAAAGGCATAAGGAATT-3′ | NM_001360800.1 | 348 bp |
| IL-1R2 reverse | 5′-CACACGGCCTCTTGGGTAAGCAGCC-3′ | ||
| GADPH forward | 5′-ACCACAGTCCATGCCATCAC-3′ | NM_001411843.1 | 450 bp |
| GADPH reverse | 5′-TCCACCACCCTGTTGCTGTA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvello, R.; Porro, C.; Lofrumento, D.D.; Ruggiero, M.; Panaro, M.A.; Cianciulli, A. Decoy Receptors Regulation by Resveratrol in Lipopolysaccharide-Activated Microglia. Cells 2023, 12, 681. https://doi.org/10.3390/cells12050681
Calvello R, Porro C, Lofrumento DD, Ruggiero M, Panaro MA, Cianciulli A. Decoy Receptors Regulation by Resveratrol in Lipopolysaccharide-Activated Microglia. Cells. 2023; 12(5):681. https://doi.org/10.3390/cells12050681
Chicago/Turabian StyleCalvello, Rosa, Chiara Porro, Dario Domenico Lofrumento, Melania Ruggiero, Maria Antonietta Panaro, and Antonia Cianciulli. 2023. "Decoy Receptors Regulation by Resveratrol in Lipopolysaccharide-Activated Microglia" Cells 12, no. 5: 681. https://doi.org/10.3390/cells12050681
APA StyleCalvello, R., Porro, C., Lofrumento, D. D., Ruggiero, M., Panaro, M. A., & Cianciulli, A. (2023). Decoy Receptors Regulation by Resveratrol in Lipopolysaccharide-Activated Microglia. Cells, 12(5), 681. https://doi.org/10.3390/cells12050681