

Maternal and Intrauterine Influences on Feto-Placental Growth Are Accompanied by Sexually Dimorphic Changes in Placental Mitochondrial Respiration, and Metabolic Signalling Pathways


 and
and 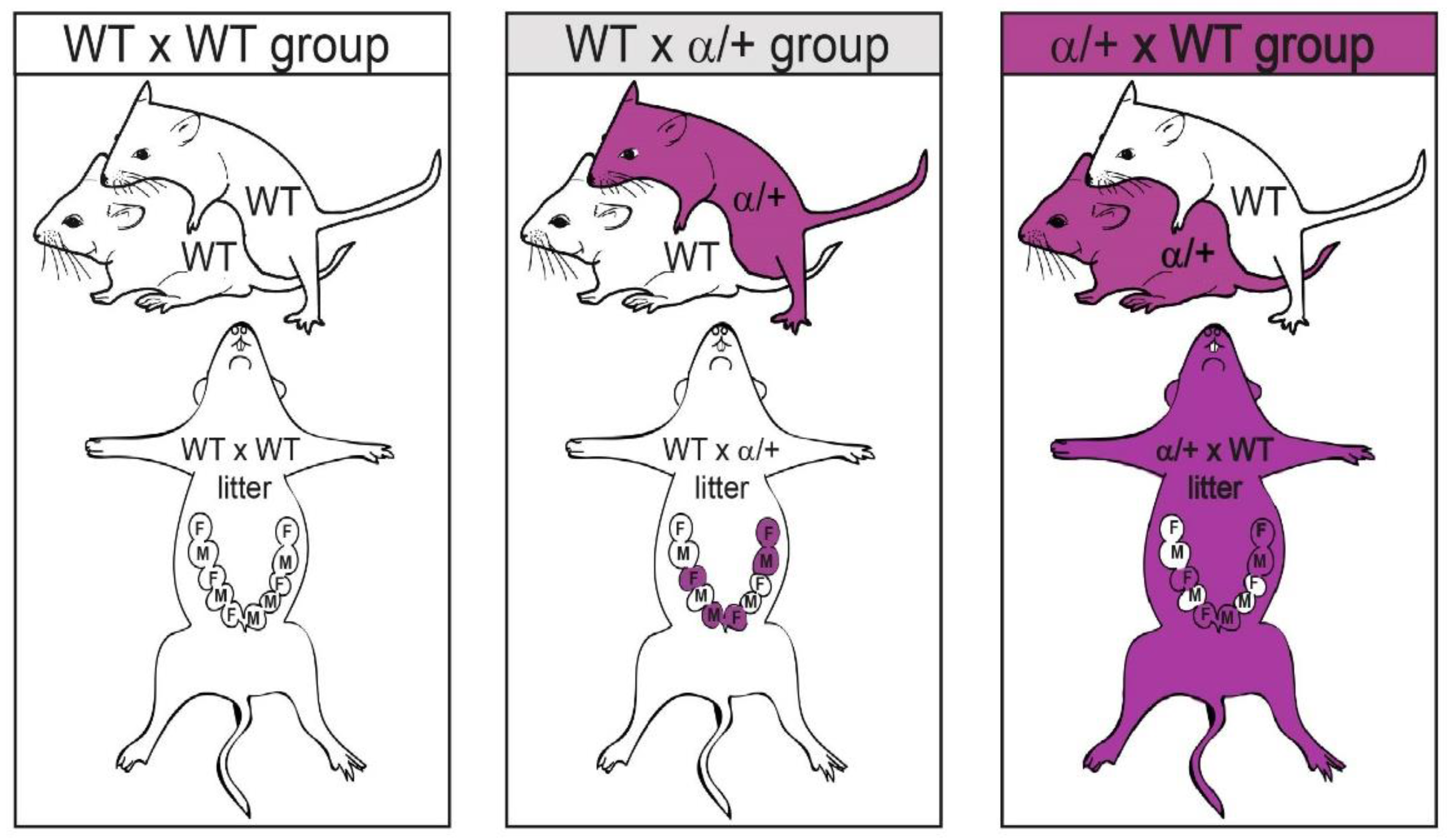{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Tissue Collection and Genotyping
2.3. Placental LZ Mitochondrial Respirometry
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
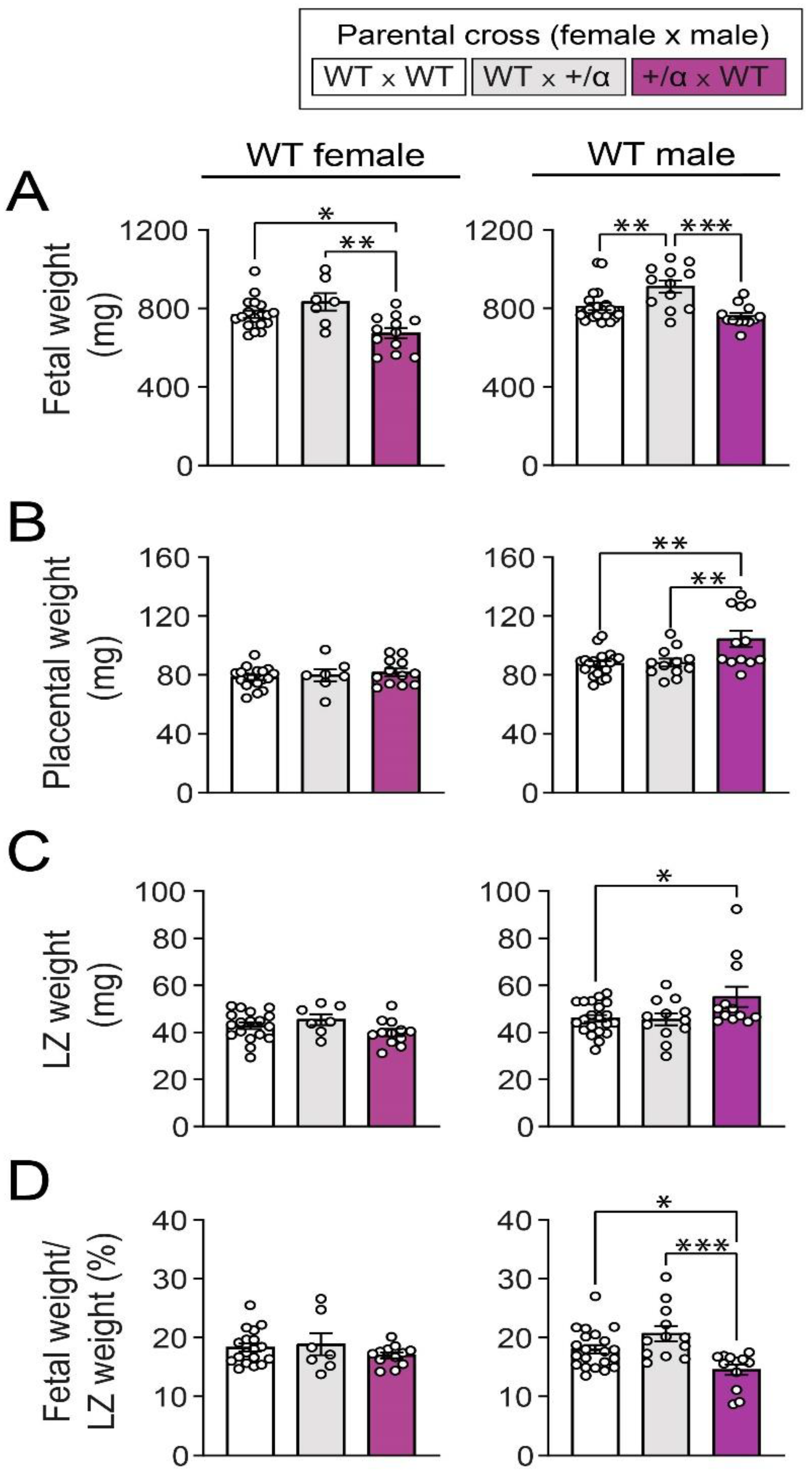
3.1. Littermate and/or Maternal p110α Deficiency Alters Feto-Placental Growth of WT Fetuses in a Sex-Specific Manner
3.2. Littermate and/or Maternal p110α Deficiency Alters Mitochondrial Bioenergetics in the Placental LZ of WT Fetuses of Both Sexes
3.3. Littermate and/or Maternal p110α Deficiency Alters the Expression of Mitochondrial-Related Proteins in the Placental LZ of WT Fetuses in a Sex-Specific Manner
3.4. Littermate and/or Maternal p110α Deficiency Alters the Abundance of Key Growth and Metabolic Signalling Proteins in a Manner That Depends on Fetal Sex
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salazar-Petres, E.; Carvalho, D.P.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. Placental structure, function and mitochondrial phenotype relate to fetal size in each fetal sex in mice. Biol. Reprod. 2022, 106, 1292–1311. [Google Scholar] [CrossRef]
- Coan, P.M.; Angiolini, E.; Sandovici, I.; Burton, G.J.; Constância, M.; Fowden, A.L. Adaptations in placental nutrient transfer capacity to meet fetal growth demands depend on placental size in mice. J. Physiol. 2008, 586, 4567–4576. [Google Scholar] [CrossRef] [PubMed]
- Sferruzzi-Perri, A.N.; Lopez-Tello, J.; Salazar-Petres, E. Placental adaptations supporting fetal growth during normal and adverse gestational environments. Exp. Physiol. 2022, 108, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Fowden, A.L.; Burton, G.J. The placenta: A multifaceted, transient organ. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Regnault, T.R.H.; Barker, P.L.; Botting, K.J.; McMillen, I.C.; McMillan, C.M.; Roberts, C.T.; Morrison, J.L. Placental adaptations in growth restriction. Nutrients 2015, 7, 360–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandovici, I.; Hoelle, K.; Angiolini, E.; Constância, M. Placental adaptations to the maternal-fetal environment: Implications for fetal growth and developmental programming. Reprod. Biomed. Online 2012, 25, 68–89. [Google Scholar] [CrossRef] [Green Version]
- Napso, T.; Yong, H.E.J.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The role of placental hormones in mediating maternal adaptations to support pregnancy and lactation. Front. Physiol. 2018, 9, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.A. The endocrine function of human placenta: An overview. Reprod. Biomed. Online 2015, 32, 14–43. [Google Scholar] [CrossRef] [Green Version]
- Stern, C.; Schwarz, S.; Moser, G.; Cvitic, S.; Jantscher-Krenn, E.; Gauster, M.; Hiden, U. Placental endocrine activity: Adaptation and disruption of maternal glucose metabolism in pregnancy and the influence of fetal sex. Int. J. Mol. Sci. 2021, 22, 12722. [Google Scholar] [CrossRef]
- Simpson, S.; Smith, L.; Bowe, J. Placental peptides regulating islet adaptation to pregnancy: Clinical potential in gestational diabetes mellitus. Curr. Opin. Pharmacol. 2018, 43, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Burton, G.J.; Jauniaux, E.; Charnock-Jones, D.S. The influence of the intrauterine environment on human placental development. Int. J. Dev. Biol. 2010, 54, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimasuay, K.G.; Boeuf, P.; Powell, T.L.; Jansson, T. Placental responses to changes in the maternal environment determine fetal growth. Front. Physiol. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, L.; Perez-Garcia, V.; Hemberger, M.; Brett, K.E.; Ferraro, Z.M.; Yockell-Lelievre, J.; Gruslin, A.; Adamo, K.B. Maternal–Fetal nutrient transport in pregnancy pathologies: The role of the placenta. Int. J. Mol. Sci. 2014, 15, 16153–16185. [Google Scholar]
- Hemberger, M.; Hanna, C.W.; Dean, W. Mechanisms of early placental development in mouse and humans. Nat. Rev. Genet. 2020, 21, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Morales-Prieto, D.M.; Pastuschek, J.; Fröhlich, K.; Markert, U.R. Only humans have human placentas: Molecular differences between mice and humans. J. Reprod. Immunol. 2015, 108, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Cox, B.; Kotlyar, M.; Evangelou, A.I.; Ignatchenko, V.; Ignatchenko, A.; Whiteley, K.; Jurisica, I.; Adamson, S.L.; Rossant, J.; Kislinger, T. Comparative systems biology of human and mouse as a tool to guide the modeling of human placental pathology. Mol. Syst. Biol. 2009, 5, 1–15. [Google Scholar] [CrossRef]
- Coan, P.M.; Vaughan, O.R.; Sekita, Y.; Finn, S.L.; Burton, G.J.; Constancia, M.; Fowden, A.L. Adaptations in placental phenotype support fetal growth during undernutrition of pregnant mice. J. Physiol. 2010, 3, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; McKnight, R.A.; Raychaudhuri, S.; Shin, B.C.; Ma, Z.; Moley, K.; Devaskar, S.U. Glucose transporter isoform-3 mutations cause early pregnancy loss and fetal growth restriction. Am. J. Physiol.—Endocrinol. Metab. 2007, 292, E1241–E1255. [Google Scholar] [CrossRef] [Green Version]
- Constância, M.; Angiolini, E.; Sandovici, I.; Smith, P.; Smith, R.; Kelsey, G.; Dean, W.; Ferguson-smith, A.; Sibley, C.P.; Reik, W.; et al. Adaptation of nutrient supply to fetal demand in the mouse involves interaction between the Igf2 gene and placental transporter systems. Proc. Natl. Acad. Sci. USA 2005, 102, 19219–19224. [Google Scholar] [CrossRef] [Green Version]
- Wyrwoll, C.S.; Seckl, J.R.; Holmes, M.C. Altered placental function of 11β-hydroxysteroid dehydrogenase 2 knockout mice. Endocrinology 2009, 150, 1287–1293. [Google Scholar] [CrossRef] [Green Version]
- López-Tello, J.; Pérez-García, V.; Khaira, J.; Kusinski, L.C.; Cooper, W.N.; Andreani, A.; Grant, I.; de Liger, E.F.; Lam, B.Y.H.; Hemberger, M.; et al. Fetal and trophoblast PI3K p110α have distinct roles in regulating resource supply to the growing fetus in mice. Elife 2019, 8, 1–25. [Google Scholar] [CrossRef] [PubMed]
- James-Allan, L.B.; Teal, S.; Powell, T.L.; Jansson, T. Changes in Placental Nutrient Transporter Protein Expression and Activity Across Gestation in Normal and Obese Women. Reprod. Sci. 2020, 27, 1758–1769. [Google Scholar] [CrossRef]
- Borges, M.H.; Pullockaran, J.; Catalano, P.M.; Baumann, M.U.; Zamudio, S.; Illsley, N.P. Human placental GLUT1 glucose transporter expression and the fetal insulin-like growth factor axis in pregnancies complicated by diabetes. Biochim. Biophys. Acta—Mol. Basis Dis. 2019, 1865, 2411–2419. [Google Scholar] [CrossRef] [PubMed]
- Sferruzzi-Perri, A.N.; López-Tello, J.; Fowden, A.L.; Constancia, M. Maternal and fetal genomes interplay through phosphoinositol 3-kinase (PI3K)-p110α signaling to modify placental resource allocation. Proc. Natl. Acad. Sci. USA 2016, 113, 11255–11260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.; Sferruzzi-Perri, A.N. Placental mitochondrial function in response to gestational exposures. Placenta 2021, 104, 124–137. [Google Scholar] [CrossRef]
- Aye, I.L.M.H.; Aiken, C.E.; Charnock-Jones, D.S.; Smith, G.C.S. Placental energy metabolism in health and disease—Significance of development and implications for preeclampsia. Am. J. Obstet. Gynecol. 2022, 226, S928–S944. [Google Scholar] [CrossRef]
- Martinez, F.; Olvera-Sanchez, S.; Esparza-Perusquia, M.; Gomez-Chang, E.; Flores-Herrera, O.; Jornayvaz, F.R.; Shulman, G.I.; Katic, M.; Kennedy, A.R.; Leykin, I.; et al. Multiple functions of syncytiotrophoblast mitochondria. Essays Biochem. 2010, 103, 69–84. [Google Scholar] [CrossRef]
- Waker, C.A.; Albers, R.E.; Pye, R.L.; Doliboa, S.R.; Wyatt, C.N.; Brown, T.L.; Mayes, D.A. AMPK Knockdown in Placental Labyrinthine Progenitor Cells Results in Restriction of Critical Energy Resources and Terminal Differentiation Failure. Stem Cells Dev. 2017, 26, 808–817. [Google Scholar] [CrossRef]
- Bartho, L.A.; Fisher, J.J.; Walton, S.L.; Perkins, A.V.; Cuffe, J.S.M. The effect of gestational age on mitochondrial properties of the mouse placenta. Reprod. Fertil. 2022, 3, 19–29. [Google Scholar] [CrossRef]
- Murray, A.J. Oxygen delivery and fetal-placental growth: Beyond a question of supply and demand? Placenta 2012, 33, e16–e22. [Google Scholar] [CrossRef]
- Sferruzzi-Perri, A.N.; Higgins, J.S.; Vaughan, O.R.; Murray, A.J.; Fowden, A.L. Placental mitochondria adapt developmentally and in response to hypoxia to support fetal growth. Proc. Natl. Acad. Sci. USA 2019, 116, 1621–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, O.J.; Hickey, A.J.R.; Alvsaker, A.; Moran, S.; Hedges, C.; Chamley, L.W.; Perkins, A.V. Changes in mitochondrial respiration in the human placenta over gestation. Placenta 2017, 57, 102–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebelato, H.J.; Esquisatto, M.A.M.; Moraes, C.; Amaral, M.E.C.; Catisti, R. Gestational protein restriction induces alterations in placental morphology and mitochondrial function in rats during late pregnancy. J. Mol. Histol. 2013, 44, 629–637. [Google Scholar] [CrossRef]
- Sobrevia, L.; Valero, P.; Grismaldo, A.; Villalobos-Labra, R.; Pardo, F.; Subiabre, M.; Armstrong, G.; Toledo, F.; Vega, S.; Cornejo, M.; et al. Mitochondrial dysfunction in the fetoplacental unit in gestational diabetes mellitus. Biochim. Biophys. Acta—Mol. Basis Dis. 2020, 1866, 165948. [Google Scholar] [CrossRef]
- Mayeur, S.; Lancel, S.; Theys, N.; Lukaszewski, M.-A.; Duban-Deweer, S.; Bastide, B.; Hachani, J.; Cecchelli, R.; Breton, C.; Gabory, A.; et al. Maternal calorie restriction modulates placental mitochondrial biogenesis and bioenergetic efficiency: Putative involvement in fetoplacental growth defects in rats. Am. J. Physiol. Endocrinol. Metab. 2013, 304, 14–22. [Google Scholar] [CrossRef]
- Yung, H.W.; Colleoni, F.; Dommett, E.; Cindrova-Davies, T.; Kingdom, J.; Murray, A.J.; Burton, G.J. Noncanonical mitochondrial unfolded protein response impairs placental oxidative phosphorylation in early-onset preeclampsia. Proc. Natl. Acad. Sci. USA 2019, 116, 18109–18118. [Google Scholar] [CrossRef] [Green Version]
- Mandò, C.; De Palma, C.; Stampalija, T.; Anelli, G.M.; Figus, M.; Novielli, C.; Parisi, F.; Clementi, E.; Ferrazzi, E.; Cetin, I. Placental mitochondrial content and function in intrauterine growth restriction and preeclampsia. Am. J. Physiol.—Endocrinol. Metab. 2014, 306, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Holland, O.J.; Cuffe, J.S.M.; Dekker Nitert, M.; Callaway, L.; Kwan Cheung, K.A.; Radenkovic, F.; Perkins, A.V. Placental mitochondrial adaptation in preeclampsia associated with progression to term delivery. Cell Death Dis. 2018, 9, 1150. [Google Scholar] [CrossRef] [Green Version]
- Abbade, J.; Klemetti, M.M.; Farrell, A.; Ermini, L.; Gillmore, T.; Sallais, J.; Tagliaferro, A.; Post, M.; Caniggia, I. Increased placental mitochondrial fusion in gestational diabetes mellitus: An adaptive mechanism to optimize feto-placental metabolic homeostasis? BMJ Open Diabetes Res. Care 2020, 8, e000923. [Google Scholar] [CrossRef]
- Napso, T.; Hung, Y.P.; Davidge, S.T.; Care, A.S.; Sferruzzi-Perri, A.N. Advanced maternal age compromises fetal growth and induces sex-specific changes in placental phenotype in rats. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Napso, T.; Lean, S.C.; Lu, M.; Mort, E.J.; Desforges, M.; Moghimi, A.; Bartels, B.; El-Bacha, T.; Fowden, A.L.; Camm, E.J.; et al. Diet-induced maternal obesity impacts feto-placental growth and induces sex-specific alterations in placental morphology, mitochondrial bioenergetics, dynamics, lipid metabolism and oxidative stress in mice. Acta Physiol. 2022, 234, e13795. [Google Scholar] [CrossRef] [PubMed]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat. Rev. Genet. 2006, 7, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Bettedi, L.; Yan, A.; Schuster, E.; Alic, N.; Foukas, L.C. Increased mitochondrial and lipid metabolism is a conserved effect of Insulin/PI3K pathway downregulation in adipose tissue. Sci. Rep. 2020, 10, 3418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, M.; Wang, J.; Wang, W.; Liu, J.; Wong, C.W. Phosphatidylinositol 3-kinase affects mitochondrial function in part through inducing peroxisome proliferator-activated receptor γ coactivator-1β expression. Br. J. Pharmacol. 2011, 162, 1000–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.S.L.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/ mTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu-Sekine, B.; Kim, S.F. The Inositol Phosphate System—A Coordinator of Metabolic Adaptability. Int. J. Mol. Sci. 2022, 23, 6747. [Google Scholar] [CrossRef]
- Cardoso, S.; López, I.P.; Piñeiro-Hermida, S.; Pichel, J.G.; Moreira, P.I. Igf1r deficiency modulates brain signaling pathways and disturbs mitochondria and redox homeostasis. Biomedicines 2021, 9, 158. [Google Scholar] [CrossRef]
- Zhang, D.; Wei, Y.; Huang, Q.; Chen, Y.; Zeng, K.; Yang, W.; Chen, J.; Chen, J. Important Hormones Regulating Lipid Metabolism. Molecules 2022, 27, 7052. [Google Scholar] [CrossRef]
- Foukas, L.C.; Claret, M.; Pearce, W.; Okkenhaug, K.; Meek, S.; Peskett, E.; Sancho, S.; Smith, A.J.H.; Withers, D.J.; Vanhaesebroeck, B. Critical role for the p110 a phosphoinositide-3-OH kinase in growth and metabolic regulation. Nature 2006, 441, 3–7. [Google Scholar] [CrossRef]
- Knight, Z.A.; Gonzalez, B.; Feldman, M.E.; Zunder, E.R.; Goldenberg, D.D.; Williams, O.; Loewith, R.; Stokoe, D.; Balla, A.; Toth, B.; et al. A Pharmacological Map of the PI3-K Family Defines a Role for p110 a in Insulin Signaling. Cell 2006, 125, 733–747. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, C.M.; Kondo, T.; Sajan, M.; Luo, J.; Bronson, R.; Asano, T.; Farese, R.; Cantley, L.C.; Kahn, C.R. Divergent regulation of hepatic glucose and lipid metabolism by phosphoinositide 3-kinase via Akt and PKC l/z. Cell Metab. 2006, 3, 343–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sopasakis, V.R.; Liu, P.; Suzuki, R.; Kondo, T.; Winnay, J.; Tran, T.T.; Asano, T.; Smyth, G.; Sajan, M.P.; Farese, R.V.; et al. Short Article Specific Roles of the p110 a Isoform of Phosphatidylinsositol 3-Kinase in Hepatic Insulin Signaling and Metabolic Regulation. Cell Metab. 2010, 11, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-García, V.; Redondo-Muñoz, J.; Kumar, A.; Carrera, A.C. Cell activation-induced phosphoinositide 3-kinase alpha/beta dimerization regulates PTEN activity. Mol. Cell Biol. 2014, 34, 3359–3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Tello, J.; Salazar-Petres, E.; Webb, L.; Fowden, A.L.; Sferruzzi-Perri, A.N. Ablation of PI3K-p110alpha Impairs Maternal Metabolic Adaptations to Pregnancy. Front. Cell Dev. Biol. 2022, 10, 928210. [Google Scholar] [CrossRef]
- Makarova, E.N.; Kochubei, E.D.; Bazhan, N.M. Regulation of food consumption during pregnancy and lactation in mice. Neurosci. Behav. Physiol. 2010, 40, 263–267. [Google Scholar] [CrossRef]
- Pereira-Carvalho, D.; Salazar-Petres, E.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. Maternal and Fetal PI3K-p110α Deficiency Induces Sex-Specific Changes in Conceptus Growth and Placental Mitochondrial Bioenergetic Reserve in Mice. Vet. Sci. 2022, 9, 501. [Google Scholar] [CrossRef]
- Djafarzadeh, S.; Jakob, S.M. High-resolution respirometry to assess mitochondrial function in permeabilized and intact cells. J. Vis. Exp. 2017, 2017, 1–11. [Google Scholar]
- Romero-Calvo, I.; Ocón, B.; Martínez-Moya, P.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; de Medina, F.S. Reversible Ponceau staining as a loading control alternative to actin in Western blots. Anal. Biochem. 2010, 401, 318–320. [Google Scholar] [CrossRef]
- Tunster, S.J.; Creeth, H.D.J.; John, R.M. The imprinted Phlda2 gene modulates a major endocrine compartment of the placenta to regulate placental demands for maternal resources. Dev. Biol. 2016, 409, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Kalisch-Smith, J.I.; Simmons, D.G.; Dickinson, H.; Moritz, K.M. Review: Sexual dimorphism in the formation, function and adaptation of the placenta. Placenta 2017, 54, 10–16. [Google Scholar] [CrossRef]
- Eriksson, J.G.; Kajantie, E.; Osmond, C.; Thornburg, K.; Barker, D.J.P. Boys live dangerously in the womb. Am. J. Hum. Biol. Counc. 2010, 22, 330–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartels, H.C.; Geraghty, A.A.; O’Brien, E.C.; Kranidi, A.; Mehegan, J.; Yelverton, C.; McDonnell, C.M.; McAuliffe, F.M. Fetal Growth Trajectories and Their Association with Maternal, Cord Blood, and 5-year Child Adipokines. J. Nutr. Metab. 2020, 2020, 4861523. [Google Scholar] [CrossRef] [PubMed]
- Broere-Brown, Z.A.; Baan, E.; Schalekamp-Timmermans, S.; Verburg, B.O.; Jaddoe, V.W.V.; Steegers, E.A.P. Sex-specific differences in fetal and infant growth patterns: A prospective population-based cohort study. Biol. Sex Differ. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aykroyd, B.R.L.; Tunster, S.J.; Sferruzzi-Perri, A.N. Loss of imprinting of the Igf2-H19 ICR1 enhances placental endocrine capacity via sex-specific alterations in signalling pathways in the mouse. Development 2022, 149, dev199811. [Google Scholar] [CrossRef] [PubMed]
- Aykroyd, B.R.L.; Tunster, S.J.; Sferruzzi-Perri, A.N. Igf2 deletion alters mouse placenta endocrine capacity in a sexually dimorphic manner. J. Endocrinol. 2020, 246, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Sferruzzi-Perri, A.N.; Sandovici, I.; Constancia, M.; Fowden, A.L. Placental phenotype and the insulin-like growth factors: Resource allocation to fetal growth. J. Physiol. 2017, 15, 5057–5093. [Google Scholar] [CrossRef] [Green Version]
- Wen, Q.; Cheng, C.Y.; Liu, Y.X. Development, function and fate of fetal Leydig cells. Semin. Cell Dev. Biol. 2016, 59, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Biason-Lauber, A.; Chaboissier, M.C. Ovarian development and disease: The known and the unexpected. Semin. Cell Dev. Biol. 2015, 45, 59–67. [Google Scholar] [CrossRef]
- Hebert, J.F.; Myatt, L. Placental mitochondrial dysfunction with metabolic diseases: Therapeutic approaches. Biochim. Biophys. Acta—Mol. Basis Dis. 2021, 1867, 165967. [Google Scholar] [CrossRef]
- Muralimanoharan, S.; Guo, C.; Myatt, L.; Maloyan, A. Sexual dimorphism in MIR-210 expression and mitochondrial dysfunction in the placenta with maternal obesity. Int. J. Obes. 2015, 39, 1274–1281. [Google Scholar] [CrossRef] [Green Version]
- Mishra, J.S.; Blesson, C.S.; Kumar, S. Testosterone decreases placental mitochondrial content and cellular bioenergetics. Biology 2020, 9, 176. [Google Scholar] [CrossRef] [PubMed]
- Cuffe, J.S.M.; Dickinson, H.; Simmons, D.G.; Moritz, K.M. Sex specific changes in placental growth and MAPK following short term maternal dexamethasone exposure in the mouse. Placenta 2011, 32, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.S.; Myatt, L. Sexual dimorphism in the effect of maternal obesity on antioxidant defense mechanisms in the human placenta. Placenta 2017, 51, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Teague, A.M.; Tryggestad, J.B.; Aston, C.E.; Lyons, T.; Chernausek, S.D. Effects of maternal diabetes and fetal sex on human placenta mitochondrial biogenesis. Placenta 2017, 57, 26–32. [Google Scholar] [CrossRef]
- Ganguly, E.; Kirschenman, R.; Spaans, F.; Holody, C.D.; Phillips, T.E.J.; Case, C.P.; Murphy, M.P.; Lemieux, H.; Davidge, S.T. Nanoparticle-encapsulated antioxidant improves placental mitochondrial function in a sexually dimorphic manner in a rat model of prenatal hypoxia. FASEB J. 2021, 35, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Vangrieken, P.; Al-Nasiry, S.; Bast, A.; Leermakers, P.A.; Tulen, C.B.M.; Schiffers, P.M.H.; van Schooten, F.J.; Remels, A.H.V. Placental Mitochondrial Abnormalities in Preeclampsia. Reprod. Sci. 2021, 28, 2186–2199. [Google Scholar] [CrossRef]
- Fisher, J.J.; Bartho, L.A.; Perkins, A.V.; Holland, O.J. Placental mitochondria and reactive oxygen species in the physiology and pathophysiology of pregnancy. Clin. Exp. Pharmacol. Physiol. 2020, 47, 176–184. [Google Scholar] [CrossRef]
- Hastie, R.; Lappas, M. The effect of pre-existing maternal obesity and diabetes on placental mitochondrial content and electron transport chain activity. Placenta 2014, 35, 673–683. [Google Scholar] [CrossRef]
- Muralimanoharan, S.; Maloyan, A.; Myatt, L. Mitochondrial function and glucose metabolism in the placenta with gestational diabetes mellitus: Role of miR-143. Clin. Sci. 2016, 130, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Luo, W.; Li, S.; Zhao, S.; Sho, T.; Xu, X.; Zhang, J.; Xu, W.; Xu, J. Reactive oxygen species mediated placental oxidative stress, mitochondrial content, and cell cycle progression through mitogen-activated protein kinases in intrauterine growth restricted pigs. Reprod. Biol. 2018, 18, 422–431. [Google Scholar] [CrossRef]
- Ramírez-Emiliano, J.; Fajardo-Araujo, M.E.; Zúñiga-Trujillo, I.; Pérez-Vázquez, V.; Sandoval-Salazar, C.; Órnelas-Vázquez, J.K. Mitochondrial content, oxidative, and nitrosative stress in human full-term placentas with gestational diabetes mellitus. Reprod. Biol. Endocrinol. 2017, 15, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, R.; Chiarello, D.I.; Abad, C.; Rojas, D.; Toledo, F.; Sobrevia, L. Oxidative stress and mitochondrial dysfunction in early-onset and late-onset preeclampsia. Biochim. Biophys. Acta—Mol. Basis Dis. 2020, 1866, 165961. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Expert Reviews Pathophysiology of placental-derived fetal growth restriction. Am. J. Obstet. Gynecol. 2018, 218, S745–S761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Tello, J.; Sferruzzi-Perri, A.N. Characterization of placental endocrine function and fetal brain development in a mouse model of small for gestational age. Front. Endocrinol. 2023, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar-Petres, E.; Pereira-Carvalho, D.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. Maternal and Intrauterine Influences on Feto-Placental Growth Are Accompanied by Sexually Dimorphic Changes in Placental Mitochondrial Respiration, and Metabolic Signalling Pathways. Cells 2023, 12, 797. https://doi.org/10.3390/cells12050797
Salazar-Petres E, Pereira-Carvalho D, Lopez-Tello J, Sferruzzi-Perri AN. Maternal and Intrauterine Influences on Feto-Placental Growth Are Accompanied by Sexually Dimorphic Changes in Placental Mitochondrial Respiration, and Metabolic Signalling Pathways. Cells. 2023; 12(5):797. https://doi.org/10.3390/cells12050797
Chicago/Turabian StyleSalazar-Petres, Esteban, Daniela Pereira-Carvalho, Jorge Lopez-Tello, and Amanda N. Sferruzzi-Perri. 2023. "Maternal and Intrauterine Influences on Feto-Placental Growth Are Accompanied by Sexually Dimorphic Changes in Placental Mitochondrial Respiration, and Metabolic Signalling Pathways" Cells 12, no. 5: 797. https://doi.org/10.3390/cells12050797
APA StyleSalazar-Petres, E., Pereira-Carvalho, D., Lopez-Tello, J., & Sferruzzi-Perri, A. N. (2023). Maternal and Intrauterine Influences on Feto-Placental Growth Are Accompanied by Sexually Dimorphic Changes in Placental Mitochondrial Respiration, and Metabolic Signalling Pathways. Cells, 12(5), 797. https://doi.org/10.3390/cells12050797