




Molecular Regulation of Autophagy and Asymmetric Cell Division by Cancer Stem Cell Marker CD133


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Cancer Stem Cell Marker CD133
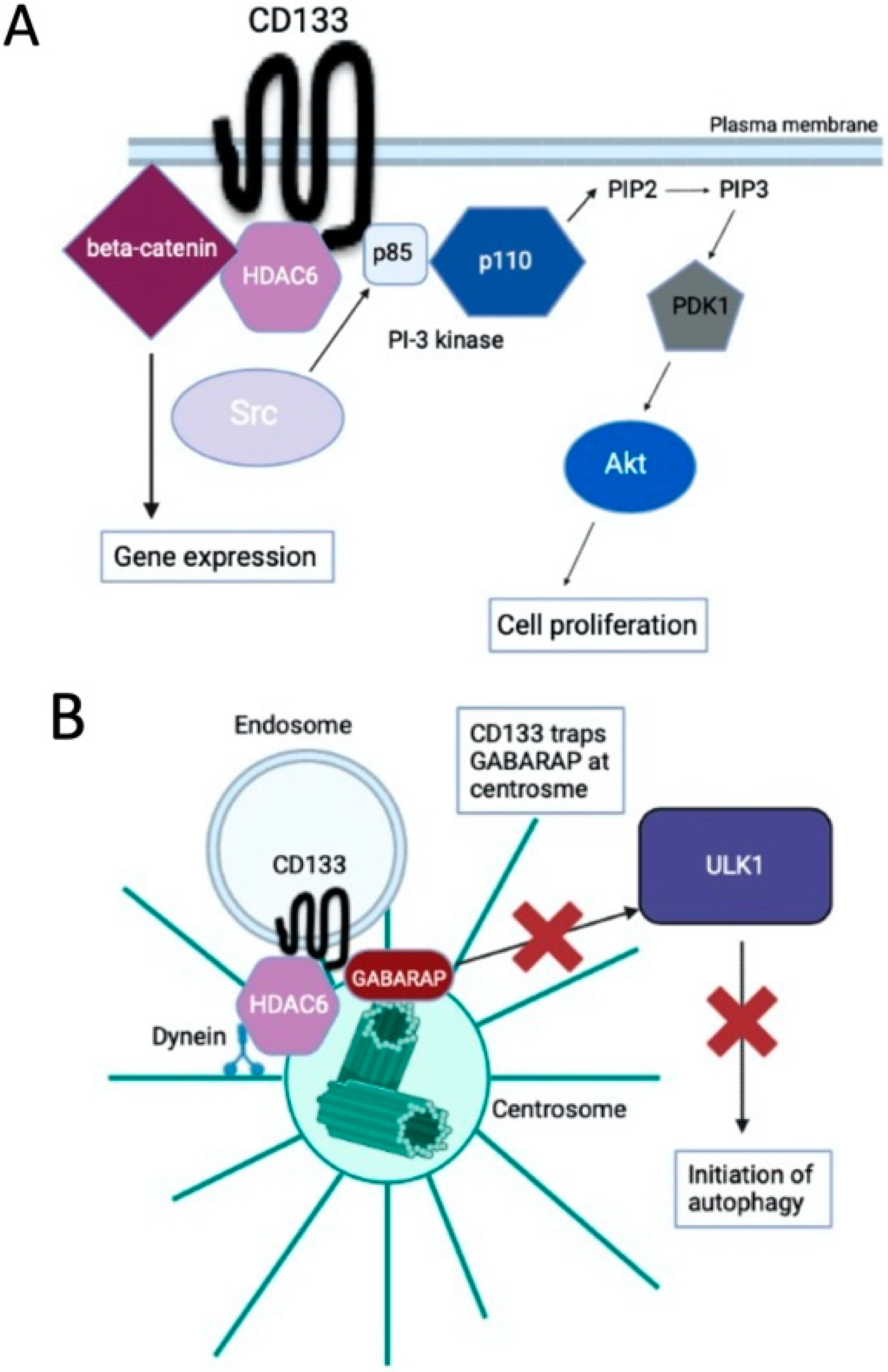
3. CD133 Localizes to Centrosomes and Suppresses Autophagy Activation
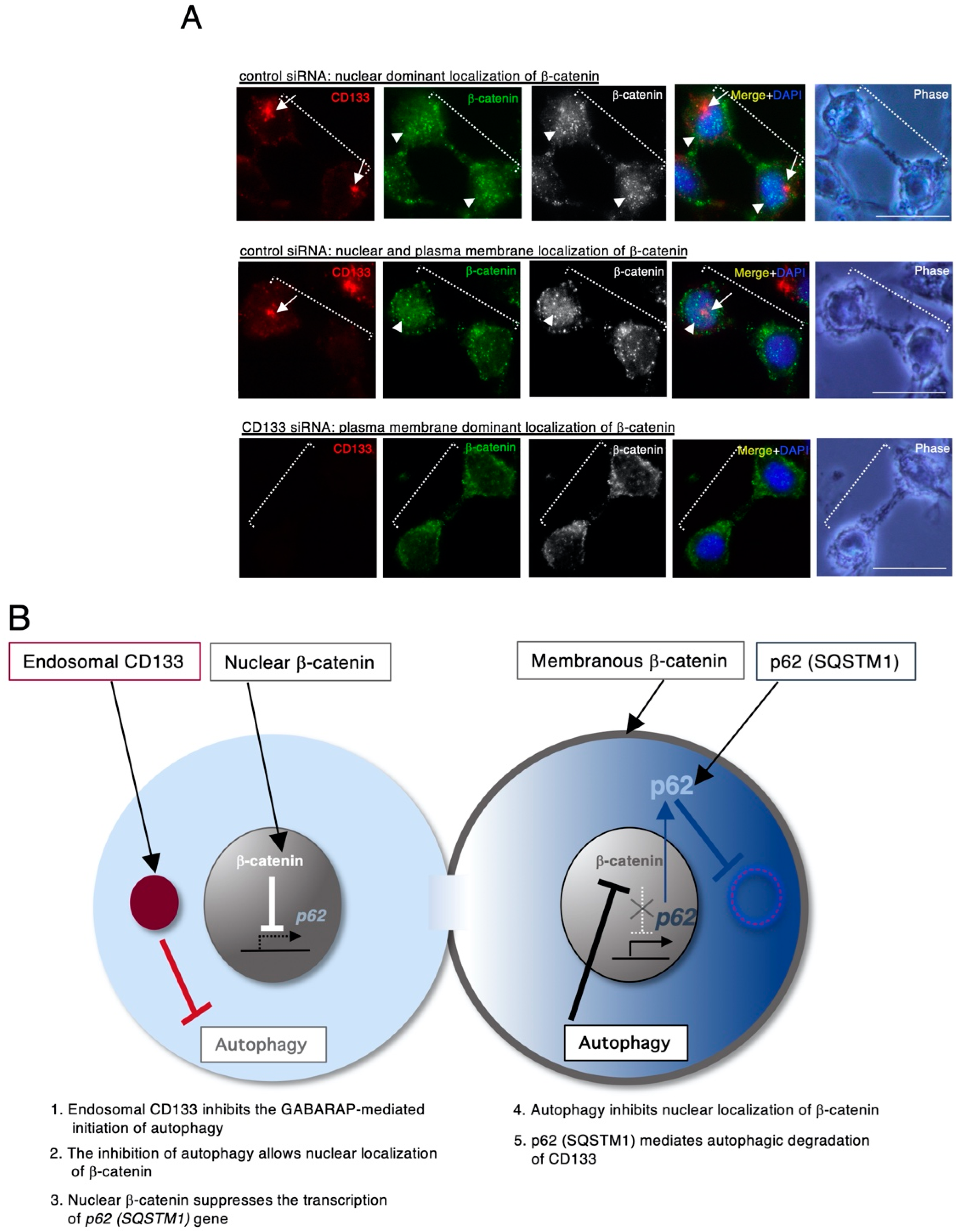
4. CD133 Heterogeneously Localizes to Centrosomes and Generates Asymmetric Cell Division, Resulting in Daughter Cells Exhibiting a Variety of Autophagic Activities
5. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clevers, H. The cancer stem cell: Premises, promises and challenges. Nat. Med. 2011, 17, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Grosse-Gehling, P.; Fargeas, C.A.; Dittfeld, C.; Garbe, Y.; Alison, M.R.; Corbeil, D.; Kunz-Schughart, L.A. CD133 as a biomarker for putative cancer stem cells in solid tumours: Limitations, problems and challenges. J. Pathol. 2012, 229, 355–378. [Google Scholar] [CrossRef]
- Li, Z. CD133: A stem cell biomarker and beyond. Exp. Hematol. Oncol. 2013, 2, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, H.; Li, Y.; Shibaki, M.; Mori, D.; Yasunami, M.; Sato, S.; Matsunaga, H.; Mae, T.; Kodama, K.; Kamijo, T.; et al. Recycling endosomal CD133 functions as an inhibitor of autophagy at the pericentrosomal region. Sci. Rep. 2019, 9, 2236. [Google Scholar] [CrossRef] [Green Version]
- Izumi, H.; Li, Y.; Yasunami, M.; Sato, S.; Mae, T.; Kaneko, Y.; Nakagawara, A. Asymmetric Pericentrosomal CD133 Endosomes Induce the Unequal Autophagic Activity during Cytokinesis in CD133-Positive Human Neuroblastoma Cells. Stem Cells 2022, 40, 371–384. [Google Scholar] [CrossRef]
- Miraglia, S.; Godfrey, W.; Yin, A.H.; Atkins, K.; Warnke, R.; Holden, J.T.; Bray, R.A.; Waller, E.K.; Buck, D.W. A novel five-transmembrane hematopoietic stem cell antigen: Isolation, characterization, and molecular cloning. Blood 1997, 90, 5013–5021. [Google Scholar] [CrossRef] [PubMed]
- Weigmann, A.; Corbeil, D.; Hellwig, A.; Huttner, W.B. Prominin, a novel microvilli-specific polytopic membrane protein of the apical surface of epithelial cells, is targeted to plasmalemmal protrusions of non-epithelial cells. Proc. Natl. Acad. Sci. USA 1997, 94, 12425–12430. [Google Scholar] [CrossRef] [Green Version]
- Yin, A.H.; Miraglia, S.; Zanjani, E.D.; Almeida-Porada, G.; Ogawa, M.; Leary, A.G.; Olweus, J.; Kearney, J.; Buck, D.W. AC133, a novel marker for human hematopoietic stem and progenitor cells. Blood 1997, 90, 5002–5012. [Google Scholar] [CrossRef] [Green Version]
- Boivin, D.; Labbé, D.; Fontaine, N.; Lamy, S.; Beaulieu, E.; Gingras, D.; Béliveau, R. The Stem Cell Marker CD133 (Prominin-1) is Phosphorylated on Cytoplasmic Tyrosine-828 and Tyrosine-852 by Src and Fyn Tyrosine Kinases. Biochemistry 2009, 48, 3998–4007. [Google Scholar] [CrossRef]
- Wei, Y.; Jiang, Y.; Zou, F.; Liu, Y.; Wang, S.; Xu, N.; Xu, W.; Cui, C.; Xing, Y.; Liu, Y.; et al. Activation of PI3K/Akt pathway by CD133-p85 interaction promotes tumorigenic capacity of glioma stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, 6829–6834. [Google Scholar] [CrossRef] [Green Version]
- Mak, A.B.; Nixon, A.M.; Kittanakom, S.; Stewart, J.M.; Chen, G.I.; Curak, J.; Gingras, A.-C.; Mazitschek, R.; Neel, B.G.; Stagljar, I.; et al. Regulation of CD133 by HDAC6 Promotes β-Catenin Signaling to Suppress Cancer Cell Differentiation. Cell Rep. 2012, 2, 951–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, H.; Kaneko, Y.; Nakagawara, A. Regulation of autophagy and asymmetric cell division by the stem cell marker CD133. SEIKAGAKU 2022, 94, 415–418. (In Japanese) [Google Scholar] [CrossRef]
- Nigg, E.A.; Holland, A.J. Once and only once: Mechanisms of centriole duplication and their deregulation in disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 297–312. [Google Scholar] [CrossRef]
- Joachim, J.; Jefferies, H.B.; Razi, M.; Frith, D.; Snijders, A.P.; Chakravarty, P.; Judith, D.; Tooze, S.A. Activation of ULK Kinase and Autophagy by GABARAP Trafficking from the Centrosome Is Regulated by WAC and GM130. Mol. Cell 2015, 60, 899–913. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in Mammalian Autophagy Research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, C.A.; Dooley, H.C.; Tooze, S.A. Endocytosis and autophagy: Shared machinery for degradation. BioEssays 2012, 35, 34–45. [Google Scholar] [CrossRef]
- Tooze, S.A.; Abada, A.; Elazar, Z. Endocytosis and Autophagy: Exploitation or Cooperation? Cold Spring Harb. Perspect. Biol. 2014, 6, a018358. [Google Scholar] [CrossRef]
- Lamb, C.A.; Yoshimori, T.; Tooze, S.A. The autophagosome: Origins unknown, biogenesis complex. Nat. Rev. Mol. Cell Biol. 2013, 14, 759–774. [Google Scholar] [CrossRef]
- Rogov, V.; Dötsch, V.; Johansen, T.; Kirkin, V. Interactions between Autophagy Receptors and Ubiquitin-Like Proteins Form the Molecular Basis for Selective Autophagy. Mol. Cell 2014, 53, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Levine, B. Autophagy in mammalian development and differentiation. Nat. Cell Biol. 2010, 12, 823–830. [Google Scholar] [CrossRef]
- Pan, H.; Cai, N.; Li, M.; Liu, G.-H.; Belmonte, J.C.I. Autophagic control of cell ‘stemness’. EMBO Mol. Med. 2013, 5, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Petherick, K.J.; Williams, A.C.; Lane, J.D.; Ordóñez-Morán, P.; Huelsken, J.; Collard, T.J.; Smartt, H.J.; Batson, J.; Malik, K.; Paraskeva, C.; et al. Autolysosomal β-catenin degradation regulates Wnt-autophagy-p62 crosstalk. EMBO J. 2013, 32, 1903–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, E.; Skop, A.R. Endosomal recycling regulation during cytokinesis. Commun. Integr. Biol. 2009, 2, 444–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, H.; Kaneko, Y. Evidence of asymmetric cell division and centrosome inheritance in human neuroblastoma cells. Proc. Natl. Acad. Sci. USA 2012, 109, 18048–18053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Yamashita, Y.M. Centrosome-centric view of asymmetric stem cell division. Open Biol. 2021, 11, 200314. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Izumi, H.; Kaneko, Y.; Nakagawara, A. Molecular Regulation of Autophagy and Asymmetric Cell Division by Cancer Stem Cell Marker CD133. Cells 2023, 12, 819. https://doi.org/10.3390/cells12050819
Izumi H, Kaneko Y, Nakagawara A. Molecular Regulation of Autophagy and Asymmetric Cell Division by Cancer Stem Cell Marker CD133. Cells. 2023; 12(5):819. https://doi.org/10.3390/cells12050819
Chicago/Turabian StyleIzumi, Hideki, Yasuhiko Kaneko, and Akira Nakagawara. 2023. "Molecular Regulation of Autophagy and Asymmetric Cell Division by Cancer Stem Cell Marker CD133" Cells 12, no. 5: 819. https://doi.org/10.3390/cells12050819
APA StyleIzumi, H., Kaneko, Y., & Nakagawara, A. (2023). Molecular Regulation of Autophagy and Asymmetric Cell Division by Cancer Stem Cell Marker CD133. Cells, 12(5), 819. https://doi.org/10.3390/cells12050819