
Isolation and Characterization of Novel Canine Osteosarcoma Cell Lines from Chemotherapy-Naïve Patients

 , and
, and
Abstract
:1. Introduction
2. Materials and Method
2.1. Clinical Material, Cell Isolation and Cultivation
2.2. Cell Line Authentication (STR Analysis)
2.3. Growth Kinetic Analysis
2.4. Soft Agar Colony Formation Assay
2.5. Wound Closure Assay (Scratch Assay)
2.6. Cell Culture Insert Migration and Invasion Assay
2.7. Spheroid Formation Assay
2.8. Histochemical and Immunohistochemical Analyses
2.9. Reverse Transcription Quantitative PCR Analysis
2.10. Statistical Analysis
3. Results
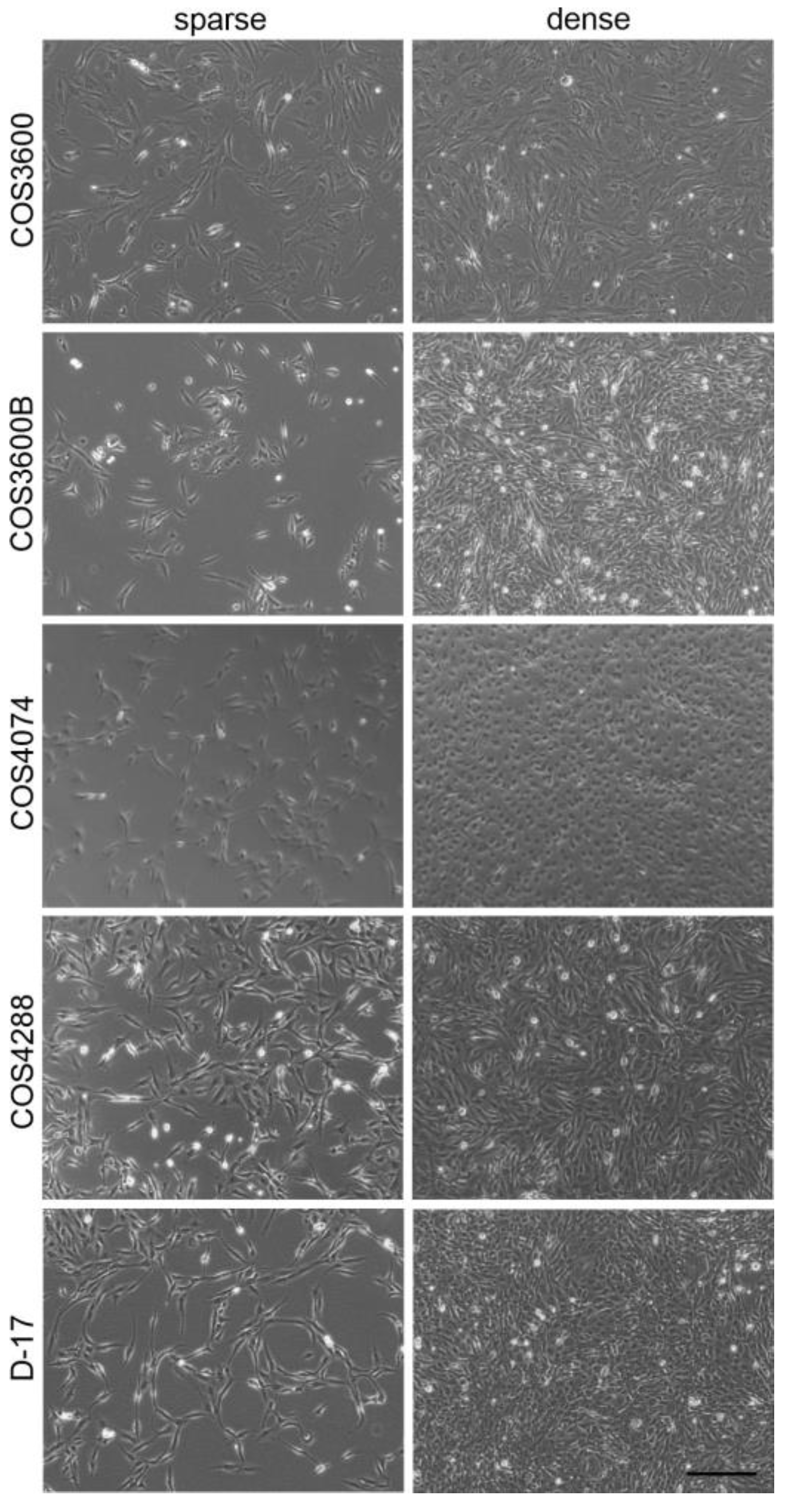
3.1. Cell Isolation and Morphology
3.2. Authentication of the Cell Lines
3.3. Growth Kinetic Analysis
3.4. Anchorage-Independent Growth in Soft Agar
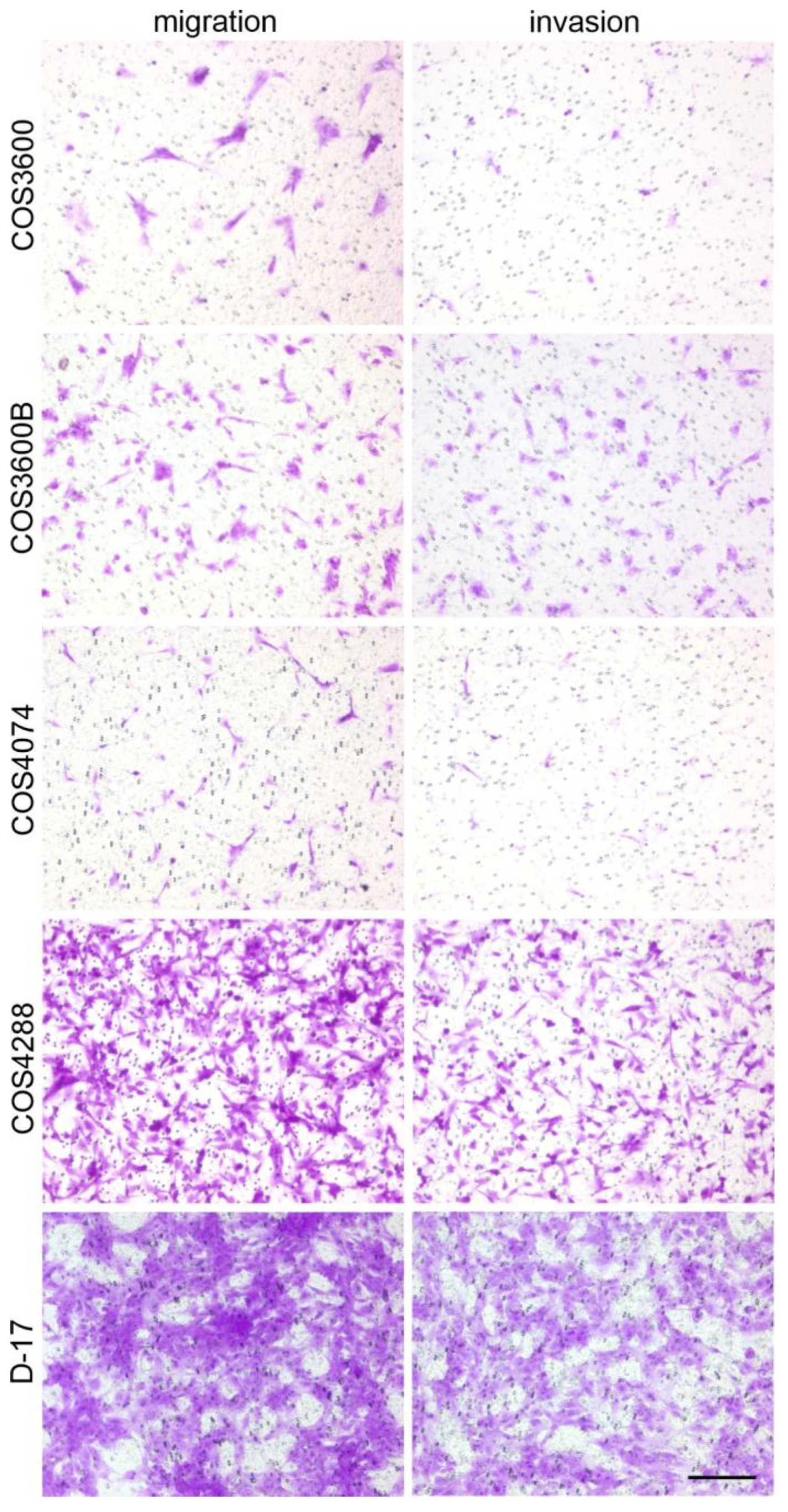
3.5. Cell Migration and Invasion Analyses
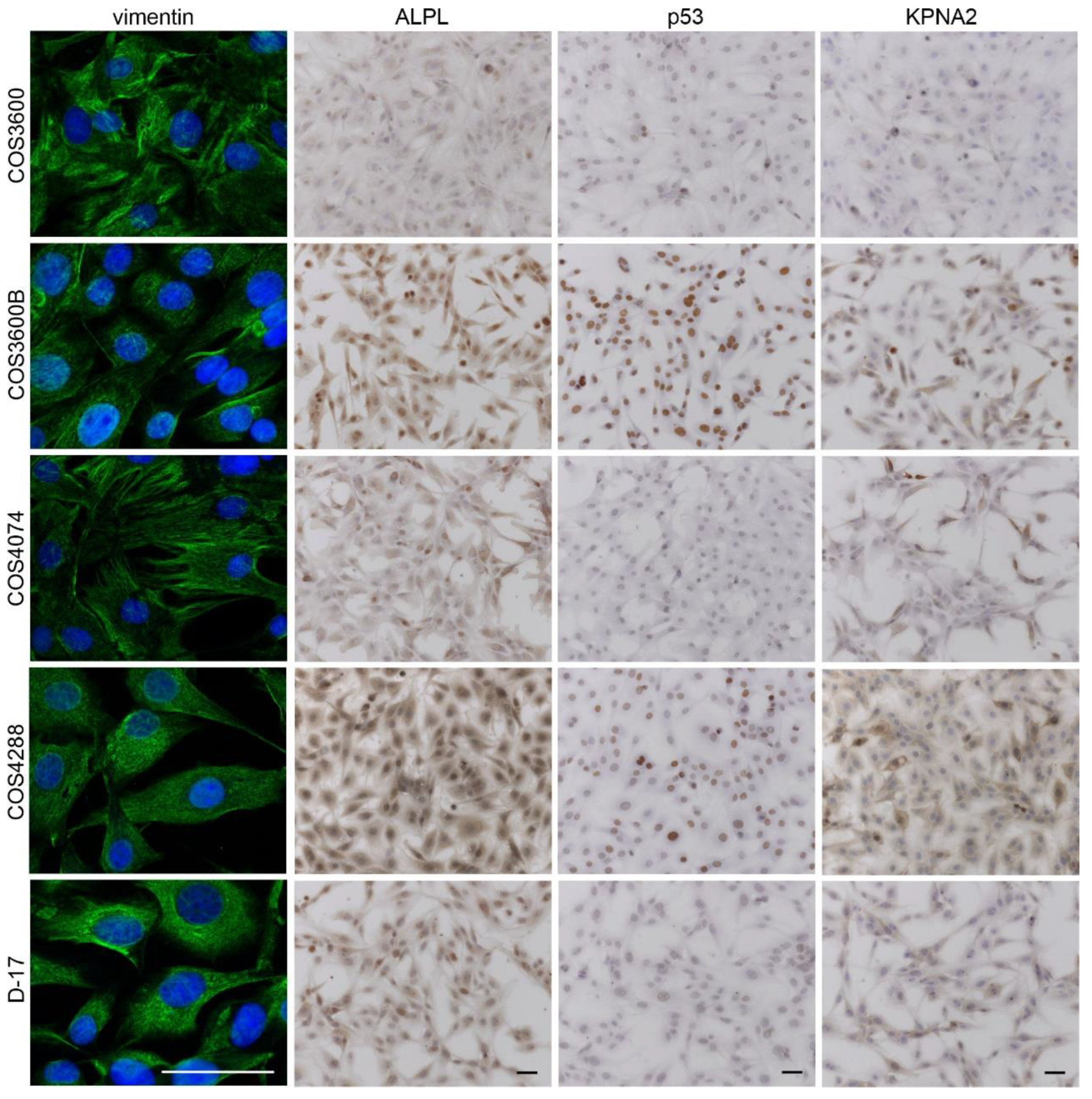
3.6. Immunohistochemical Analyses on Cell Monolayers
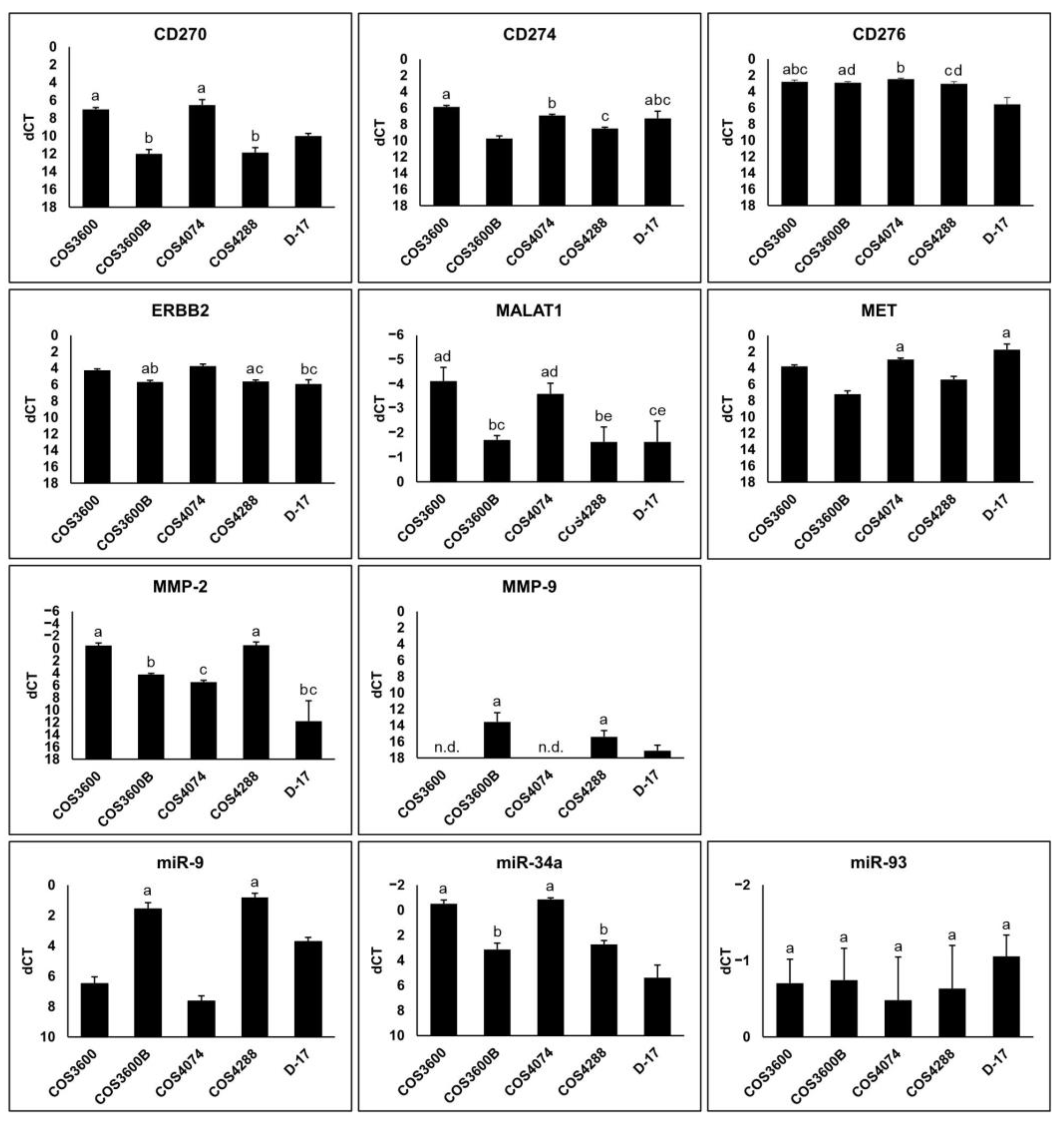
3.7. Reverse Transcription Quantitative PCR Analysis
3.8. Spheroid Formation
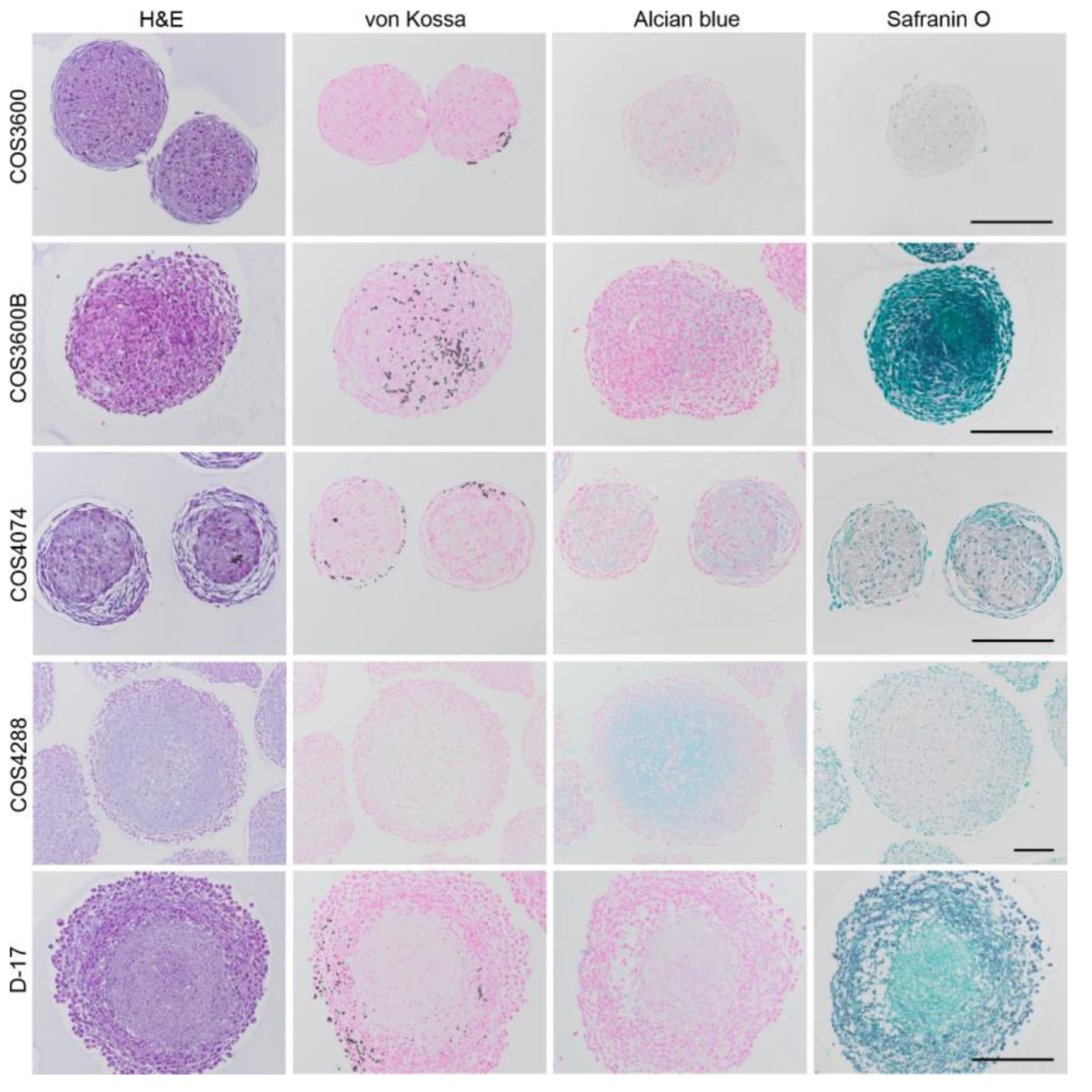
3.9. Histochemical Analyses of 3D Spheroids
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jongeward, S.J. Primary bone tumors. Veter-Clin. North Am. Small Anim. Pr. 1985, 15, 609–641. [Google Scholar] [CrossRef]
- Szewczyk, M.; Lechowski, R.; Zabielska, K. What do we know about canine osteosarcoma treatment?—Review. Veter-Res. Commun. 2015, 39, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Selmic, L.; Burton, J.; Thamm, D.; Withrow, S.; Lana, S. Comparison of carboplatin and doxorubicin-based chemotherapy protocols in 470 dogs after amputation for treatment of appendicular osteosarcoma. J. Veter-Intern. Med. 2014, 28, 554–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poon, A.; Matsuyama, A.; Mutsaers, A.J. Recent and current clinical trials in canine appendicular osteosarcoma. Can. Vet. J. 2020, 61, 301–308. [Google Scholar]
- Fan, T.M.; Roberts, R.D.; Lizardo, M.M. Understanding and modeling metastasis biology to improve therapeutic strategies for combating osteosarcoma progression. Front. Oncol. 2020, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilk, S.; Zabielska-Koczywąs, K. Molecular mechanisms of canine osteosarcoma metastasis. Int. J. Mol. Sci. 2021, 22, 3639. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.; Ren, L.; Huang, S.; Berger, E.; Bardales, K.; Mannheimer, J.; Mazcko, C.; LeBlanc, A. Canine and murine models of osteosarcoma. Veter-Pathol. 2022, 59, 399–414. [Google Scholar] [CrossRef]
- Higuchi, T.; Igarashi, K.; Yamamoto, N.; Hayashi, K.; Kimura, H.; Miwa, S.; Bouvet, M.; Tsuchiya, H.; Hoffman, R.M. Osteosarcoma patient-derived orthotopic xenograft (PDOX) models used to identify novel and effective therapeutics: A review. Anticancer Res. 2021, 41, 5865–5871. [Google Scholar] [CrossRef]
- Landuzzi, L.; Manara, M.; Lollini, P.-L.; Scotlandi, K. Patient derived xenografts for genome-driven therapy of osteosarcoma. Cells 2021, 10, 416. [Google Scholar] [CrossRef]
- Brulin, B.; Nolan, J.C.; Marangon, T.; Kovacevic, M.; Chatelais, M.; Meheust, P.; Abadie, J.; Le Nail, L.-R.; Rosset, P.; Brennan, M.; et al. Evaluation of the chemotherapy drug response using organotypic cultures of osteosarcoma tumours from mice models and canine patients. Cancers 2021, 13, 4890. [Google Scholar] [CrossRef]
- Ren, L.; Mendoza, A.; Zhu, J.; Briggs, J.W.; Halsey, C.; Hong, E.S.; Burkett, S.S.; Morrow, J.J.; Lizardo, M.M.; Osborne, T.; et al. Characterization of the metastatic phenotype of a panel of established osteosarcoma cells. Oncotarget 2015, 6, 29469–29481. [Google Scholar] [CrossRef] [Green Version]
- Laschi, M.; Bernardini, G.; Geminiani, M.; Ghezzi, L.; Amato, L.; Braconi, D.; Millucci, L.; Frediani, B.; Spreafico, A.; Franchi, A.; et al. Establishment of four new human primary cell cultures from chemo-naïve Italian osteosarcoma patients. J. Cell. Physiol. 2015, 230, 2718–2727. [Google Scholar] [CrossRef]
- Meyer, F.R.L.; Walter, I. Establishment and characterization of new canine and feline osteosarcoma primary cell lines. Veter-Sci. 2016, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Palmini, G.; Zonefrati, R.; Romagnoli, C.; Aldinucci, A.; Mavilia, C.; Leoncini, G.; Franchi, A.; Capanna, R.; Brandi, M.L. Establishment and characterization of a human small cell osteosarcoma cancer stem cell line: A new possiblein vitromodel for discovering small cell osteosarcoma biology. Stem Cells Int. 2016, 2016, 3042198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massimini, M.; DE Maria, R.; Malatesta, D.; Romanucci, M.; D’Anselmo, A.; Della Salda, L.; Marcella, M. Establishment of three-dimensional canine osteosarcoma cell lines showing vasculogenic mimicry and evaluation of biological properties after treatment with 17-AAG. Veter-Comp. Oncol. 2019, 17, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Robles, H.; Franks, K.; Pool, R.; Miller, T. Characterization of five newly derived canine osteosarcoma cell lines. BMC Veter-Res. 2019, 15, 357. [Google Scholar] [CrossRef]
- Ren, K.; Ni, Y.; Li, X.; Wang, C.; Chang, Q.; Li, Y.; Gao, Z.; Wu, S.; Shi, X.; Song, J.; et al. Expression profiling of long noncoding RNAs associated with vasculogenic mimicry in osteosarcoma. J. Cell. Biochem. 2019, 120, 12473–12488. [Google Scholar] [CrossRef]
- Fan, Z.; Huang, G.; Zhao, J.; Li, W.; Lin, T.; Su, Q.; Yin, J.; Shen, J. Establishment and characterization of a highly metastatic human osteosarcoma cell line from osteosarcoma lung metastases. J. Bone Oncol. 2021, 29, 100378. [Google Scholar] [CrossRef]
- Tao, Y.; Ou, Y.; Yin, H.; Chen, Y.; Zhong, S.; Gao, Y.; Zhao, Z.; He, B.; Huang, Q.; Deng, Q. Establishment and characterization of human osteosarcoma cells resistant to pyropheophorbide-α methyl ester-mediated photodynamic therapy. Int. J. Oncol. 2017, 51, 1427–1438. [Google Scholar] [CrossRef] [Green Version]
- Fenger, J.M.; London, C.A.; Kisseberth, W.C. Canine osteosarcoma: A naturally occurring disease to inform pediatric oncology. ILAR J. 2014, 55, 69–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, S.; Dunning, M.D.; de Brot, S.; Grau-Roma, L.; Mongan, N.P.; Rutland, C.S. Comparative review of human and canine osteosarcoma: Morphology, epidemiology, prognosis, treatment and genetics. Acta Vet. Scand. 2017, 59, 71. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, L.; Scotlandi, K.; Pettinari, I.; Benassi, M.; Porcellato, I.; Pazzaglia, L. MiRNAs in canine and human osteosarcoma: A highlight review on comparative biomolecular aspects. Cells 2021, 10, 428. [Google Scholar] [CrossRef] [PubMed]
- Walter, I.; Burger, S.; Stargardt, M.; Kummer, S.; Wieser, M. VetBiobank, vetmeduni Vienna: A bioresource for clinical animal biospecimens. Open J. Bioresour. 2020, 7, 9. [Google Scholar] [CrossRef]
- Slayter, M.V.; Boosinger, T.R.; Pool, R.R.; Dammrich, K.; Misdorp, W.; Larsen, S. Histological Classification of Bone and Joint Tumors of Domestic Animals; Armed Forces Institute of Pathology, American Registry of Pathology and World Health Organization Collaborating Center for Worldwide Reference on Comparative Oncology: Washington, DC, USA, 1994. [Google Scholar]
- Loukopoulos, P.; Robinson, W. Clinicopathological relevance of tumour grading in canine osteosarcoma. J. Comp. Pathol. 2007, 136, 65–73. [Google Scholar] [CrossRef]
- Leitner, N.; Hlavatý, J.; Ertl, R.; Gabner, S.; Fuchs-Baumgartinger, A.; Walter, I. Lipid droplets and perilipins in canine osteosarcoma. Investigations on tumor tissue, 2D and 3D cell culture models. Veter-Res. Commun. 2022, 46, 1175–1193. [Google Scholar] [CrossRef]
- Vojdani, A.; Choppa, P.; Tagle, C.; Andrin, R.; Samimi, B.; Lapp, C. Detection of mycoplasmagenus and mycoplasma fermentansby PCR in patients with chronic fatigue syndrome. FEMS Immunol. Med. Microbiol. 1998, 22, 355–365. [Google Scholar] [CrossRef]
- Lin, L.-C.; Elkashty, O.; Ramamoorthi, M.; Trinh, N.; Liu, Y.; Sunavala-Dossabhoy, G.; Pranzatelli, T.; Michael, D.G.; Chivasso, C.; Perret, J.; et al. Cross-contamination of the human salivary gland HSG cell line with HeLa cells: A STR analysis study. Oral Dis. 2018, 24, 1477–1483. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In vitro cell migration and invasion assays. J. Vis. Exp. 2014, 88, e51046. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.; Flaherty, P. Corning Assay Methods. Protocol Cell Invasion Assay. Available online: https://www.corning.com/catalog/cls/documents/protocols/protocol_DL_031_Cell_Invasion_Assay.pdf.2012–2018CorningIncorporated6/18CLS-DL-CC-031REV (accessed on 27 July 2022).
- Romeis, B. Mikroskopische Technik. 17. Aufl; Böck, P., Ed.; Urban and Schwarzenberg: Munich, Germany, 1989. [Google Scholar]
- Mulisch, M.; Welsch, U. (Eds.) Romeis-Mikroskopische Technik, 18th ed.; Springer Spektrum; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Gabriel, C.; Becher-Deichsel, A.; Hlavaty, J.; Mair, G.; Walter, I. The physiological expression of scavenger receptor SR-B1 in canine endometrial and placental epithelial cells and its potential involvement in pathogenesis of pyometra. Theriogenology 2016, 85, 1599–1609.e2. [Google Scholar] [CrossRef]
- Leitner, N.; Hlavaty, J.; Heider, S.; Ertl, R.; Gabriel, C.; Walter, I. Lipid droplet dynamics in healthy and pyometra-affected canine endometrium. BMC Veter-Res. 2022, 18, 221. [Google Scholar] [CrossRef]
- Saxena, S.; Shrivastava, S.; Arora, R.; Jena, S.C.; Vasu, R.K.; Srivastava, S.; Hussain, S.; Kumar, M.; Sharma, P.; Kumar, N.; et al. Development of real-time pcr assays for detecting matrix metalloproteinases-2 & 9 over-expression in canine mammary tumours. Adv. Anim. Veter-Sci. 2016, 4, 342–345. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Busk, P.K. A tool for design of primers for microRNA-specific quantitative RT-qPCR. BMC Bioinform. 2014, 15, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riester, S.M.; Torres-Mora, J.; Dudakovic, A.; Camilleri, E.T.; Wang, W.; Xu, F.; Thaler, R.R.; Evans, J.M.; Zwartbol, R.; Bruijn, I.H.B.-D.; et al. Hypoxia-related microRNA-210 is a diagnostic marker for discriminating osteoblastoma and osteosarcoma. J. Orthop. Res. 2016, 35, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebhard, C.; Gabriel, C.; Walter, I. Morphological and immunohistochemical characterization of canine osteosarcoma spheroid cell cultures. Anat. Histol. Embryol. 2015, 45, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Rickel, K.; Fang, F.; Tao, J. Molecular genetics of osteosarcoma. Bone 2017, 102, 69–79. [Google Scholar] [CrossRef]
- Tuohy, J.L.; Shaevitz, M.H.; Garrett, L.D.; Ruple, A.; Selmic, L.E. Demographic characteristics, site and phylogenetic distribution of dogs with appendicular osteosarcoma: 744 dogs (2000–2015). PLoS ONE 2019, 14, e0223243. [Google Scholar] [CrossRef] [Green Version]
- Ayers, J.; Milner, R.J.; Cortés-Hinojosa, G.; Riva, A.; Bechtel, S.; Sahay, B.; Cascio, M.; Lejeune, A.; Shiomitsu, K.; Souza, C.; et al. Novel application of single-cell next-generation sequencing for determination of intratumoral heterogeneity of canine osteosarcoma cell lines. J. Veter-Diagn. Investig. 2021, 33, 261–278. [Google Scholar] [CrossRef]
- Maeda, J.; Yurkon, C.R.; Fujisawa, H.; Kaneko, M.; Genet, S.C.; Roybal, E.J.; Rota, G.W.; Saffer, E.R.; Rose, B.J.; Hanneman, W.H.; et al. Genomic instability and telomere fusion of canine osteosarcoma cells. PLoS ONE 2012, 7, e43355. [Google Scholar] [CrossRef] [PubMed]
- Masters, J.R.; Thomson, J.A.; Daly-Burns, B.; Reid, Y.A.; Dirks, W.G.; Packer, P.; Toji, L.H.; Ohno, T.; Tanabe, H.; Arlett, C.F.; et al. Short tandem repeat profiling provides an international reference standard for human cell lines. Proc. Natl. Acad. Sci. USA 2001, 98, 8012–8017. [Google Scholar] [CrossRef] [Green Version]
- Parson, W.; Kirchebner, R.; Mühlmann, R.; Renner, K.; Kofler, A.; Schmidt, S.; Kofler, R. Cancer cell line identification by short tandem repeat profiling: Power and limitations. FASEB J. 2004, 19, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, A.W.; Salmon, S.E.; Kim, M.B.; Trent, J.M.; Soehnlen, B.J.; Alberts, D.S.; Schmidt, H.J. Direct cloning of human ovarian carcinoma cells in agar. Cancer Res. 1978, 38, 3438–3444. [Google Scholar]
- Legare, M.E.; Bush, J.; Ashley, A.K.; Kato, T.; Hanneman, W.H. Cellular and phenotypic characterization of canine osteosarcoma cell lines. J. Cancer 2011, 2, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Aapro, M.S.; Eliason, J.F.; Krauer, F.; Alberto, P. Colony formation in vitro as a prognostic indicator for primary breast cancer. J. Clin. Oncol. 1987, 5, 890–896. [Google Scholar] [CrossRef]
- Page, R.H.; Tilchen, E.J.; Davis, H.L. Effects of tumor cell viability and inoculation density on growth parameters in the human tumor, soft-agar clonogenic assay. Cancer Res. 1988, 48, 3944–3948. [Google Scholar]
- Devarajan, E.; Song, Y.-H.; Krishnappa, S.; Alt, E. Epithelial-mesenchymal transition in breast cancer lines is mediated through PDGF-D released by tissue-resident stem cells. Int. J. Cancer 2011, 131, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Fenger, J.M.; Roberts, R.D.; Iwenofu, O.H.; Bear, M.D.; Zhang, X.; Couto, J.I.; Modiano, J.F.; Kisseberth, W.C.; London, C.A. MiR-9 is overexpressed in spontaneous canine osteosarcoma and promotes a metastatic phenotype including invasion and migration in osteoblasts and osteosarcoma cell lines. BMC Cancer 2016, 16, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, C.M.; Yu, P.Y.; Zhang, X.; Yilmaz, A.S.; London, C.A.; Fenger, J.M. MiR-34a regulates the invasive capacity of canine osteosarcoma cell lines. PLoS ONE 2018, 13, e0190086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riggs, J.L.; McAllister, R.M.; Lennette, E.H. Immunofluorescent Studies of RD-114 Virus Replication in Cell Culture. J. Gen. Virol. 1974, 25, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Modesto, P.; Fernandez, J.; Martini, I.; Zoccola, R.; Pugliano, M.; Ciucis, C.; Goria, M.; Ferrari, A.; Razzuoli, E. Characterization of D-17 canine osteosarcoma cell line and evaluation of its ability to response to infective stressor used as alternative anticancer therapy. Animals 2020, 10, 1981. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, X.; Zhang, Y.; Jiang, H.; Cai, X.; Yan, X.; Huang, Z.; Mo, F.; Yang, W.; Yang, C.; et al. Establishment and characterization of a novel osteosarcoma cell line: CHOS. J. Orthop. Res. 2016, 34, 2116–2125. [Google Scholar] [CrossRef] [Green Version]
- Barger, A.; Graca, R.; Bailey, K.; Messick, J.; de Lorimier, L.-P.; Fan, T.; Hoffmann, W. Use of alkaline phosphatase staining to differentiate canine osteosarcoma from other vimentin-positive tumors. Veter-Pathol. 2005, 42, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Borrero, L.J.H.; El-Deiry, W.S. Tumor suppressor p53: Biology, signaling pathways, and therapeutic targeting. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2021, 1876, 188556. [Google Scholar] [CrossRef]
- Muller, P.A.J.; Vousden, K.H. p53 mutations in cancer. Nat. Cell Biol. 2013, 15, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Kirpensteijn, J.; Kik, M.; Teske, E.; Rutteman, G.R. TP53 gene mutations in canine osteosarcoma. Veter-Surg. 2008, 37, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.S.; Jaworski, L.; Kisseberth, W.C. Immunohistochemical detection of p53, PTEN, Rb, and p16 in canine osteosarcoma using tissue microarray. J. Veter-Diagn. Investig. 2018, 30, 504–509. [Google Scholar] [CrossRef] [Green Version]
- Maltzman, W.; Czyzyk, L. UV irradiation stimulates levels of p53 cellular tumor antigen in nontransformed mouse cells. Mol. Cell. Biol. 1984, 4, 1689–1694. [Google Scholar] [CrossRef]
- Maki, C.G.; Howley, P.M. Ubiquitination of p53 and p21 is differentially affected by ionizing and UV radiation. Mol. Cell. Biol. 1997, 17, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Gola, C.; Giannuzzi, D.; Rinaldi, A.; Iussich, S.; Modesto, P.; Morello, E.; Buracco, P.; Aresu, L.; De Maria, R. Genomic and transcriptomic characterization of canine osteosarcoma cell lines: A valuable resource in translational medicine. Front. Veter-Sci. 2021, 8, 666838. [Google Scholar] [CrossRef] [PubMed]
- Piaskowski, S.; Zawlik, I.; Szybka, M.; Kulczycka-Wojdala, D.; Stoczynska-Fidelus, E.; Bienkowski, M.; Robak, T.; Kusinska, R.; Jesionek-Kupnicka, D.; Kordek, R.; et al. Detection of P53 mutations in different cancer types is improved by cDNA sequencing. Oncol. Lett. 2010, 1, 717–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camplejohn, R.; Rutherford, J. p53 functional assays: Detecting p53 mutations in both the germline and in sporadic tumours. Cell Prolif. 2001, 34, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, X. The emerging roles of KPNA2 in cancer. Life Sci. 2020, 241, 117140. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, J.; Wei, Q.; Wang, Y. KPNA2 expression is a potential marker for differential diagnosis between osteosarcomas and other malignant bone tumor mimics. Diagn. Pathol. 2020, 15, 135. [Google Scholar] [CrossRef]
- Thanindratarn, P.; Dean, D.C.; Nelson, S.D.; Hornicek, F.J.; Duan, Z. Advances in immune checkpoint inhibitors for bone sarcoma therapy. J. Bone Oncol. 2019, 15, 100221. [Google Scholar] [CrossRef]
- Chen, C.; Xie, L.; Ren, T.; Huang, Y.; Xu, J.; Guo, W. Immunotherapy for osteosarcoma: Fundamental mechanism, rationale, and recent breakthroughs. Cancer Lett. 2020, 500, 1–10. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, J.; Chen, Y.; Kang, Y.; Liao, Z.; He, Y.; Zhang, C. Novel immunotherapies for osteosarcoma. Front. Oncol. 2022, 12, 830546. [Google Scholar] [CrossRef]
- Cascio, M.J.; Whitley, E.M.; Sahay, B.; Cortes-Hinojosa, G.; Chang, L.-J.; Cowart, J.; Salute, M.; Sayour, E.; Dark, M.; Sandoval, Z.; et al. Canine osteosarcoma checkpoint expression correlates with metastasis and T-cell infiltrate. Vet. Immunol. Immunopathol. 2021, 232, 110169. [Google Scholar] [CrossRef]
- McEachron, T.A.; Triche, T.J.; Sorenson, L.; Parham, D.M.; Carpten, J.D. Profiling targetable immune checkpoints in osteosarcoma. Oncoimmunology 2018, 7, e1475873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacEwen, E.G.; Kutzke, J.; Carew, J.; Pastor, J.; Schmidt, J.A.; Tsan, R.; Thamm, D.H.; Radinsky, R. C-Met tyrosine kinase receptor expression and function in human and canine osteosarcoma cells. Clin. Exp. Metastasis 2003, 20, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Flint, A.F.; U’Ren, L.; Legare, M.E.; Withrow, S.J.; Dernell, W.; Hanneman, W.H. Overexpression of the erbB-2 Proto-oncogene in Canine Osteosarcoma Cell Lines and Tumors. Veter-Pathol. 2004, 41, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.E.; Ba, M.B.; Kim, M.Y.; Lin, J.; Bs, S.P.; Gorlick, R.; Geller, D.S. Cell surface receptor expression patterns in osteosarcoma. Cancer 2011, 118, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Gorlick, S.; Barkauskas, D.A.; Krailo, M.; Piperdi, S.; Sowers, R.; Gill, J.; Geller, D.; Randall, R.L.; Janeway, K.; Schwartz, C.; et al. HER-2 expression is not prognostic in osteosarcoma; a children’s oncology group prospective biology study. Pediatr. Blood Cancer 2014, 61, 1558–1564. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ma, Y.-H.; Sun, Z.-Z.; Rui, Y.-J.; Yin, Q.-D.; Song, S.; Wei, X.-M.; Liu, J.; Liu, X.-G.; Hu, K.-J. Effect of c-erbB2 overexpression on prognosis in osteosarcoma: Evidence from eight studies. Tumor Biol. 2014, 35, 8939–8943. [Google Scholar] [CrossRef]
- Fieten, H.; Spee, B.; Ijzer, J.; Kik, M.J.; Penning, L.C.; Kirpensteijn, J. Expression of hepatocyte growth factor and the proto-oncogenic receptor c-met in canine osteosarcoma. Veter-Pathol. 2009, 46, 869–877. [Google Scholar] [CrossRef] [Green Version]
- De Maria, R.; Miretti, S.; Iussich, S.; Olivero, M.; Morello, E.; Bertotti, A.; Christensen, J.G.; Biolatti, B.; Levine, R.A.; Buracco, P.; et al. Metoncogene activation qualifies spontaneous canine osteosarcoma as a suitable pre-clinical model of human osteosarcoma. J. Pathol. 2009, 218, 399–408. [Google Scholar] [CrossRef]
- de Almeida, L.G.; Thode, H.; Eslambolchi, Y.; Chopra, S.; Young, D.; Gill, S.; Devel, L.; Dufour, A. Matrix metalloproteinases: From molecular mechanisms to physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 2022, 74, 714–770. [Google Scholar] [CrossRef]
- Siddhartha, R.; Garg, M. Molecular and clinical insights of matrix metalloproteinases into cancer spread and potential therapeutic interventions. Tox Appl. Pharm. 2021, 426, 115593. [Google Scholar] [CrossRef]
- Niland, S.; Riscanevo, A.X.; Eble, J.A. matrix metalloproteinases shape the tumor microenvironment in cancer progression. Int. J. Mol. Sci. 2021, 23, 146. [Google Scholar] [CrossRef]
- Loukopoulos, P.; Mungall, B.; Straw, R.C.; Thornton, J.R.; Robinson, W.F. Matrix metalloproteinase-2 and -9 Involvement in Canine Tumors. Veter-Pathol. 2003, 40, 382–394. [Google Scholar] [CrossRef]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and biological attributes of matrix metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzaneh, M.; Najafi, S.; Anbiyaee, O.; Azizidoost, S.; Khoshnam, S.E. LncRNA MALAT1-related signaling pathways in osteosarcoma. Clin. Transl. Oncol. 2022, 25, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhang, T.; Chen, B. Long non-coding RNA metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) promotes proliferation and metastasis of osteosarcoma cells by targeting c-Met and SOX4 via miR-34a/c-5p and miR-449a/b. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 1410–1422. [Google Scholar] [CrossRef]
- Zhang, J.; Piao, C.-D.; Ding, J.; Li, Z.-W. LncRNA MALAT1 facilitates lung metastasis of osteosarcomas through miR-202 sponging. Sci. Rep. 2020, 10, 12757. [Google Scholar] [CrossRef]
- Liu, J.; Shang, G. The roles of noncoding RNAs in the development of osteosarcoma stem cells and potential therapeutic targets. Front. Cell Dev. Biol. 2022, 10, 773038. [Google Scholar] [CrossRef]
- Song, X.J.; Bi, M.C.; Zhu, Q.S.; Liu, X.L. The emerging role of lncRNAs in the regulation of osteosarcoma stem cells. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 966–974. [Google Scholar] [CrossRef]
- Liu, C.; Han, X.; Li, B.; Huang, S.; Zhou, Z.; Wang, Z.; Wang, W. MALAT-1 is associated with the doxorubicin resistance in U-2OS osteosarcoma cells. Cancer Manag. Res. 2021, 13, 6879–6889. [Google Scholar] [CrossRef]
- Li, F.; Li, X.; Qiao, L.; Liu, W.; Xu, C.; Wang, X. MALAT1 regulates miR-34a expression in melanoma cells. Cell Death Dis. 2019, 10, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Ikeda, S.; Iwasaki, T.; Tsumura, H. microRNA-93 promotes cell proliferation via targeting of PTEN in Osteosarcoma cells. J. Exp. Clin. Cancer Res. 2015, 34, 76. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, J.; Meng, F.; Zhu, H.; Yan, H.; Guo, Y.; Zhang, S. MicroRNA-93 promotes the tumorigenesis of osteosarcoma by targeting TIMP2. Biosci. Rep. 2019, 39, BSR20191237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, B.; Henriques, A.C.; Silva, P.M.A.; Bousbaa, H. Three-dimensional spheroids as in vitro preclinical models for cancer research. Pharmaceutics 2020, 12, 1186. [Google Scholar] [CrossRef]
- El Atat, O.; Farzaneh, Z.; Pourhamzeh, M.; Taki, F.; Abi-Habib, R.; Vosough, M.; El-Sibai, M. 3D modeling in cancer studies. Hum. Cell 2022, 35, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Langhans, S.A. Three-dimensional in vitro cell culture models in drug discovery and drug repositioning. Front. Pharmacol. 2018, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.S.; Barros, A.S.; Costa, E.C.; Moreira, A.F.; Correia, I.J. 3D tumor spheroids as in vitro models to mimic in vivo human solid tumors resistance to therapeutic drugs. Biotechnol. Bioeng. 2018, 116, 206–226. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Brown, P.C.; Chow, E.C.; Ewart, L.; Ferguson, S.S.; Fitzpatrick, S.; Freedman, B.S.; Guo, G.L.; Hedrich, W.; Heyward, S.; et al. 3D cell culture models: Drug pharmacokinetics, safety assessment, and regulatory consideration. Clin. Transl. Sci. 2021, 14, 1659–1680. [Google Scholar] [CrossRef]
- Thanindratarn, P.; Li, X.; Dean, D.C.; Nelson, S.D.; Hornicek, F.J.; Duan, Z. Establishment and Characterization of a Recurrent Osteosarcoma Cell Line: OSA 1777. J. Orthop. Res. 2019, 38, 902–910. [Google Scholar] [CrossRef]
- Lin, R.-Z.; Chang, H.-Y. Recent advances in three-dimensional multicellular spheroid culture for biomedical research. Biotechnol. J. 2008, 3, 1172–1184. [Google Scholar] [CrossRef]
- Thompson, K.G.; Dittmer, K.E. Tumors of Bone. In Tumors in Domestic Animals, 5th ed.; Meuten, D.J., Ed.; Wiley: New York, NY, USA, 2017; p. 6. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COS3600 | COS3600B | COS4074 | COS4288 | |
|---|---|---|---|---|
| Amelogenin | Y/X | Y/X | Y/X | X/X |
| AHT 121 | 98/100 | 98/100 | 96/102 | 102/106 |
| AHT 137 | 145/153 | 153/153 | 147/147 | 149/149 |
| AHTH 130 | 129/129 | 129/129 | 119/127 | 123/129 |
| AHTH 171 | 219/233 | 219/233 | 225/231 | 223/225 |
| AHTH 260 | 238/244 | 238/238 | 246/246 | 246/254 |
| AHTK 211 | 89/91 | 89/89 | 89/95 | 95/95 |
| AHTK 253 | 288/292 | 288/292 | 286/286 | 284/284 |
| CXX 279 | 116/124 | 124/124 | 126/126 | 124/124 |
| FH 2054 | 160/172 | 160/172 | 152/156 | 164/164 |
| FH 2848 | 232/240 | 232/232 | 238/238 | 238/244 |
| INRA 21 | 91/101 | 91/91 | 95/97 | 95/95 |
| INU 005 | 124/124 | 124/124 | 124/124 | 124/124 |
| INU 030 | 144/150 | 144/150 | 144/144 | 144/144 |
| INU 055 | 210/210 | 210/210 | 210/218 | 218/218 |
| REN 105 L 03 | 231/241 | 241/241 | 231/233 | 235/235 |
| REN 162 C 04 | 200/206 | 200/206 | 202/206 | 200/202 |
| REN 169 D 01 | 216/218 | 216/218 | 216/216 | 212/216 |
| REN 169 O 18 | 162/168 | 168/168 | 162/172 | 168/168 |
| REN 247 M 23 | 272/272 | 272/272 | 268/268 | 268/268 |
| REN 54 P 11 | 238/238 | 238/238 | 232/234 | 222/226 |
| REN 64 E 19 | 139/147 | 139/147 | 149/149 | 147/147 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leitner, N.; Ertl, R.; Gabner, S.; Fuchs-Baumgartinger, A.; Walter, I.; Hlavaty, J. Isolation and Characterization of Novel Canine Osteosarcoma Cell Lines from Chemotherapy-Naïve Patients. Cells 2023, 12, 1026. https://doi.org/10.3390/cells12071026
Leitner N, Ertl R, Gabner S, Fuchs-Baumgartinger A, Walter I, Hlavaty J. Isolation and Characterization of Novel Canine Osteosarcoma Cell Lines from Chemotherapy-Naïve Patients. Cells. 2023; 12(7):1026. https://doi.org/10.3390/cells12071026
Chicago/Turabian StyleLeitner, Natascha, Reinhard Ertl, Simone Gabner, Andrea Fuchs-Baumgartinger, Ingrid Walter, and Juraj Hlavaty. 2023. "Isolation and Characterization of Novel Canine Osteosarcoma Cell Lines from Chemotherapy-Naïve Patients" Cells 12, no. 7: 1026. https://doi.org/10.3390/cells12071026
APA StyleLeitner, N., Ertl, R., Gabner, S., Fuchs-Baumgartinger, A., Walter, I., & Hlavaty, J. (2023). Isolation and Characterization of Novel Canine Osteosarcoma Cell Lines from Chemotherapy-Naïve Patients. Cells, 12(7), 1026. https://doi.org/10.3390/cells12071026