
HAT1: Landscape of Biological Function and Role in Cancer


 ,
,  and
and 
Abstract
:1. Introduction
2. Discovery and Enzymatic Activities of HAT1
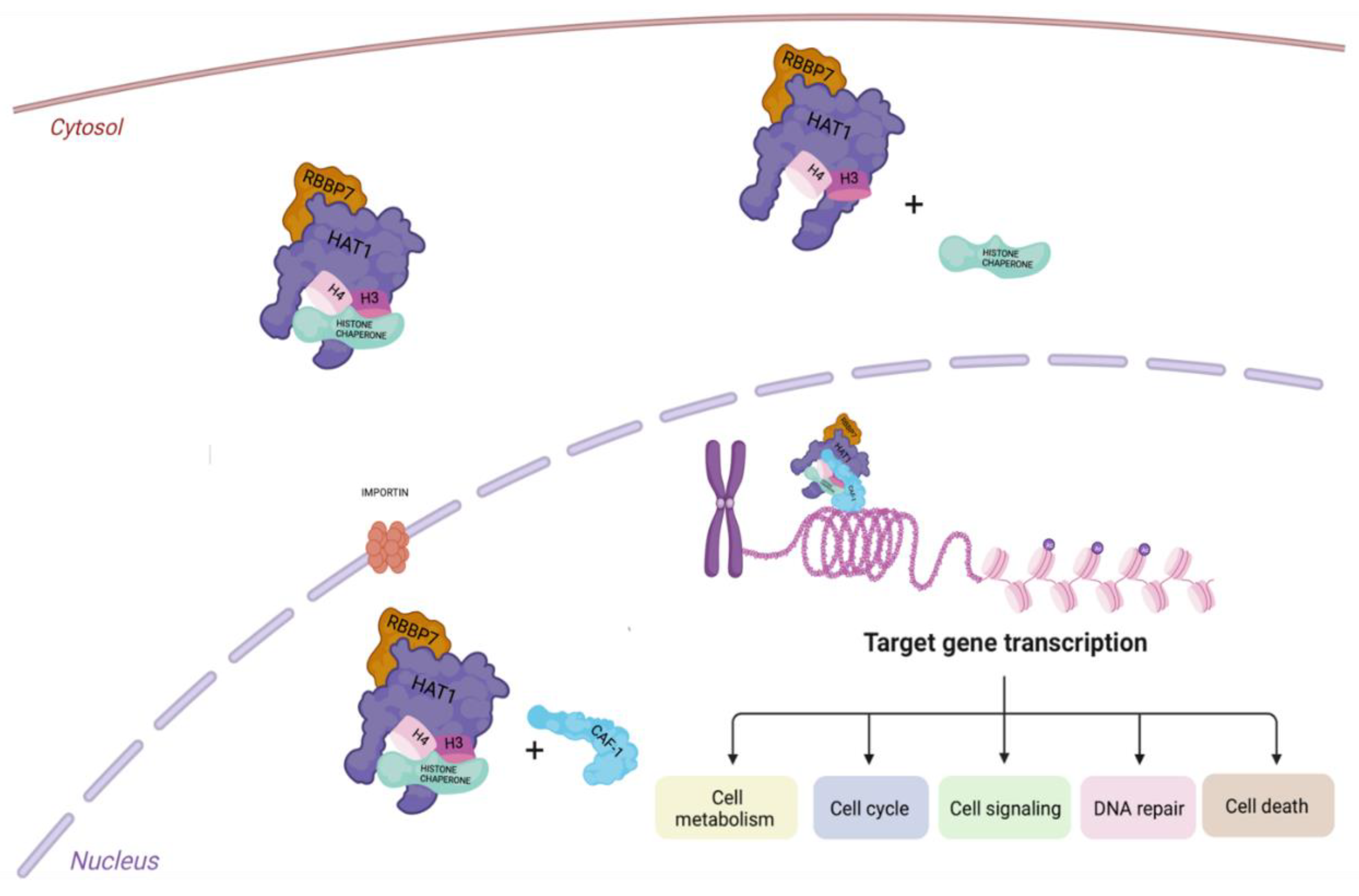
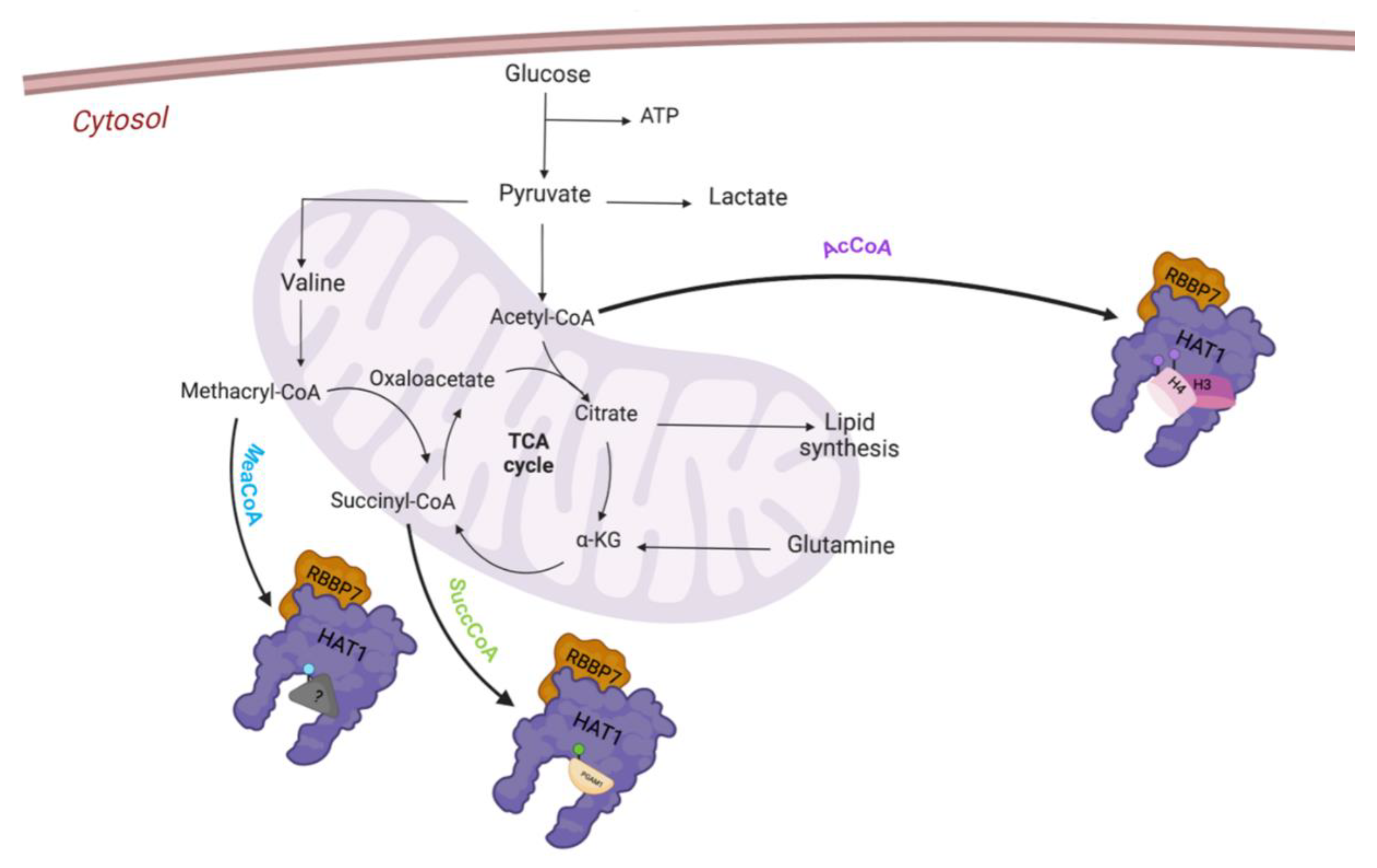
3. HAT1 Biological Activity
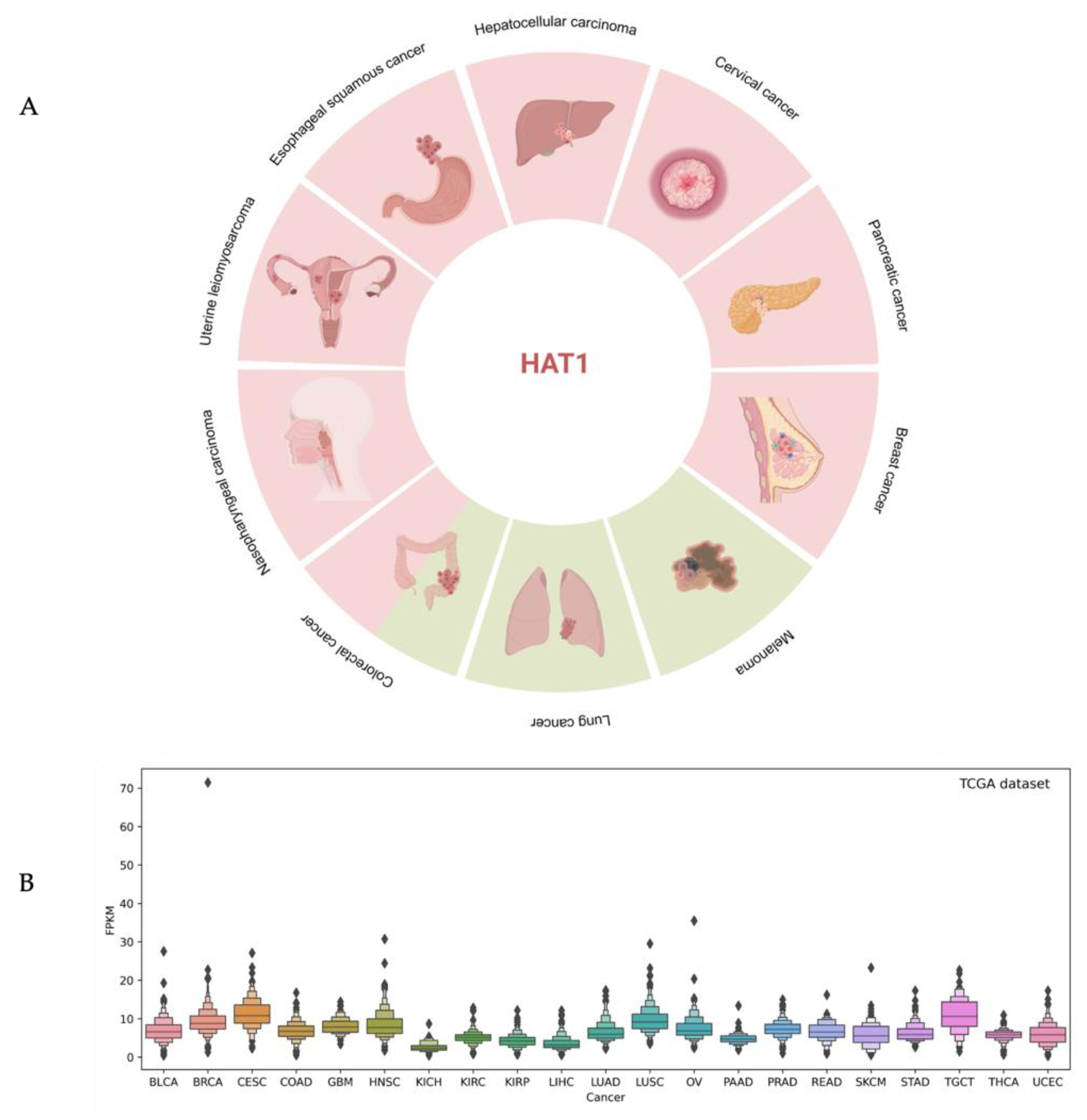
4. HAT1 in Cancer
5. Pharmacological Response
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
List of Abbreviations
| ABCA1 | ATP-binding cassette transporter A1 |
| AC | Actinic cheilitis |
| Acetyl-CoA | Acetyl-coenzyme A |
| ACLY | ATP citrate lyase |
| ACSS2 | Acyl-CoA synthetase short chain family member 2 |
| Asf1 | Anti-silencing factor 1 |
| ATR | Rad3-related protein |
| Baz1a | Bromodomain adjacent to zinc finger domain protein 1A |
| BC | Breast cancer |
| BCL2 | B cell-lymphoma 2 |
| BCL2L12 | BCL2-like protein-12 |
| BRAF | v-raf murine sarcoma viral oncogene homolog |
| BRD | Bromodomain |
| Brd3 | BRD-containing Protein 3 |
| Brg1 | Brahma-related gene-1 |
| CAF-1 | Chromatin assembly factor 1 |
| CaMK2 | Calcium/calmodulin-dependent protein kinase II |
| CC | Cervical cancer |
| cccDNA | Covalently closed circular DNA |
| CCR4 | Chemokine receptor 4 |
| CENP-A | Centromere protein A |
| CLIC1 | Chloride intracellular channel 1 |
| CRC | Colorectal cancer |
| CRPC | Castration-resistant prostate cancer |
| DLBCL | Diffuse large B-cell lymphoma |
| DSB | DNA double-strand breaks |
| EGF | Epidermal growth factor |
| ESC | Esophageal squamous cancer |
| EZH2 | Enhancer of zeste homolog 2 |
| FAS | Fas Cell Surface Death Receptor) |
| FOXP3 | Forkhead box P3 |
| GNAT | GCN5-related N-acetyltransferases |
| GST | Glutathione S-transferase |
| HAT | Histone Acetyltrasferase |
| HAT1 | Histone Acetyltrasferase 1 |
| HAT4 | Histone Acetyltrasferase 4 |
| HBc | Hepatitis B core |
| Hbo1 | Histone Acetyltransferase Binding to ORC1 |
| HBV | Hepatitis B virus |
| HCC | Hepatocellular carcinoma |
| HDAC | Histone Deacetylase |
| HIRA | Histone cell cycle regulator |
| HIV | Human immunodeficiency virus |
| Kac | Lysine acetylation |
| KAT | Lysine Acetyltransferases |
| KDAC | Lysine Deacetylase |
| Kmea | Lysine Methacrylation |
| LMS | Uterine leiomyosarcoma |
| lncRNA | Long non-coding RNAs |
| LSCC | Lip squamous cell carcinoma |
| MC-CoA | Methacryl-coenzyme A |
| MEF | Mouse embryonic fibroblasts |
| NASP | Nuclear autoantigenic sperm protein |
| NF-κB | Nuclear factor kappa B |
| NOX5 | NADPH oxidase 5 |
| NPC | Nasopharyngeal carcinoma |
| NRCF | Nuclear receptor cofactors |
| NRF1 | Nuclear respiratory factor 1 |
| NRF2 | Nuclear respiratory factor 2 |
| p300/CBP | Protein 300/CREB-binding protein |
| PAR2 | Protease-activated receptor-2 |
| PARP | Poly(ADP-ribose) polymerase |
| PC | Pancreatic cancer |
| PCNA | Proliferating cell nuclear antigen |
| PD-L1 | Programmed death-ligand 1 |
| PGAM1 | Phosphoglycerate mutase 1 |
| PGC-1α | Peroxisome proliferator activated receptor gamma coactivator 1 |
| PLZF | Transcriptional regulator promyelocytic leukemia zinc finger |
| PTCL | Peripheral T-cell lymphoma |
| PTM | Post-transcriptional modification |
| PVT1 | Plasmacytoma variant translocation 1 |
| Rbp46 | Retinoblastoma-associated protein 46 |
| Rbp48 | Retinoblastoma-associated protein 48 |
| SIRT7 | Sirtuin 7 |
| STAT5 | Signal activator of transcription 5 |
| TF | Basal transcription factor |
| TLR | Toll-like receptor |
| TNF | Tumor necrosis factor receptor |
| UBR4 | Ubiquitin protein ligase E3 component N-recognin 4 |
| UCP2 | Uncoupling protein 2 |
| UCP3 | Uncoupling protein 3 |
| γH2AX | Gamma-H2A family member X |
References
- Portela, A.; Esteller, M. Epigenetic modifications and human disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, C.; Kumar, C.; Gnad, F.; Nielsen, M.L.; Rehman, M.; Walther, T.C.; Olsen, J.V.; Mann, M. Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science 2009, 325, 834–840. [Google Scholar] [CrossRef] [Green Version]
- Seto, E.; Yoshida, M. Erasers of histone acetylation: The histone deacetylase enzymes. Cold Spring Harb. Perspect. Biol. 2014, 6, a018713. [Google Scholar] [CrossRef] [Green Version]
- Di Cerbo, V.; Schneider, R. Cancers with wrong HATs: The impact of acetylation. Brief. Funct. Genom. 2013, 12, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Hyndman, K.A.; Knepper, M.A. Dynamic regulation of lysine acetylation: The balance between acetyltransferase and deacetylase activities. Am. J. Physiol. Renal Physiol. 2017, 313, F842–F846. [Google Scholar] [CrossRef] [Green Version]
- Ali, I.; Conrad, R.J.; Verdin, E.; Ott, M. Lysine Acetylation Goes Global: From Epigenetics to Metabolism and Therapeutics. Chem. Rev. 2018, 118, 1216–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narita, T.; Weinert, B.T.; Choudhary, C. Functions and mechanisms of non-histone protein acetylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 156–174. [Google Scholar] [CrossRef]
- Shvedunova, M.; Akhtar, A. Modulation of cellular processes by histone and non-histone protein acetylation. Nat. Rev. Mol. Cell Biol. 2022, 23, 329–349. [Google Scholar] [CrossRef]
- Marmorstein, R.; Zhou, M.M. Writers and readers of histone acetylation: Structure, mechanism, and inhibition. Cold Spring Harb. Perspect. Biol. 2014, 6, a018762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, B.T.; Iesmantavicius, V.; Wagner, S.A.; Schölz, C.; Gummesson, B.; Beli, P.; Nyström, T.; Choudhary, C. Acetyl-phosphate is a critical determinant of lysine acetylation in E. coli. Mol. Cell 2013, 51, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Graf, L.G.; Vogt, R.; Blasl, A.T.; Qin, C.; Schulze, S.; Zühlke, D.; Sievers, S.; Lammers, M. Assays to Study Enzymatic and Non-Enzymatic Protein Lysine Acetylation In Vitro. Curr. Protoc. 2021, 1, e277. [Google Scholar] [CrossRef] [PubMed]
- Parthun, M.R. Hat1: The emerging cellular roles of a type B histone acetyltransferase. Oncogene 2007, 26, 5319–5328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parthun, M.R.; Widom, J.; Gottschling, D.E. The major cytoplasmic histone acetyltransferase in yeast: Links to chromatin replication and histone metabolism. Cell 1996, 87, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T.J.; Qin, S.; Gottschling, D.E.; Parthun, M.R. Type B histone acetyltransferase Hat1p participates in telomeric silencing. Mol. Cell Biol. 2000, 20, 7051–7058. [Google Scholar] [CrossRef] [Green Version]
- Annunziato, A.T.; Hansen, J.C. Role of histone acetylation in the assembly and modulation of chromatin structures. Gene Expr. 2000, 9, 37–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verreault, A. De novo nucleosome assembly: New pieces in an old puzzle. Genes. Dev. 2000, 14, 1430–1438. [Google Scholar] [CrossRef]
- Parthun, M.R. Histone acetyltransferase 1: More than just an enzyme? Biochim. Biophys. Acta 2012, 1819, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Verreault, A.; Kaufman, P.D.; Kobayashi, R.; Stillman, B. Nucleosomal DNA regulates the core-histone-binding subunit of the human Hat1 acetyltransferase. Curr. Biol. 1998, 8, 96–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, F.; Muñoz, F.; Schilcher, P.; Imhof, A.; Almouzni, G.; Loyola, A. Sequential establishment of marks on soluble histones H3 and H4. J. Biol. Chem. 2011, 286, 17714–17721. [Google Scholar] [CrossRef] [Green Version]
- Barman, H.K.; Takami, Y.; Nishijima, H.; Shibahara, K.; Sanematsu, F.; Nakayama, T. Histone acetyltransferase-1 regulates integrity of cytosolic histone H3-H4 containing complex. Biochem. Biophys. Res. Commun. 2008, 373, 624–630. [Google Scholar] [CrossRef]
- Chang, L.; Loranger, S.S.; Mizzen, C.; Ernst, S.G.; Allis, C.D.; Annunziato, A.T. Histones in transit: Cytosolic histone complexes and diacetylation of H4 during nucleosome assembly in human cells. Biochemistry 1997, 36, 469–480. [Google Scholar] [CrossRef]
- Wu, H.; Moshkina, N.; Min, J.; Zeng, H.; Joshua, J.; Zhou, M.M.; Plotnikov, A.N. Structural basis for substrate specificity and catalysis of human histone acetyltransferase 1. Proc. Natl. Acad. Sci. USA 2012, 109, 8925–8930. [Google Scholar] [CrossRef] [Green Version]
- Dutnall, R.N.; Tafrov, S.T.; Sternglanz, R.; Ramakrishnan, V. Structure of the histone acetyltransferase Hat1: A paradigm for the GCN5-related N-acetyltransferase superfamily. Cell 1998, 94, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Gruber, J.J.; Geller, B.; Lipchik, A.M.; Chen, J.; Salahudeen, A.A.; Ram, A.N.; Ford, J.M.; Kuo, C.J.; Snyder, M.P. HAT1 Coordinates Histone Production and Acetylation via H4 Promoter Binding. Mol. Cell 2019, 75, 711–724.e5. [Google Scholar] [CrossRef]
- Mendoza, M.; Egervari, G.; Sidoli, S.; Donahue, G.; Alexander, D.C.; Sen, P.; Garcia, B.A.; Berger, S.L. Enzymatic transfer of acetate on histones from lysine reservoir sites to lysine activating sites. Sci. Adv. 2022, 8, eabj5688. [Google Scholar] [CrossRef] [PubMed]
- Ai, X.; Parthun, M.R. The nuclear Hat1p/Hat2p complex: A molecular link between type B histone acetyltransferases and chromatin assembly. Mol. Cell 2004, 14, 195–205. [Google Scholar] [CrossRef]
- Poveda, A.; Pamblanco, M.; Tafrov, S.; Tordera, V.; Sternglanz, R.; Sendra, R. Hif1 is a component of yeast histone acetyltransferase B, a complex mainly localized in the nucleus. J. Biol. Chem. 2004, 279, 16033–16043. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Nagamatsu, N.; Kouriki, H.; Takami, Y.; Nakayama, T. Leucine zipper motif of chicken histone acetyltransferase-1 is essential for in vivo and in vitro interactions with the p48 subunit of chicken chromatin assembly factor-1. Nucleic Acids Res. 2001, 29, 629–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhof, A.; Wolffe, A.P. Purification and properties of the Xenopus Hat1 acetyltransferase: Association with the 14-3-3 proteins in the oocyte nucleus. Biochemistry 1999, 38, 13085–13093. [Google Scholar] [CrossRef] [PubMed]
- Agudelo Garcia, P.A.; Nagarajan, P.; Parthun, M.R. Hat1-Dependent Lysine Acetylation Targets Diverse Cellular Functions. J. Proteome Res. 2020, 19, 1663–1673. [Google Scholar] [CrossRef]
- Yang, G.; Yuan, Y.; Yuan, H.; Wang, J.; Yun, H.; Geng, Y.; Zhao, M.; Li, L.; Weng, Y.; Liu, Z.; et al. Histone acetyltransferase 1 is a succinyltransferase for histones and non-histones and promotes tumorigenesis. EMBO Rep. 2021, 22, e50967. [Google Scholar] [CrossRef]
- Bachhawat, B.K.; Coon, M.J.; Kupiecki, F.P.; Nagle, R.; Robinson, W.G. Coenzyme A thiol esters of isobutyric, methacrylic, and beta-hydroxyisobutyric acids as intermediates in the enzymatic degradation of valine. J. Biol. Chem. 1957, 224, 1–11. [Google Scholar]
- Agudelo Garcia, P.A.; Lovejoy, C.M.; Nagarajan, P.; Park, D.; Popova, L.V.; Freitas, M.A.; Parthun, M.R. Histone acetyltransferase 1 is required for DNA replication fork function and stability. J. Biol. Chem. 2020, 295, 8363–8373. [Google Scholar] [CrossRef]
- Yang, X.; Li, L.; Liang, J.; Shi, L.; Yang, J.; Yi, X.; Zhang, D.; Han, X.; Yu, N.; Shang, Y. Histone acetyltransferase 1 promotes homologous recombination in DNA repair by facilitating histone turnover. J. Biol. Chem. 2013, 288, 18271–18282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agudelo Garcia, P.A.; Hoover, M.E.; Zhang, P.; Nagarajan, P.; Freitas, M.A.; Parthun, M.R. Identification of multiple roles for histone acetyltransferase 1 in replication-coupled chromatin assembly. Nucleic Acids Res. 2017, 45, 9319–9335. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, T.; Filippakopoulos, P. Functions of bromodomain-containing proteins and their roles in homeostasis and cancer. Nat. Rev. Mol. Cell Biol. 2017, 18, 246–262. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, P.; Ge, Z.; Sirbu, B.; Doughty, C.; Agudelo Garcia, P.A.; Schlederer, M.; Annunziato, A.T.; Cortez, D.; Kenner, L.; Parthun, M.R. Histone acetyl transferase 1 is essential for mammalian development, genome stability, and the processing of newly synthesized histones H3 and H4. PLoS Genet. 2013, 9, e1003518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, T.L.; Gongol, B.; Zhang, F.; Martin, M.; Johnson, D.A.; Xiao, H.; Wang, Y.; Subramaniam, S.; Chien, S.; Shyy, J.Y. AMPK promotes mitochondrial biogenesis and function by phosphorylating the epigenetic factors DNMT1, RBBP7, and HAT1. Sci. Signal 2017, 10, eaaf7478. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Zhang, M.; Xie, W.; Lan, G.; Cheng, H.P.; Gong, D.; Huang, C.; Lv, Y.C.; Yao, F.; Tan, Y.L.; et al. MiR-486 regulates cholesterol efflux by targeting HAT1. Biochem. Biophys. Res. Commun. 2016, 472, 418–424. [Google Scholar] [CrossRef]
- Sadler, A.J.; Suliman, B.A.; Yu, L.; Yuan, X.; Wang, D.; Irving, A.T.; Sarvestani, S.T.; Banerjee, A.; Mansell, A.S.; Liu, J.P.; et al. The acetyltransferase HAT1 moderates the NF-κB response by regulating the transcription factor PLZF. Nat. Commun. 2015, 6, 6795. [Google Scholar] [CrossRef] [Green Version]
- Vlad, M.L.; Manea, S.A.; Lazar, A.G.; Raicu, M.; Muresian, H.; Simionescu, M.; Manea, A. Histone Acetyltransferase-Dependent Pathways Mediate Upregulation of NADPH Oxidase 5 in Human Macrophages under Inflammatory Conditions: A Potential Mechanism of Reactive Oxygen Species Overproduction in Atherosclerosis. Oxid. Med. Cell Longev. 2019, 2019, 3201062. [Google Scholar] [CrossRef] [Green Version]
- Espíndola, M.S.; Soares, L.S.; Galvão-Lima, L.J.; Zambuzi, F.A.; Cacemiro, M.C.; Brauer, V.S.; Marzocchi-Machado, C.M.; de Souza Gomes, M.; Amaral, L.R.; Martins-Filho, O.A.; et al. Epigenetic alterations are associated with monocyte immune dysfunctions in HIV-1 infection. Sci. Rep. 2018, 8, 5505. [Google Scholar] [CrossRef] [Green Version]
- Seiden-Long, I.M.; Brown, K.R.; Shih, W.; Wigle, D.A.; Radulovich, N.; Jurisica, I.; Tsao, M.S. Transcriptional targets of hepatocyte growth factor signaling and Ki-ras oncogene activation in colorectal cancer. Oncogene 2006, 25, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Tian, S.; Li, P. Histone Acetyltransferase 1 Promotes Cell Proliferation and Induces Cisplatin Resistance in Hepatocellular Carcinoma. Oncol. Res. 2017, 25, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Hou, J.; Wang, Q.; Yao, L.; Xu, S.; Ge, D. RNAi screening identifies HAT1 as a potential drug target in esophageal squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 3898–3907. [Google Scholar] [PubMed]
- Lin, C.Y.; Chao, A.; Wu, R.C.; Lee, L.Y.; Ueng, S.H.; Tsai, C.L.; Lee, Y.S.; Peng, M.T.; Yang, L.Y.; Huang, H.J.; et al. Synergistic effects of pazopanib and hyperthermia against uterine leiomyosarcoma growth mediated by downregulation of histone acetyltransferase 1. J. Mol. Med. 2020, 98, 1175–1188. [Google Scholar] [CrossRef] [PubMed]
- Miao, B.P.; Zhang, R.S.; Yang, G.; Sun, J.J.; Tang, Y.Y.; Liang, W.F.; Liu, T.; Wen, Z.; Yang, P.C.; Nie, G.H. Histone acetyltransferase 1 up regulates Bcl2L12 expression in nasopharyngeal cancer cells. Arch. Biochem. Biophys. 2018, 646, 72–79. [Google Scholar] [CrossRef]
- Fan, P.; Zhao, J.; Meng, Z.; Wu, H.; Wang, B.; Wu, H.; Jin, X. Overexpressed histone acetyltransferase 1 regulates cancer immunity by increasing programmed death-ligand 1 expression in pancreatic cancer. J. Exp. Clin. Cancer Res. 2019, 38, 47. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Li, X.; Xu, Y.; Guo, W.; Yu, H.; Zhang, L.; Wang, Y.; Chen, X. Acetylation-stabilized chloride intracellular channel 1 exerts a tumor-promoting effect on cervical cancer cells by activating NF-κB. Cell. Oncol. 2021, 44, 557–568. [Google Scholar] [CrossRef]
- Chrun, E.S.; Modolo, F.; Vieira, D.; Borges-Júnior, Á.; Castro, R.G.; Daniel, F.I. Immunoexpression of HDAC1, HDAC2, and HAT1 in actinic cheilitis and lip squamous cell carcinoma. Oral. Dis. 2017, 23, 505–510. [Google Scholar] [CrossRef]
- Sarkar, T.; Dhar, S.; Chakraborty, D.; Pati, S.; Bose, S.; Panda, A.K.; Basak, U.; Chakraborty, S.; Mukherjee, S.; Guin, A.; et al. FOXP3/HAT1 Axis Controls Treg Infiltration in the Tumor Microenvironment by Inducing CCR4 Expression in Breast Cancer. Front. Immunol. 2022, 13, 740588. [Google Scholar] [CrossRef] [PubMed]
- Min, S.K.; Koh, Y.H.; Park, Y.; Kim, H.J.; Seo, J.; Park, H.R.; Cho, S.J.; Kim, I.S. Expression of HAT1 and HDAC1, 2, 3 in Diffuse Large B-Cell Lymphomas, Peripheral T-Cell Lymphomas, and NK/T-Cell Lymphomas. Korean J. Pathol. 2012, 46, 142–150. [Google Scholar] [CrossRef]
- Han, N.; Shi, L.; Guo, Q.; Sun, W.; Yu, Y.; Yang, L.; Zhang, X.; Zhang, M. HAT1 induces lung cancer cell apoptosis via up regulating Fas. Oncotarget 2017, 8, 89970–89977. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, C.; Li, Q.; Chang, H.C.; Tang, Y.C. SIRT7 Facilitates CENP-A Nucleosome Assembly and Suppresses Intestinal Tumorigenesis. iScience 2020, 23, 101461. [Google Scholar] [CrossRef] [PubMed]
- Bugide, S.; Parajuli, K.R.; Chava, S.; Pattanayak, R.; Manna, D.L.D.; Shrestha, D.; Yang, E.S.; Cai, G.; Johnson, D.B.; Gupta, R. Loss of HAT1 expression confers BRAFV600E inhibitor resistance to melanoma cells by activating MAPK signaling via IGF1R. Oncogenesis 2020, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ren, D.; Zhou, Y.; Shen, J.; Wu, H.; Jin, X. Histone acetyltransferase 1 promotes gemcitabine resistance by regulating the PVT1/EZH2 complex in pancreatic cancer. Cell Death Dis. 2021, 12, 878. [Google Scholar] [CrossRef]
- You, L.; Chang, D.; Du, H.Z.; Zhao, Y.P. Genome-wide screen identifies PVT1 as a regulator of Gemcitabine sensitivity in human pancreatic cancer cells. Biochem. Biophys. Res. Commun. 2011, 407, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Hao, C.; Wang, C.; Li, L. Long noncoding RNA PVT1 modulates hepatocellular carcinoma cell proliferation and apoptosis by recruiting EZH2. Cancer Cell Int. 2018, 18, 98. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Chen, J.; Feng, J.; Wang, J. Long noncoding RNA PVT1 modulates thyroid cancer cell proliferation by recruiting EZH2 and regulating thyroid-stimulating hormone receptor (TSHR). Tumour Biol. 2016, 37, 3105–3113. [Google Scholar] [CrossRef]
- Hong, Z.; Xiang, Z.; Zhang, P.; Wu, Q.; Xu, C.; Wang, X.; Shi, G.; Hong, Z.; Wu, D. Histone acetyltransferase 1 upregulates androgen receptor expression to modulate CRPC cell resistance to enzalutamide. Clin. Transl. Med. 2021, 11, e495. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | HAT1 Expression | Outcomes | References |
|---|---|---|---|
| Pancreatic cancer (PC) | High | Resistance to chemotherapy and radiotherapy Poor prognosis | [31] |
| Colorectal cancer (CRC) | High Low | Not reported | [43,54] |
| Hepatocellular carcinoma (HCC) | High | Resistance to chemotherapy | [44] |
| Esophageal squamous cancer (ESC) | High | Poor tumor differentiation | [45] |
| Uterine leiomyosarcoma (LMS) | High | Poor clinical prognosis Resistant to chemotherapy and radiotherapy | [46] |
| Nasopharyngeal carcinoma (NPC) | High | Not reported | [47] |
| Cervical cancer (CC) | High | Poor prognosis | [49] |
| Lip squamous cell carcinoma (LSCC) | High | Represents 2.1% of all cancers | [50] |
| Actinic cheilitis (AC) | High | Represents 2.1% of all cancers | [50] |
| Breast cancer (BC) | High | Poor prognosis | [51] |
| Diffuse large B-cell lymphoma (DLBCL) | High | Tumor invasion and metastasis Poor prognosis | [52] |
| Peripheral T-cell lymphoma (PTCL) | High | Tumor invasion and metastasis Poor prognosis | [52] |
| Lung cancer cell (LCC) | Low | Five-year survival rate is less than 20% | [53] |
| Melanoma | Low | Resistance to chemotherapy | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capone, V.; Della Torre, L.; Carannante, D.; Babaei, M.; Altucci, L.; Benedetti, R.; Carafa, V. HAT1: Landscape of Biological Function and Role in Cancer. Cells 2023, 12, 1075. https://doi.org/10.3390/cells12071075
Capone V, Della Torre L, Carannante D, Babaei M, Altucci L, Benedetti R, Carafa V. HAT1: Landscape of Biological Function and Role in Cancer. Cells. 2023; 12(7):1075. https://doi.org/10.3390/cells12071075
Chicago/Turabian StyleCapone, Vincenza, Laura Della Torre, Daniela Carannante, Mehrad Babaei, Lucia Altucci, Rosaria Benedetti, and Vincenzo Carafa. 2023. "HAT1: Landscape of Biological Function and Role in Cancer" Cells 12, no. 7: 1075. https://doi.org/10.3390/cells12071075
APA StyleCapone, V., Della Torre, L., Carannante, D., Babaei, M., Altucci, L., Benedetti, R., & Carafa, V. (2023). HAT1: Landscape of Biological Function and Role in Cancer. Cells, 12(7), 1075. https://doi.org/10.3390/cells12071075