


New Findings: Hindlimb Unloading Causes Nucleocytoplasmic Ca2+ Overload and DNA Damage in Skeletal Muscle

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hindlimb Unloading Animal Modeling and Sample Preparation
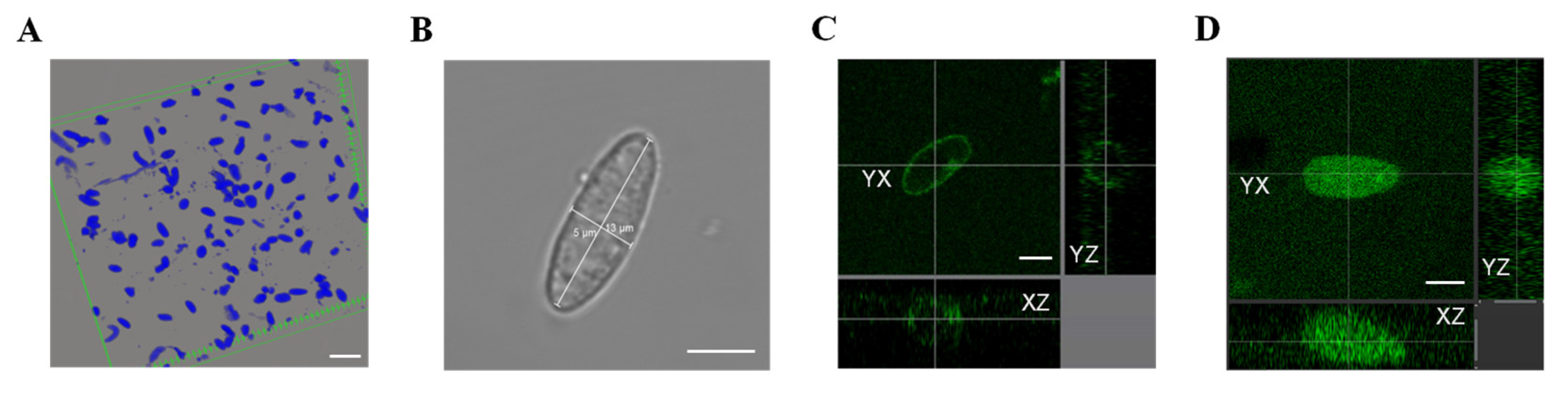
2.2. Nuclei Isolation and [Ca2+]NE and [Ca2+]NC Measurement
2.3. Western Blot Assay (WB)
2.3.1. Nuclear Protein Extraction
2.3.2. Western Blot
2.4. IP3 and IP4 Content Measurement
2.5. TUNEL Staining for Nucleus and Immunofluorescence Staining for DNase X
2.6. DNA Fragmentation Assay
2.7. Data Statistics
3. Results
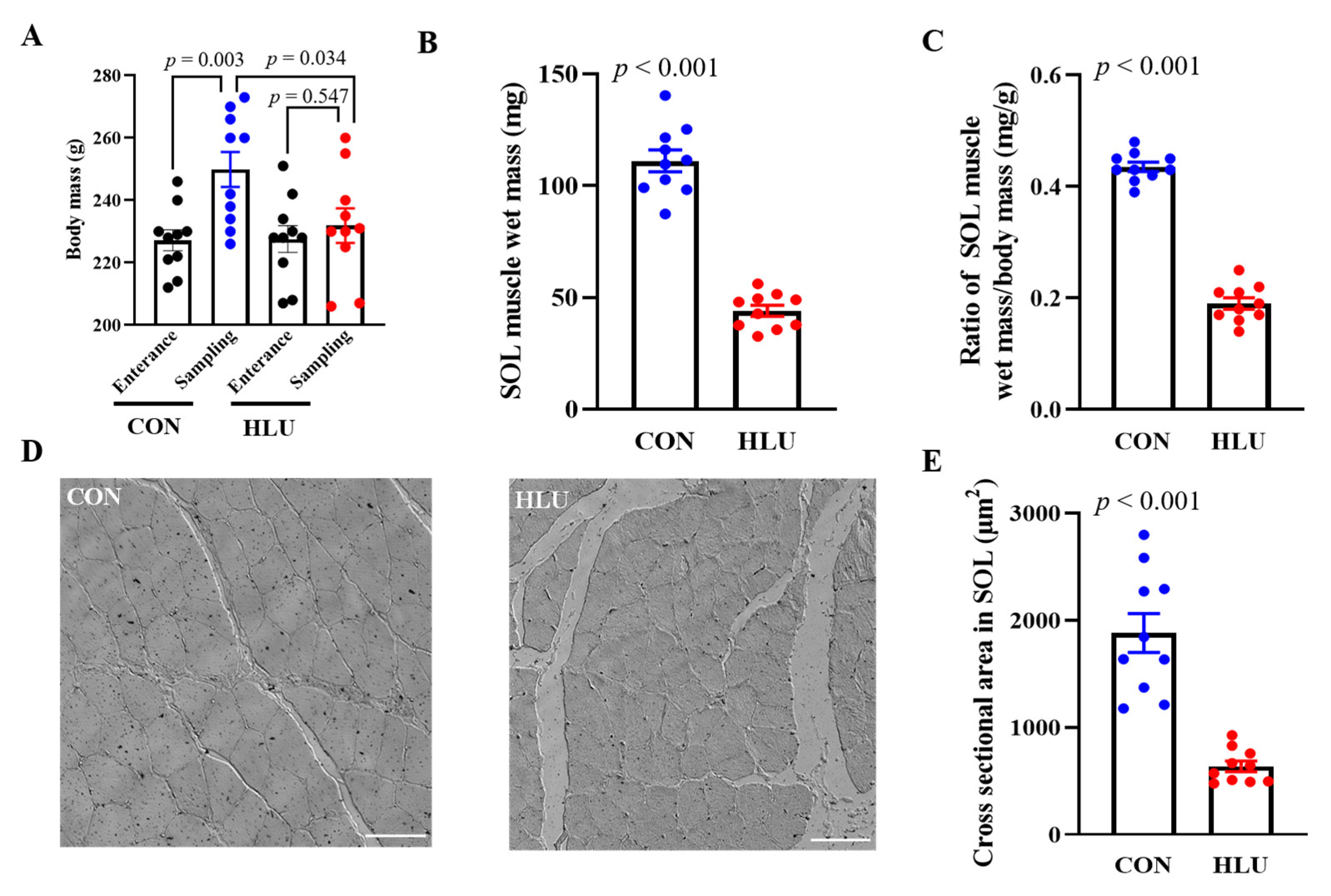
3.1. The Changes of Body Mass and Muscle Morphology
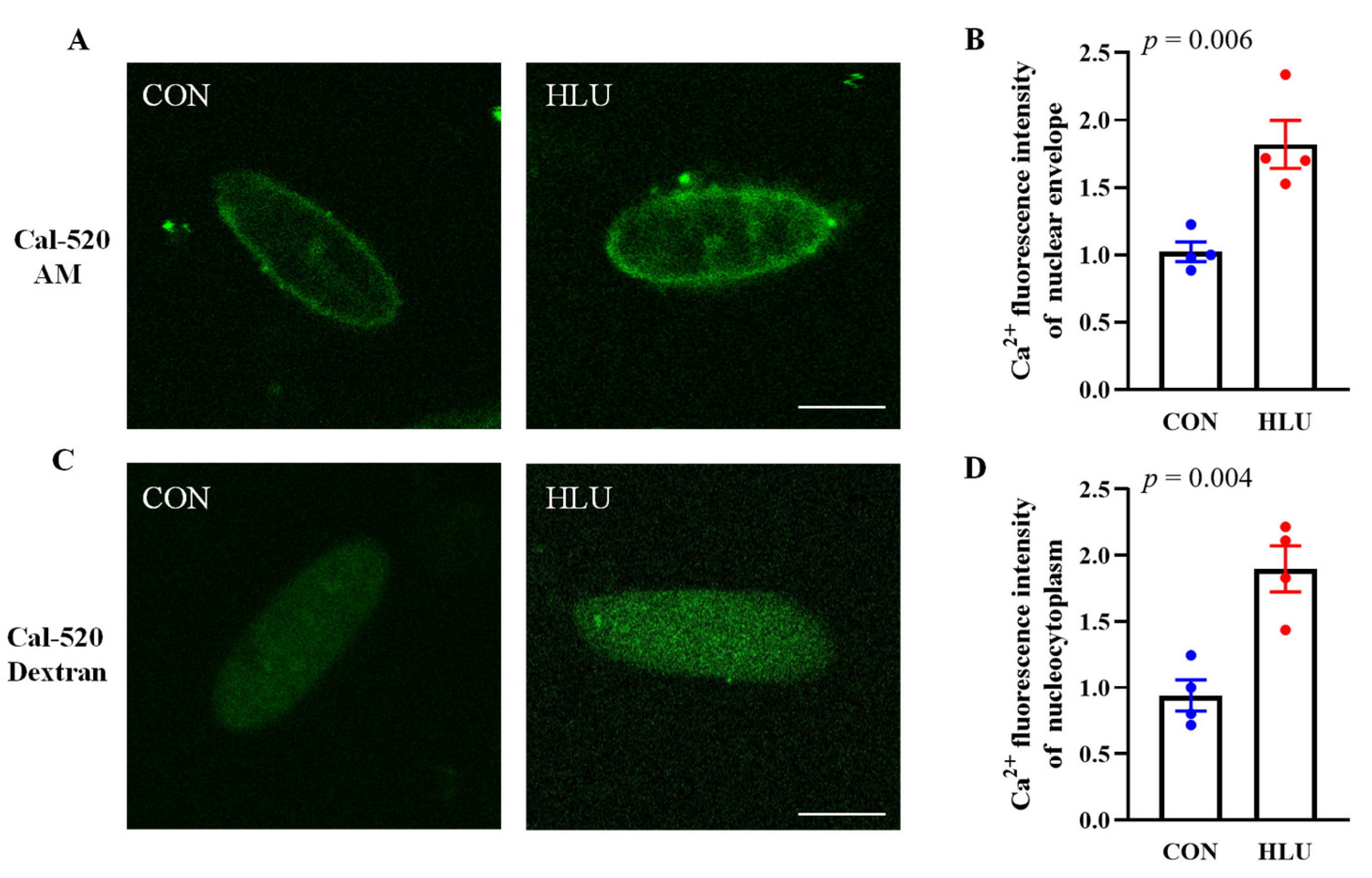
3.2. Changes of [Ca2+]NE and [Ca2+]NC
3.3. Relative Expression level of Ca2+ Transporters Located on Nuclear Membrane and Its Regulatory Proteins
3.4. The IP3 and IP4 Content in Muscle
3.5. Apoptosis of Myonucleus and Involvement of DNase X
3.6. The DNase X Activity
4. Discussion
4.1. Hindlimb Unloading Leads to [Ca2+]NE and [Ca2+]NC Elevating in SOL Muscle
4.2. Changes in Ca2+ Transport Capacity by INM and ONM may Explain why Ca2+ Levels Increase in Nuclear Envelope and Nucleocytoplasm during Hindlimb Unloading
4.2.1. Ca2+ Uptake from the Cytoplasm on ONM Increases in Disused SOL Muscle
4.2.2. Ca2+ Release Increases while Ca2+ Uptake Attenuates on INM in Disused Muscle
4.3. Nucleocytoplasmic Ca2+ Overload Leads to Myonuclear Apoptosis by Increasing DNase X Activity during Hindlimb Unloading
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| [Ca2+]NC | Ca2+ level in nucleocytoplasm |
| [Ca2+]NE | Ca2+ level in nuclear envelope |
| cADPR | Cyclic ADP-ribose |
| CaMK II | Ca2+/calmodulin dependent protein kinase II |
| CAX | Calnexin |
| CD38 | Cyclic ADP ribose hydrolase |
| CON | Cage control |
| CSA | Cross sectional area |
| CSQ1 | Calsequestrin 1 |
| DAG | Diacylglycerol |
| HLU | Hindlimb unloading |
| INM | Inner nuclear membrane |
| IP3 | Inositol 1,4,5-tetrakisphosphate |
| IP3K | Inositol 1,4,5-tetrakisphosphate Kinase |
| IP3R | Inositol 1,4,5-tetrakisphosphate receptor |
| IP4 | Inositol 1,3,4,5-tetrakisphosphate |
| IP4R | Inositol 1,3,4,5-tetrakisphosphate receptor |
| LaminB1 | Lamina protein B1 |
| NC | Nucleocytoplasm |
| NCX3 | Na+/Ca2+ exchanger 3 |
| NE | Nuclear envelope |
| NPC | Nuclear pore complex |
| ONM | Outer nuclear membrane |
| PCM1 | Pericentriolar material 1 |
| PIP2 | Phosphatidylinositol (4,5) bisphosphate |
| PKA | Protein kinase A |
| PLC | Phospholipase C |
| PM | Plasma membrane |
| RyR1 | Ryanodine receptor 1 |
| SOL | Soleus |
References
- Gao, Y.; Arfat, Y.; Wang, H.; Goswami, N. Muscle Atrophy Induced by Mechanical Unloading: Mechanisms and Potential Countermeasures. Front. Physiol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharlo, K.; Tyganov, S.A.; Tomilovskaya, E.; Popov, D.V.; Saveko, A.A.; Shenkman, B.S. Effects of Various Muscle Disuse States and Countermeasures on Muscle Molecular Signaling. Int. J. Mol. Sci. 2021, 23, 468. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Gao, Y.F.; Zhao, X.H.; Cui, J.H. Effects of tetramethylpyrazine on nitric oxide synthase activity and calcium ion concentration of skeletal muscle in hindlimb unloading rats. Zhonghua Yi Xue Za Zhi 2012, 92, 2075–2077. [Google Scholar] [PubMed]
- Ingalls, C.P.; Wenke, J.C.; Armstrong, R.B. Time course changes in [Ca2+]i, force, and protein content in hindlimb-suspended mouse soleus muscles. Aviat. Space Environ. Med. 2001, 72, 471–476. [Google Scholar]
- Tomiya, S.; Tamura, Y.; Kouzaki, K.; Kotani, T.; Wakabayashi, Y.; Noda, M.; Nakazato, K. Cast immobilization of hindlimb upregulates sarcolipin expression in atrophied skeletal muscles and increases thermogenesis in C57BL/6J mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, 649–661. [Google Scholar] [CrossRef]
- Echevarria, W.; Leite, M.F.; Guerra, M.T.; Zipfel, W.R.; Nathanson, M.H. Regulation of calcium signals in the nucleus by a nucleoplasmic reticulum. Nat. Cell Biol. 2003, 5, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A.; Ludwig, Y.; Shahin, V.; Oberleithner, H. A pathway separate from the central channel through the nuclear pore complex for inorganic ions and small macromolecules. J. Biol. Chem. 2007, 282, 31437–31443. [Google Scholar] [CrossRef] [Green Version]
- Secondo, A.; Petrozziello, T.; Tedeschi, V.; Boscia, F.; Pannaccione, A.; Molinaro, P.; Annunziato, L. Nuclear localization of NCX: Role in Ca2+ handling and pathophysiological implications. Cell Calcium 2020, 86, 102143. [Google Scholar] [CrossRef]
- Leite, M.F.; Thrower, E.C.; Echevarria, W.; Koulen, P.; Hirata, K.; Bennett, A.M.; Ehrlich, B.E.; Nathanson, M.H. Nuclear and cytosolic calcium are regulated independently. Proc. Natl. Acad. Sci. USA 2003, 100, 2975–2980. [Google Scholar] [CrossRef] [Green Version]
- Karin, M. Signal transduction from cell surface to nucleus in development and disease. FASEB J. 1992, 6, 2581–2590. [Google Scholar] [CrossRef]
- Peunova, N.; Enikolopov, G. Amplification of calcium-induced gene transcription by nitric oxide in neuronal cells. Nature 1993, 364, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, J.S.; Czubryt, M.P.; Pierce, G.N. Calcium and calcium-binding proteins in the nucleus. Mol. Cell Biochem. 1994, 135, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Bachs, O.; Agell, N.; Carafoli, E. Calmodulin and calmodulin-binding proteins in the nucleus. Cell Calcium 1994, 16, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Steinhardt, R.A.; Alderton, J. Intracellular free calcium rise triggers nuclear envelope breakdown in the sea urchin embryo. Nature 1988, 332, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Nicotera, P.; Rossi, A.D. Nuclear Ca2+: Physiological regulation and role in apoptosis. Mol. Cell Biochem. 1994, 135, 89–98. [Google Scholar] [CrossRef]
- Los, M.; Neubüser, D.; Coy, J.F.; Mozoluk, M.; Poustka, A.; Schulze-Osthoff, K. Functional characterization of DNase X, a novel endonuclease expressed in muscle cells. Biochemistry 2000, 39, 7365–7373. [Google Scholar] [CrossRef] [Green Version]
- Hu, N.F.; Chang, H.; Du, B.; Zhang, Q.W.; Arfat, Y.; Dang, K.; Gao, Y.F. Tetramethylpyrazine ameliorated disuse-induced gastrocnemius muscle atrophy in hindlimb unloading rats through suppression of Ca2+/ROS-mediated apoptosis. Appl. Physiol. Nutr. Metab. 2017, 42, 117–127. [Google Scholar] [CrossRef]
- Smith, H.K.; Maxwell, L.; Martyn, J.A.; Bass, J.J. Nuclear DNA fragmentation and morphological alterations in adult rabbit skeletal muscle after short-term immobilization. Cell Tissue Res. 2000, 302, 235–241. [Google Scholar] [CrossRef]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef]
- Li, B.; Jie, W.; Huang, L.; Wei, P.; Li, S.; Luo, Z.; Friedman, A.K.; Meredith, A.L.; Han, M.H.; Zhu, X.H.; et al. Nuclear BK channels regulate gene expression via the control of nuclear calcium signaling. Nat. Neurosci. 2014, 17, 1055–1063. [Google Scholar] [CrossRef] [Green Version]
- Wilkie, G.S.; Schirmer, E.C. Purification of Nuclei and Preparation of Nuclear Envelopes from Skeletal Muscle. In The Nucleus: Volume 1: Nuclei and Subnuclear Components; Hancock, R., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 23–41. [Google Scholar]
- Shiokawa, D.; Tanuma, S. Characterization of human DNase I family endonucleases and activation of DNase gamma during apoptosis. Biochemistry 2001, 40, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Schmalbruch, H.; Hellhammer, U. The number of nuclei in adult rat muscles with special reference to satellite cells. Anat. Rec. 1977, 189, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Yarotskyy, V.; Dirksen, R.T. Monovalent cationic channel activity in the inner membrane of nuclei from skeletal muscle fibers. Biophys. J. 2014, 107, 2027–2036. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, C.; Liberona, J.L.; Molgó, J.; Colasante, C.; Mignery, G.A.; Jaimovich, E. Nuclear inositol 1,4,5-trisphosphate receptors regulate local Ca2+ transients and modulate cAMP response element binding protein phosphorylation. J. Cell Sci. 2005, 118, 3131–3140. [Google Scholar] [CrossRef] [Green Version]
- Ni, M.; Lee, A.S. ER chaperones in mammalian development and human diseases. FEBS Lett. 2007, 581, 3641–3651. [Google Scholar] [CrossRef] [Green Version]
- Winje, I.M.; Bengtsen, M.; Eftestøl, E.; Juvkam, I.; Bruusgaard, J.C.; Gundersen, K. Specific labelling of myonuclei by an antibody against pericentriolar material 1 on skeletal muscle tissue sections. Acta Physiol. 2018, 223, e13034. [Google Scholar] [CrossRef] [Green Version]
- Shiokawa, D.; Tanuma, S. Molecular cloning and expression of a cDNA encoding an apoptotic endonuclease DNase gamma. Biochem. J. 1998, 332, 713–720. [Google Scholar] [CrossRef]
- Gerasimenko, J.; Maruyama, Y.; Tepikin, A.; Petersen, O.H.; Gerasimenko, O. Calcium signalling in and around the nuclear envelope. Biochem. Soc. Trans. 2003, 31, 76–78. [Google Scholar] [CrossRef]
- Gerasimenko, O.V.; Gerasimenko, J.V.; Tepikin, A.V.; Petersen, O.H. ATP-dependent accumulation and inositol trisphosphate- or cyclic ADP-ribose-mediated release of Ca2+ from the nuclear envelope. Cell 1995, 80, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Quesada, I.; Rovira, J.M.; Martin, F.; Roche, E.; Nadal, A.; Soria, B. Nuclear KATP channels trigger nuclear Ca2+ transients that modulate nuclear function. Proc. Natl. Acad. Sci. USA 2002, 99, 9544–9549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qualls, A.E.; Southern, W.M.; Call, J.A. Mitochondria-cytokine crosstalk following skeletal muscle injury and disuse: A mini-review. Am. J. Physiol. Cell Physiol. 2021, 320, C681–C688. [Google Scholar] [CrossRef]
- Lanini, L.; Bachs, O.; Carafoli, E. The calcium pump of the liver nuclear membrane is identical to that of endoplasmic reticulum. J. Biol. Chem. 1992, 267, 11548–11552. [Google Scholar] [CrossRef]
- Periasamy, M.; Kalyanasundaram, A. SERCA pump isoforms: Their role in calcium transport and disease. Muscle Nerve 2007, 35, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Quist, E.E.; Foresman, B.H.; Vasan, R.; Quist, C.W. Inositol tetrakisphosphate stimulates a novel ATP-independent Ca2+ uptake mechanism in cardiac junctional sarcoplasmic reticulum. Biochem. Biophys. Res. Commun. 1994, 204, 69–75. [Google Scholar] [CrossRef]
- Ogawa, Y. Role of ryanodine receptors. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 229–274. [Google Scholar] [CrossRef] [PubMed]
- Adebanjo, O.A.; Anandatheerthavarada, H.K.; Koval, A.P.; Moonga, B.S.; Biswas, G.; Sun, L.; Sodam, B.R.; Bevis, P.J.; Huang, C.L.; Epstein, S.; et al. A new function for CD38/ADP-ribosyl cyclase in nuclear Ca2+ homeostasis. Nat. Cell Biol. 1999, 1, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Kraner, S.D.; Wang, Q.; Novak, K.R.; Cheng, D.; Cool, D.R.; Peng, J.; Rich, M.M. Upregulation of the CaV 1.1-ryanodine receptor complex in a rat model of critical illness myopathy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1384–R1391. [Google Scholar] [CrossRef] [Green Version]
- Adebanjo, O.A.; Biswas, G.; Moonga, B.S.; Anandatheerthavarada, H.K.; Sun, L.; Bevis, P.J.; Sodam, B.R.; Lai, F.A.; Avadhani, N.G.; Zaidi, M. Novel biochemical and functional insights into nuclear Ca2+ transport through IP(3)Rs and RyRs in osteoblasts. Am. J. Physiol. Ren. Physiol. 2000, 278, F784–F791. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.Y.; Vahdati Hassani, F.; Hoffmann, D.; Xiao, J.; Xiong, F.; Villeneuve, L.R.; Ljubojevic-Holzer, S.; Kamler, M.; Abu-Taha, I.; Heijman, J.; et al. Inositol Trisphosphate Receptors and Nuclear Calcium in Atrial Fibrillation. Circ. Res. 2021, 128, 619–635. [Google Scholar] [CrossRef]
- Lanner, J.T.; Georgiou, D.K.; Joshi, A.D.; Hamilton, S.L. Ryanodine receptors: Structure, expression, molecular details, and function in calcium release. Cold Spring Harb. Perspect. Biol. 2010, 2, a003996. [Google Scholar] [CrossRef] [Green Version]
- Wagner, L.E., 2nd; Joseph, S.K.; Yule, D.I. Regulation of single inositol 1,4,5-trisphosphate receptor channel activity by protein kinase A phosphorylation. J. Physiol. 2008, 586, 3577–3596. [Google Scholar] [CrossRef] [PubMed]
- Ledeen, R.W.; Wu, G. Sodium-calcium exchangers in the nucleus: An unexpected locus and an unusual regulatory mechanism. Ann. N. Y. Acad. Sci. 2007, 1099, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lu, Z.H.; Xie, X.; Ledeen, R.W. Susceptibility of cerebellar granule neurons from GM2/GD2 synthase-null mice to apoptosis induced by glutamate excitotoxicity and elevated KCl: Rescue by GM1 and LIGA20. Glycoconj. J. 2004, 21, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, D.A.; Quednau, B.D.; Qui, Z.; Xia, Y.R.; Lusis, A.J.; Philipson, K.D. Cloning of a third mammalian Na+-Ca2+ exchanger, NCX3. J. Biol. Chem. 1996, 271, 24914–24921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bootman, M.D.; Fearnley, C.; Smyrnias, I.; MacDonald, F.; Roderick, H.L. An update on nuclear calcium signalling. J. Cell Sci. 2009, 122, 2337–2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukube, T.; McCully, J.D.; Faulk, E.A.; Federman, M.; LoCicero, J., 3rd; Krukenkamp, I.B.; Levitsky, S. Magnesium cardioplegia reduces cytosolic and nuclear calcium and DNA fragmentation in the senescent myocardium. Ann. Thorac. Surg. 1994, 58, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Tanuma, S.; Shiokawa, D. Multiple forms of nuclear deoxyribonuclease in rat thymocytes. Biochem. Biophys. Res. Commun. 1994, 203, 789–797. [Google Scholar] [CrossRef] [PubMed]
- McConkey, D.J.; Hartzell, P.; Nicotera, P.; Orrenius, S. Calcium-activated DNA fragmentation kills immature thymocytes. FASEB J. 1989, 3, 1843–1849. [Google Scholar] [CrossRef] [Green Version]
- Allen, D.L.; Linderman, J.K.; Roy, R.R.; Bigbee, A.J.; Grindeland, R.E.; Mukku, V.; Edgerton, V.R. Apoptosis: A mechanism contributing to remodeling of skeletal muscle in response to hindlimb unweighting. Am. J. Physiol. 1997, 273, C579–C587. [Google Scholar] [CrossRef]
- Leeuwenburgh, C.; Gurley, C.M.; Strotman, B.A.; Dupont-Versteegden, E.E. Age-related differences in apoptosis with disuse atrophy in soleus muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1288–R1296. [Google Scholar] [CrossRef] [Green Version]
- Duddy, W.J.; Cohen, T.; Duguez, S.; Partridge, T.A. The isolated muscle fibre as a model of disuse atrophy: Characterization using PhAct, a method to quantify f-actin. Exp. Cell Res. 2011, 317, 1979–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruusgaard, J.C.; Johansen, I.B.; Egner, I.M.; Rana, Z.A.; Gundersen, K. Myonuclei acquired by overload exercise precede hypertrophy and are not lost on detraining. Proc. Natl. Acad. Sci. USA 2010, 107, 15111–15116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmati, M.; McCarthy, J.J.; Malakoutinia, F. Myonuclear permanence in skeletal muscle memory: A systematic review and meta-analysis of human and animal studies. J. Cachexia Sarcopenia Muscle 2022, 13, 2276–2297. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Antibody Details |
|---|---|
| Ca2+-ATPase type 2 (Ca2+-ATPase2) | 1:1000, 4388S, Cell Signaling Technology, Danvers, MA, USA |
| Ca2+/calmodulin dependent protein kinase (CaMK Ⅱ) | 1:1000, DF2907, Affinity Biosciences, Cincinnati, OH, USA |
| Calnexin (CAX) | 1:1000, YT0613, Immunoway, Plano, TX, USA |
| Cyclic ADP ribose hydrolase (CD38) | 1:1000, YT5392, Immunoway, Plano, TX, USA |
| Calsequestrin 1 (CSQ1) | 1:1000, ab191564, Abcam, Cambridge, UK |
| Inositol 1,4,5-tetrakisphosphate receptor (IP3R1) | 1:1000, ab108517, Abcam, Cambridge, UK |
| Lamina protein B1 (LaminB1) | 1:1000, 12586S, Cell Signaling Technology, Danvers, MA, USA |
| Na+/Ca2+ exchanger 3 (NCX3) | 1:1000, YN1335, Immunoway, Plano, TX, USA |
| Protein kinase A (PKA) | 1:1000, AF5450, Affinity Biosciences, Cincinnati, OH, USA |
| Ryanodine receptor 1 (RyR1) | 1:1000, 8153S, Cell Signaling Technology, Danvers, MA, USA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Wang, H.; Pan, F.; Guo, Y.; Cao, L.; Yan, W.; Gao, Y. New Findings: Hindlimb Unloading Causes Nucleocytoplasmic Ca2+ Overload and DNA Damage in Skeletal Muscle. Cells 2023, 12, 1077. https://doi.org/10.3390/cells12071077
Yang H, Wang H, Pan F, Guo Y, Cao L, Yan W, Gao Y. New Findings: Hindlimb Unloading Causes Nucleocytoplasmic Ca2+ Overload and DNA Damage in Skeletal Muscle. Cells. 2023; 12(7):1077. https://doi.org/10.3390/cells12071077
Chicago/Turabian StyleYang, Huajian, Huiping Wang, Fangyang Pan, Yuxi Guo, Liqi Cao, Wenjing Yan, and Yunfang Gao. 2023. "New Findings: Hindlimb Unloading Causes Nucleocytoplasmic Ca2+ Overload and DNA Damage in Skeletal Muscle" Cells 12, no. 7: 1077. https://doi.org/10.3390/cells12071077
APA StyleYang, H., Wang, H., Pan, F., Guo, Y., Cao, L., Yan, W., & Gao, Y. (2023). New Findings: Hindlimb Unloading Causes Nucleocytoplasmic Ca2+ Overload and DNA Damage in Skeletal Muscle. Cells, 12(7), 1077. https://doi.org/10.3390/cells12071077