

Retinal Development in a Precocial Bird Species, the Quail (Coturnix coturnix, Linnaeus 1758)

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Processing
2.2. Immunohistochemistry
2.3. Image Acquisition and Processing
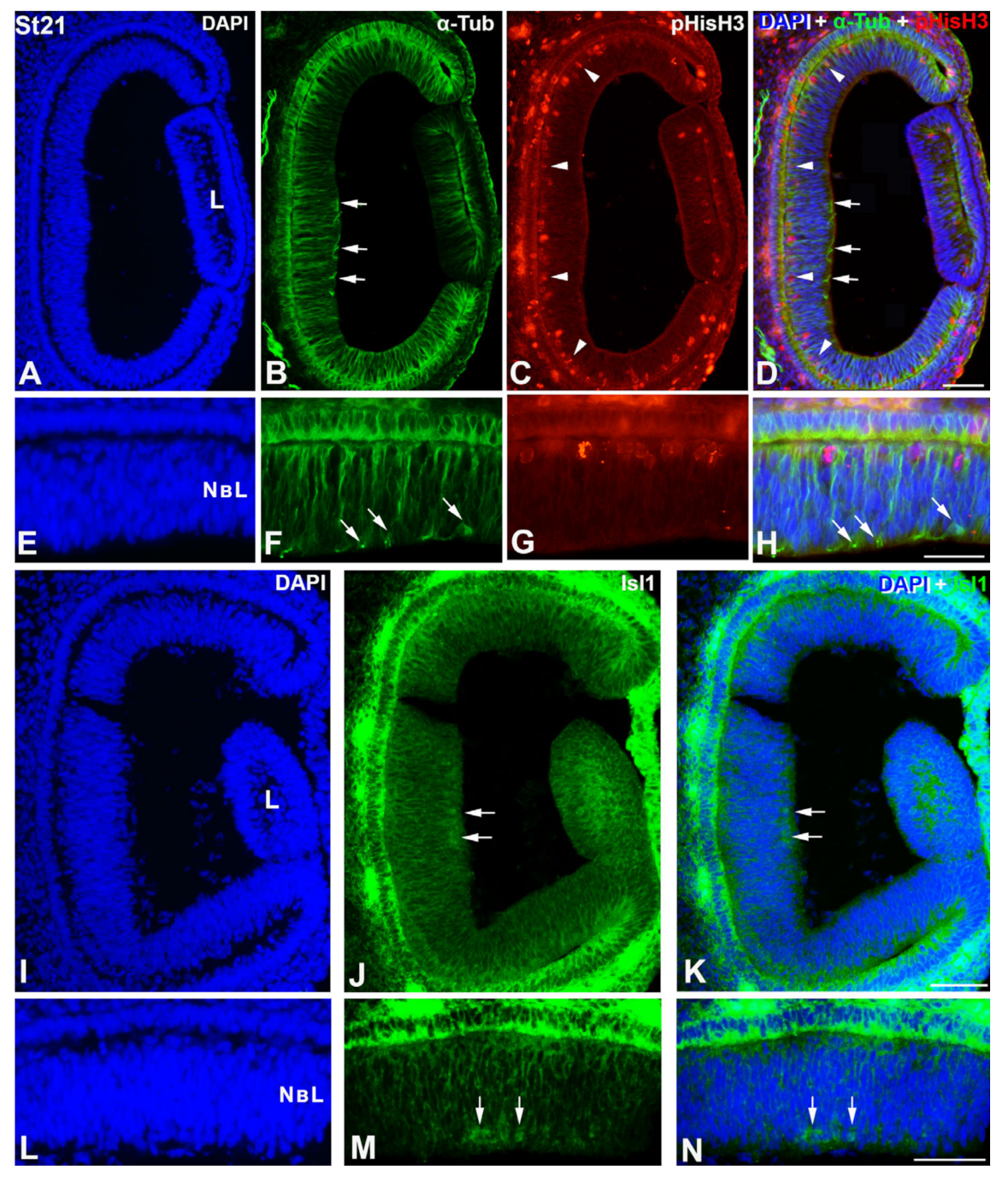
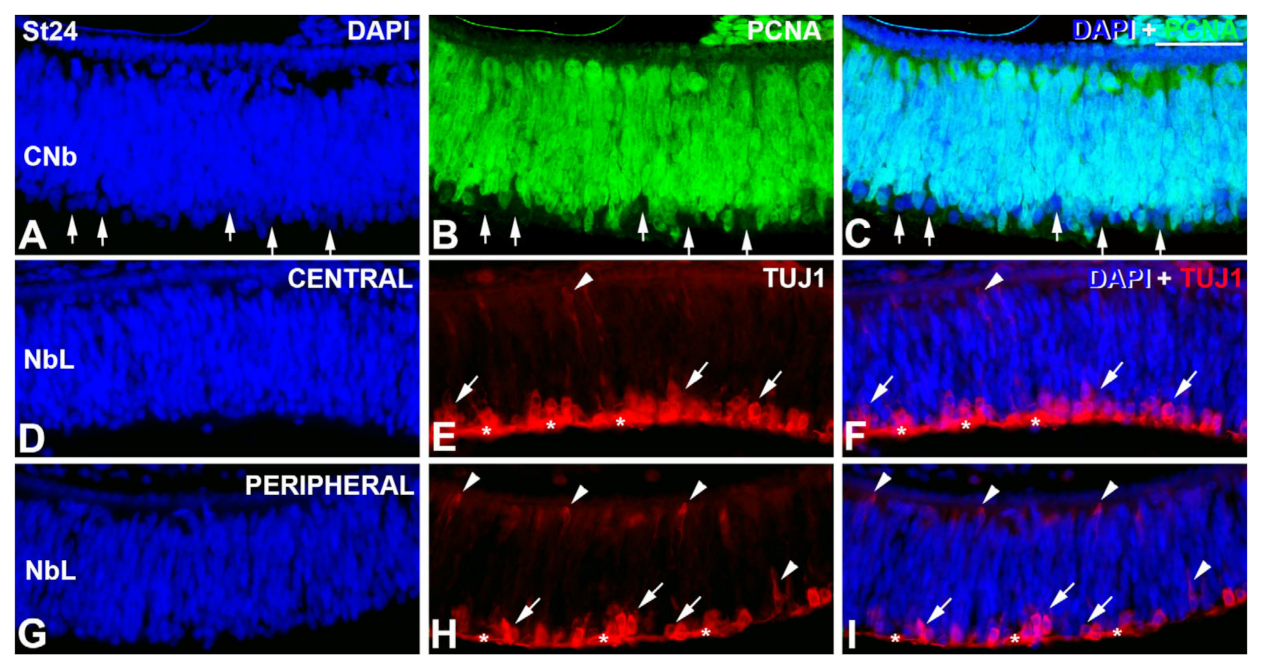
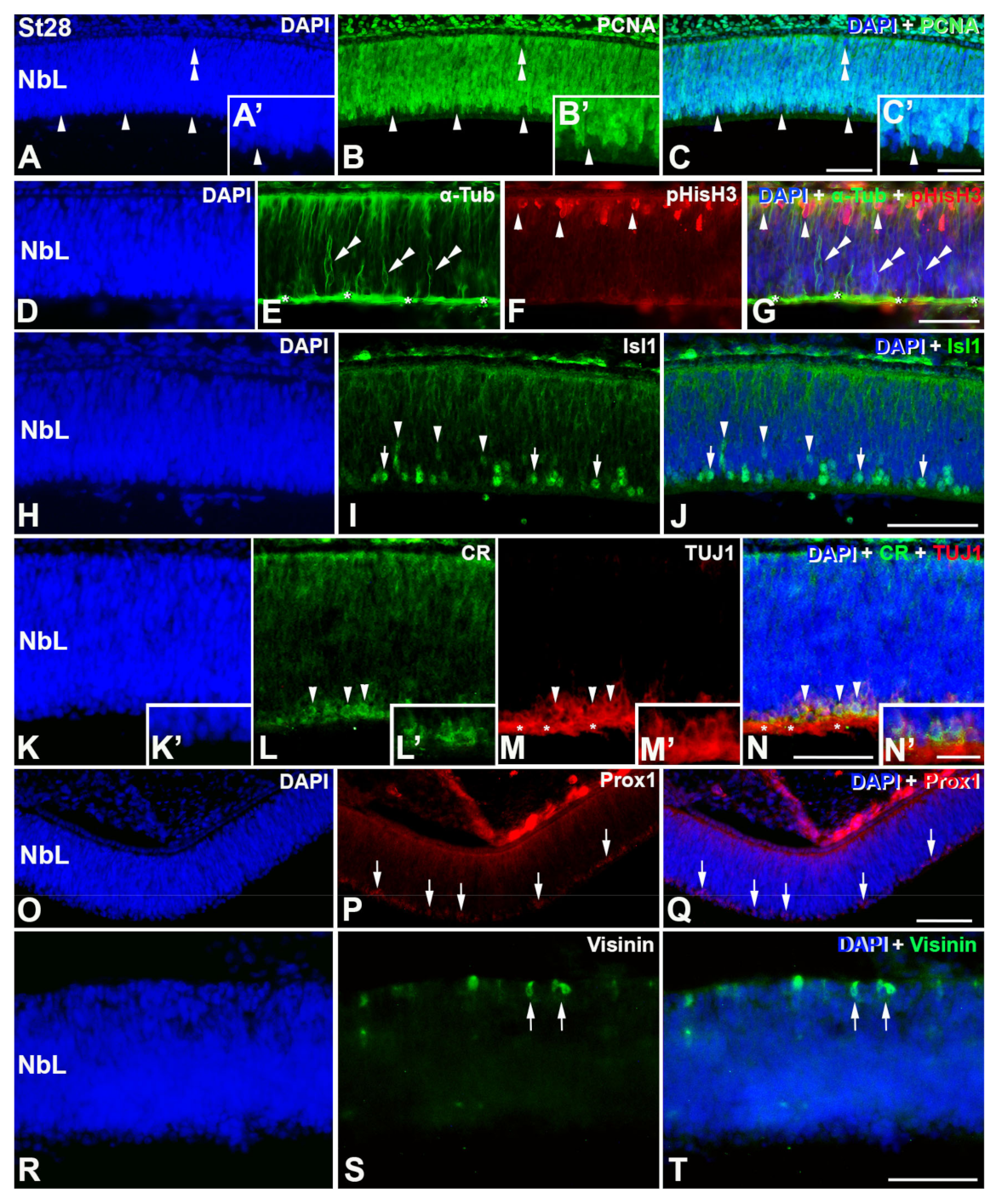
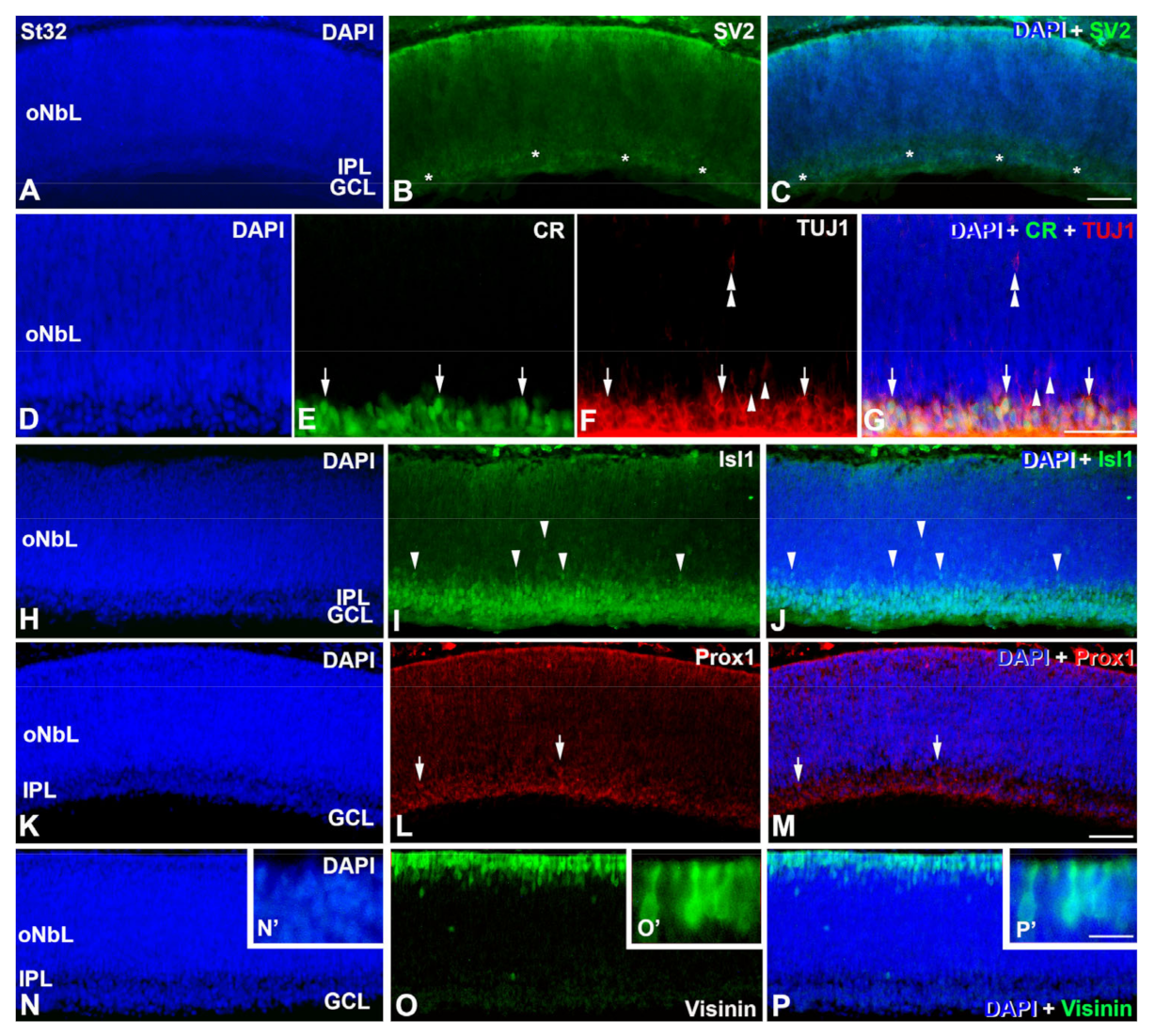
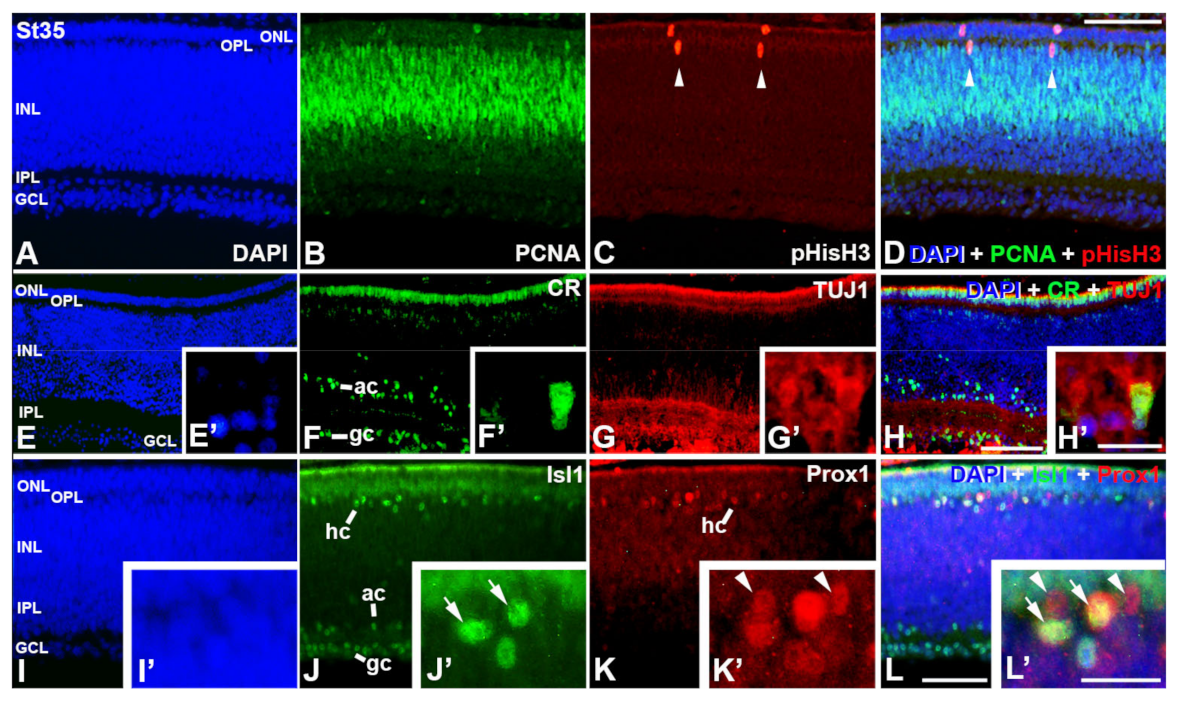
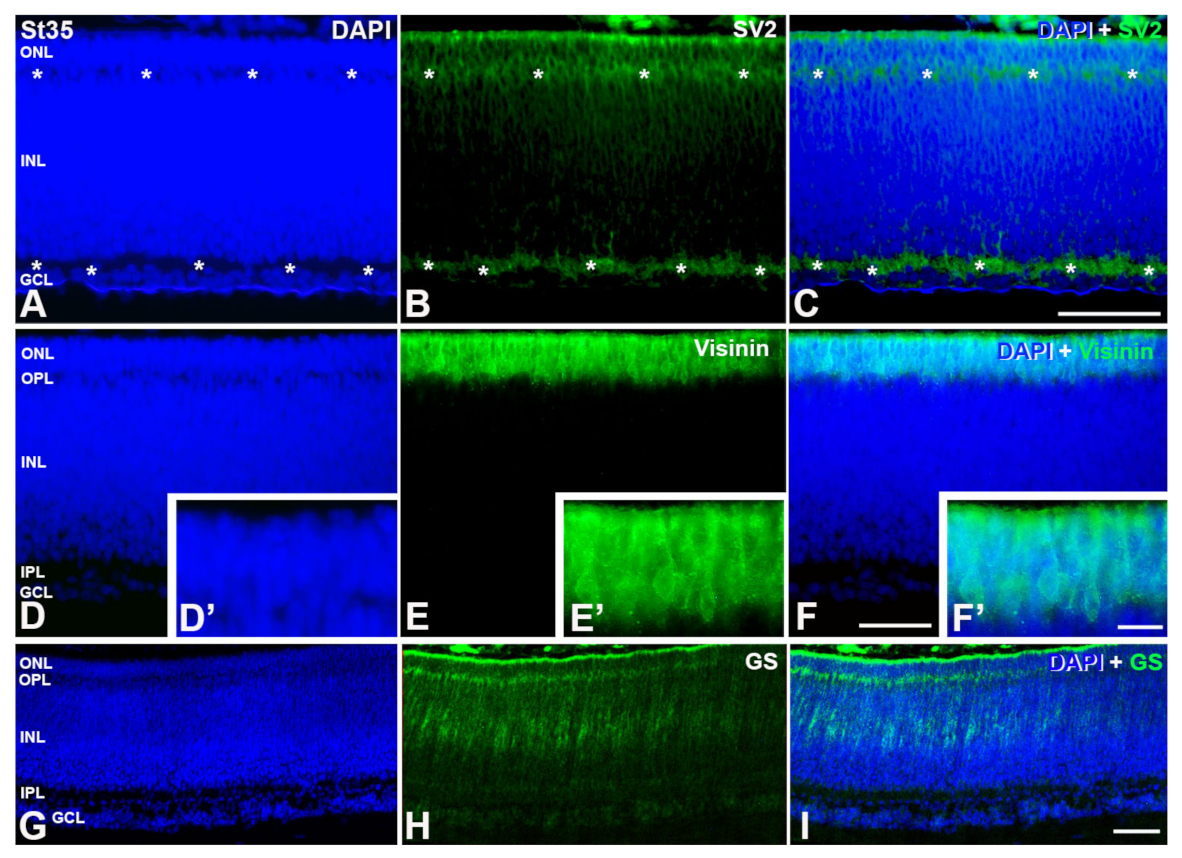
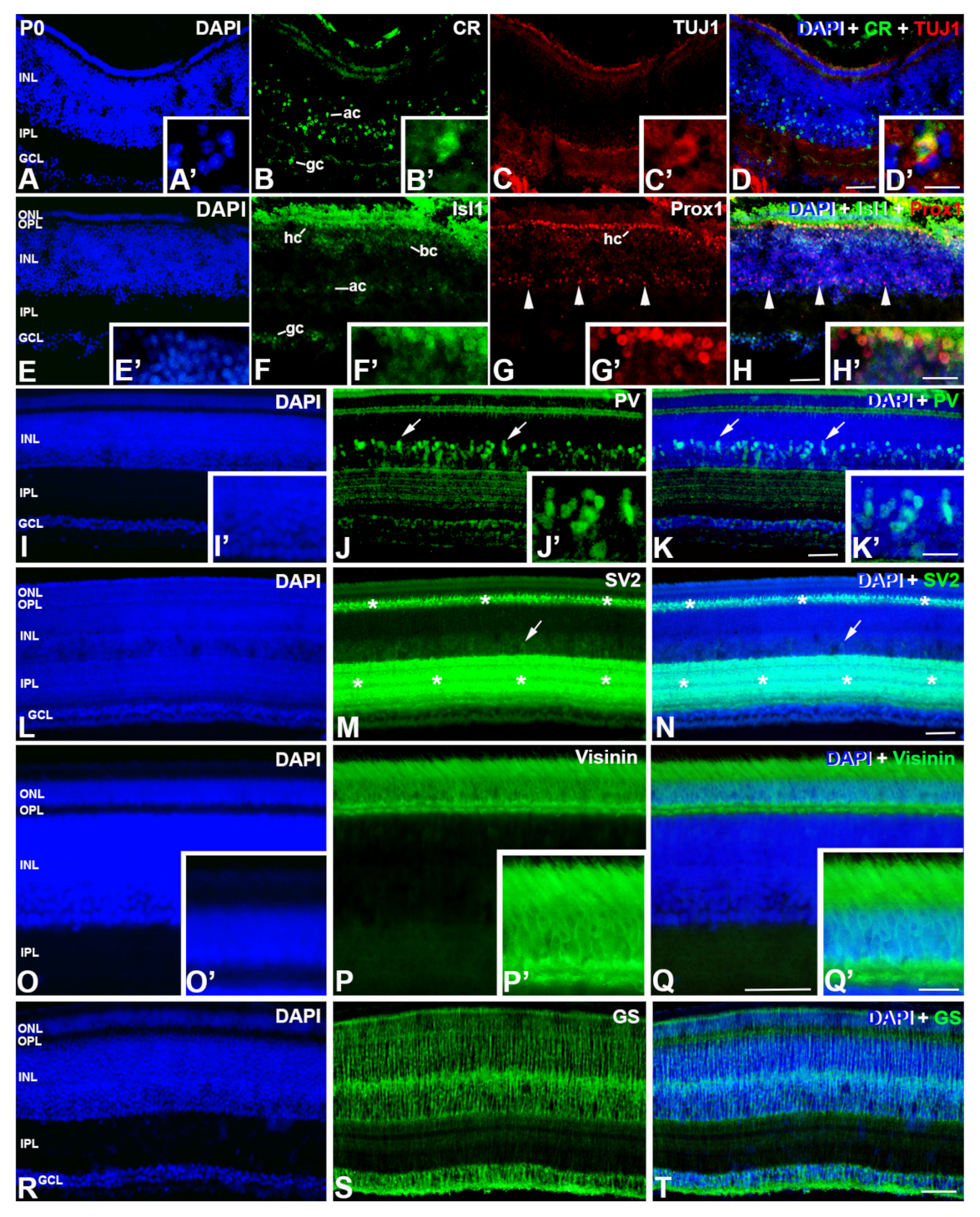
3. Results
4. Discussion
4.1. Retinal Histogenesis and Cell Differentiation in C. coturnix
4.1.1. Neurochemical Profiles in the Undifferentiated and Laminated Retina
4.1.2. Emergence of the Plexiform Layers
4.1.3. Timing of Retinal Development
4.1.4. Retinal Structure at Hatching
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bejarano-Escobar, R.; Álvarez-Hernán, G.; Morona, R.; González, A.; Martín-Partido, G.; Francisco-Morcillo, J. Expression and Function of the LIM-Homeodomain Transcription Factor Islet-1 in the Developing and Mature Vertebrate Retina. Exp. Eye Res. 2015, 138, 22–31. [Google Scholar] [CrossRef]
- Jin, K. Transitional Progenitors during Vertebrate Retinogenesis. Mol. Neurobiol. 2017, 54, 3565–3576. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Mu, X. Genetic Control of Retinal Ganglion Cell Genesis. Cell. Mol. Life Sci. 2021, 78, 4417–4433. [Google Scholar] [CrossRef]
- Diacou, R.; Nandigrami, P.; Fiser, A.; Liu, W.; Ashery-Padan, R.; Cvekl, A. Cell Fate Decisions, Transcription Factors and Signaling during Early Retinal Development. Prog. Ret. Eye Res. 2022, 91, 101093. [Google Scholar] [CrossRef]
- Petridou, E.; Godinho, L. Cellular and Molecular Determinants of Retinal Cell Fate. Annu. Rev. Vis. Sci. 2022, 8, 79–99. [Google Scholar] [CrossRef]
- Santos-França, P.L.; David, L.A.; Kassem, F.; Meng, X.Q.; Cayouette, M. Time to See: How Temporal Identity Factors Specify the Developing Mammalian Retina. Sem. Cell Dev. Biol. 2022, 142, 36–42. [Google Scholar] [CrossRef]
- Centanin, L.; Wittbrodt, J. Retinal Neurogenesis. Development 2014, 141, 241–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejarano-Escobar, R.; Blasco, M.; Durán, A.C.; Rodríguez, C.; Martín-Partido, G.; Francisco-Morcillo, J. Retinal Histogenesis and Cell Differentiation in an Elasmobranch Species, the Small-Spotted Catshark Scyliorhinus canicula: Retinogenesis in the Small-Spotted Catshark. J. Anat. 2012, 220, 318–335. [Google Scholar] [CrossRef] [PubMed]
- Calaza, K.D.C.; Gardino, P.F. Neurochemical Phenotype and Birthdating of Specific Cell Populations in the Chick Retina. An. Acad. Bras. Cienc. 2010, 82, 595–608. [Google Scholar] [CrossRef] [Green Version]
- Masland, R.H. The Neuronal Organization of the Retina. Neuron 2012, 76, 266–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergara, M.N.; Canto-Soler, M.V. Rediscovering the Chick Embryo as a Model to Study Retinal Development. Neural Dev. 2012, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Boije, H.; Shirazi Fard, S.; Edqvist, P.-H.; Hallböök, F. Horizontal Cells, the Odd Ones Out in the Retina, Give Insights into Development and Disease. Front. Neuroanat. 2016, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Langouet-Astrie, C.J.; Meinsen, A.L.; Grunwald, E.R.; Turner, S.D.; Enke, R.A. RNA Sequencing Analysis of the Developing Chicken Retina. Sci. Data 2016, 3, 160117. [Google Scholar] [CrossRef]
- Blixt, M.K.E.; Konjusha, D.; Ring, H.; Hallböök, F. Zinc Finger Gene Nolz1 Regulates the Formation of Retinal Progenitor Cells and Suppresses the Lim3/Lhx3 Phenotype of Retinal Bipolar Cells in Chicken Retina: Nolz1 in The Developing Chicken Retina. Dev. Dyn. 2018, 247, 630–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges-Martins, V.P.P.; Ferreira, D.D.P.; Souto, A.C.; Oliveira Neto, J.G.; Pereira-Figueiredo, D.; da Costa Calaza, K.; de Jesus Oliveira, K.; Manhães, A.C.; de Melo Reis, R.A.; Kubrusly, R.C.C. Caffeine Regulates GABA Transport via A1R Blockade and CAMP Signaling. Neurochem. Int. 2019, 131, 104550. [Google Scholar] [CrossRef]
- Rios, M.N.; Marchese, N.A.; Guido, M.E. Expression of Non-Visual Opsins Opn3 and Opn5 in the Developing Inner Retinal Cells of Birds. Light-Responses in Müller Glial Cells. Front. Cell. Neurosci. 2019, 13, 376. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, I.; Deistler, K.; Hoang, T.V.; Blackshaw, S.; Fischer, A.J. NF-ΚB Signaling Regulates the Formation of Proliferating Müller Glia-Derived Progenitor Cells in the Avian Retina. Development 2020, 147, dev183418. [Google Scholar] [CrossRef]
- Pereira-Figueiredo, D.; Brito, R.; Araújo, D.S.M.; Nascimento, A.A.; Lyra, E.S.B.; Cheibub, A.M.S.S.; Pereira Netto, A.D.; Ventura, A.L.M.; Paes-de-Carvalho, R.; Calaza, K.C. Caffeine Exposure Ameliorates Acute Ischemic Cell Death in Avian Developing Retina. Purinergic Signal. 2020, 16, 41–59. [Google Scholar] [CrossRef]
- Alvarez-Hernan, G.; de Mera-Rodríguez, J.A.; Gañán, Y.; Solana-Fajardo, J.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Development and Postnatal Neurogenesis in the Retina: A Comparison between Altricial and Precocial Bird Species. Neural Regen. Res. 2021, 16, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Hernán, G.; Garrido-Jiménez, S.; Román, Á.C.; Carvajal-González, J.M.; Francisco-Morcillo, J. Distribution of Planar Cell Polarity Proteins in the Developing Avian Retina. Exp. Eye Res. 2021, 209, 108681. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.J.; Hobgood, J.S.; Jia, M.; Boyd, P.; Hipp, R.I.; Hyde, D.R. Notch3 and DeltaB Maintain Müller Glia Quiescence and Act as Negative Regulators of Regeneration in the Light-damaged Zebrafish Retina. Glia 2021, 69, 546–566. [Google Scholar] [CrossRef]
- Bartley, S.C.; Proctor, M.T.; Xia, H.; Ho, E.; Kang, D.S.; Schuster, K.; Bicca, M.A.; Seckler, H.S.; Viola, K.L.; Patrie, S.M.; et al. An Essential Role for Alzheimer’s-Linked Amyloid Beta Oligomers in Neurodevelopment: Transient Expression of Multiple Proteoforms during Retina Histogenesis. Int. J. Mol. Sci. 2022, 23, 2208. [Google Scholar] [CrossRef] [PubMed]
- Gage, E.; Agarwal, D.; Chenault, C.; Washington-Brown, K.; Szvetecz, S.; Jahan, N.; Wang, Z.; Jones, M.K.; Zack, D.J.; Enke, R.A.; et al. Temporal and Isoform-Specific Expression of CTBP2 Is Evolutionarily Conserved Between the Developing Chick and Human Retina. Front. Mol. Neurosci. 2022, 14, 773356. [Google Scholar] [CrossRef] [PubMed]
- Wisely, C.E.; Sayed, J.A.; Tamez, H.; Zelinka, C.; Abdel-Rahman, M.H.; Fischer, A.J.; Cebulla, C.M. The Chick Eye in Vision Research: An Excellent Model for the Study of Ocular Disease. Prog. Retin. Eye Res. 2017, 61, 72–97. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Hernán, G.; Sánchez-Resino, E.; Hernández-Núñez, I.; Marzal, A.; Rodríguez-León, J.; Martín-Partido, G.; Francisco-Morcillo, J. Retinal Histogenesis in an Altricial Avian Species, the Zebra Finch (Taeniopygia guttata, Vieillot 1817). J. Anat. 2018, 233, 106–120. [Google Scholar] [CrossRef]
- Álvarez-Hernán, G.; Hernández-Núñez, I.; Rico-Leo, E.M.; Marzal, A.; de Mera-Rodríguez, J.A.; Rodríguez-León, J.; Martín-Partido, G.; Francisco-Morcillo, J. Retinal Differentiation in an Altricial Bird Species, Taeniopygia guttata: An Immunohistochemical Study. Exp. Eye Res. 2020, 190, 107869. [Google Scholar] [CrossRef]
- Álvarez-Hernán, G.; de Mera-Rodríguez, J.A.; Hernández-Núñez, I.; Acedo, A.; Marzal, A.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Timing and Distribution of Mitotic Activity in the Retina During Precocial and Altricial Modes of Avian Development. Front. Neurosci. 2022, 16, 853544. [Google Scholar] [CrossRef]
- Cook, B.; Portera-Cailliau, C.; Adler, R. Developmental Neuronal Death Is Not a Universal Phenomenon among Cell Types in the Chick Embryo Retina. J. Comp. Neurol. 1998, 396, 12–19. [Google Scholar] [CrossRef]
- Francisco-Morcillo, J.; Bejarano-Escobar, R.; Rodríguez-León, J.; Navascués, J.; Martín-Partido, G. Ontogenetic Cell Death and Phagocytosis in the Visual System of Vertebrates: Cell Death and Phagocytosis during Ontogeny. Dev. Dyn. 2014, 243, 1203–1225. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Hernán, G.; de Mera-Rodríguez, J.A.; Hernández-Núñez, I.; Marzal, A.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Analysis of Programmed Cell Death and Senescence Markers in the Developing Retina of an Altricial Bird Species. Cells 2021, 10, 504. [Google Scholar] [CrossRef]
- Drenhaus, U.; Voigt, T.; Rager, G. Onset of Synaptogenesis in the Plexiform Layers of the Chick Retina: A Transmission Electron Microscopic Study. Microsc. Res. Tech. 2007, 70, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Rojas, L.M.; Mitchell, M.A.; Ramírez, Y.M.; McNeil, R. Comparative analysis of retina structure and photopic electroretinograms in developing altricial pigeons (Columba livia) and precocial japanese quails (Coturnix coturnix japonica). Neotrop. Ornithol. Soc. 2007, 18, 503–518. [Google Scholar]
- Huss, D.; Poynter, G.; Lansford, R. Japanese Quail (Coturnix coturnix Japonica) as a Laboratory Animal Model. Lab. Anim. 2008, 37, 513–519. [Google Scholar] [CrossRef]
- Poynter, G.; Huss, D.; Lansford, R. Japanese Quail: An Efficient Animal Model for the Production of Transgenic Avians. Cold Spring Harb. Protoc. 2009, 2009, pdb.emo112. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, S.J.; Stanley, R.L.; Evans, D.J.R. Developmental Stages of the Japanese Quail. J. Anat. 2010, 216, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Marín-Teva, J.L.; Cuadros, M.A.; Martín-Oliva, D.; Navascués, J. Microglia and Neuronal Cell Death. Neuron Glia Biol. 2011, 7, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Martín-Estebané, M.; Navascués, J.; Sierra-Martín, A.; Martín-Guerrero, S.M.; Cuadros, M.A.; Carrasco, M.-C.; Marín-Teva, J.L. Onset of Microglial Entry into Developing Quail Retina Coincides with Increased Expression of Active Caspase-3 and Is Mediated by Extracellular ATP and UDP. PLoS ONE 2017, 12, e0182450. [Google Scholar] [CrossRef] [Green Version]
- Marín-Teva, J.L.; Cuadros, M.A.; Calvente, R.; Almendros, A.; Navascués, J. Naturally Occurring Cell Death and Migration of Microglial Precursors in the Quail Retina during Normal Development. J. Comp. Neurol. 1999, 412, 255–275. [Google Scholar] [CrossRef]
- Willbold, E.; Reinicke, M.; Lance-Jones, C.; Lagenaur, C.; Lemmon, V.; Layer, P.G. Müller Glia Stabilizes Cell Columns During Retinal Development: Lateral Cell Migration but Not Neuropil Growth Is Inhibited in Mixed Chick-Quail Retinospheroids. Eur. J. Neurosci. 1995, 7, 2277–2284. [Google Scholar] [CrossRef]
- Bejarano-Escobar, R.; Blasco, M.; Martín-Partido, G.; Francisco-Morcillo, J. Molecular Characterization of Cell Types in the Developing, Mature, and Regenerating Fish Retina. Rev. Fish Biol. Fish. 2014, 24, 127–158. [Google Scholar] [CrossRef]
- Kubota, R.; Hokoc, J.N.; Moshiri, A.; McGuire, C.; Reh, T.A. A Comparative Study of Neurogenesis in the Retinal Ciliary Marginal Zone of Homeothermic Vertebrates. Dev. Brain Res. 2002, 134, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Ghai, K.; Stanke, J.J.; Fischer, A.J. Patterning of the Circumferential Marginal Zone of Progenitors in the Chicken Retina. Brain Res. 2008, 1192, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Francisco-Morcillo, J.; Hidalgo-Sánchez, M.; Martín-Partido, G. Spatial and Temporal Patterns of Proliferation and Differentiation in the Developing Turtle Eye. Brain Res. 2006, 1103, 32–48. [Google Scholar] [CrossRef]
- Elshatory, Y.; Everhart, D.; Deng, M.; Xie, X.; Barlow, R.B.; Gan, L. Islet-1 Controls the Differentiation of Retinal Bipolar and Cholinergic Amacrine Cells. J. Neurosci. 2007, 27, 12707–12720. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Hernán, G.; Bejarano-Escobar, R.; Morona, R.; González, A.; Martín-Partido, G.; Francisco-Morcillo, J. Islet-1 Immunoreactivity in the Developing Retina of Xenopus Laevis. Sci. World J. 2013, 2013, 740420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laessing, U.; Stuermer, C.A. Spatiotemporal Pattern of Retinal Ganglion Cell Differentiation Revealed by the Expression of Neurolin in Embryonic Zebrafish. J. Neurobiol. 1996, 29, 65–74. [Google Scholar] [CrossRef]
- Mata, D.; Linn, D.M.; Linn, C.L. Retinal Ganglion Cell Neuroprotection Induced by Activation of Alpha7 Nicotinic Acetylcholine Receptors. Neuropharmacology 2015, 99, 337–346. [Google Scholar] [CrossRef]
- Prada, C.; Puga, J.; Perez-Mendez, L.; Lopez, R.; Ramirez, G. Spatial and Temporal Patterns of Neurogenesis in the Chick Retina. Eur. J. Neurosci. 1991, 3, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Snow, R.L.; Robson, J.A. Ganglion Cell Neurogenesis, Migration and Early Differentiation in the Chick Retina. Neuroscience 1994, 58, 399–409. [Google Scholar] [CrossRef]
- McCabe, K.L.; Gunther, E.C.; Reh, T.A. The Development of the Pattern of Retinal Ganglion Cells in the Chick Retina: Mechanisms That Control Differentiation. Development 1999, 126, 5713–5724. [Google Scholar] [CrossRef]
- Francisco-Morcillo, J.; Sánchez-Calderón, H.; Kawakami, Y.; Belmonte, J.C.I.; Hidalgo-Sánchez, M.; Martín-Partido, G. Expression of Fgf19 in the Developing Chick Eye. Dev. Brain Res. 2005, 156, 104–109. [Google Scholar] [CrossRef]
- De Mera-Rodríguez, J.A.; Álvarez-Hernán, G.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Senescence-associated Β-galactosidase Activity in the Developing Avian Retina. Dev. Dyn. 2019, 248, 850–865. [Google Scholar] [CrossRef]
- Edqvist, P.-H.D.; Lek, M.; Boije, H.; Lindbäck, S.M.; Hallböök, F. Axon-Bearing and Axon-Less Horizontal Cell Subtypes Are Generated Consecutively during Chick Retinal Development from Progenitors That Are Sensitive to Follistatin. BMC Dev. Biol. 2008, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boije, H.; Edqvist, P.-H.D.; Hallböök, F. Horizontal Cell Progenitors Arrest in G2-Phase and Undergo Terminal Mitosis on the Vitreal Side of the Chick Retina. Dev. Biol. 2009, 330, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, S.; Cepko, C. Development of the Pattern of Photoreceptors in the Chick Retina. J. Neurosci. 1996, 16, 1430–1439. [Google Scholar] [CrossRef]
- Fischer, A.J.; Foster, S.; Scott, M.A.; Sherwood, P. Transient Expression of LIM-Domain Transcription Factors Is Coincident with Delayed Maturation of Photoreceptors in the Chicken Retina. J. Comp. Neurol. 2008, 506, 584–603. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, M.; Grabs, D.; Rager, G. Developmental Expression of Dynamin in the Chick Retinotectal System. J. Histochem. Cytochem. 1999, 47, 1297–1306. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Layer, P.G.; Gierer, A. Binding of FITC-Coupled Peanut-Agglutinin (FITC-PNA) to Embryonic Chicken Retinas Reveals Developmental Spatio-Temporal Patterns. Dev. Brain Res. 1983, 8, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Reiss, Y.; Kröger, S.; Grassi, J.; Tsim, K.W.K.; Willbold, E.; Layer, P.G. Extracellular and Asymmetric Forms of Acetylcholinesterase Are Expressed on Cholinergic and Noncholinergic Terminal Neuropil of the Developing Chick Retina. Cell Tissue Res. 1996, 286, 13–22. [Google Scholar] [CrossRef]
- Reiss, Y.; Layer, P.G.; Kröger, S. Butyrylcholinesterase-Positive Cells of the Developing Chicken Retina That Are Non-Cholinergic and GABA-Positive. Dev. Brain Res. 1997, 100, 62–72. [Google Scholar] [CrossRef]
- Layer, P.G.; Berger, J.; Kinkl, N. Cholinesterases Precede “ON-OFF” Channel Dichotomy in the Embryonic Chick Retina before Onset of Synaptogenesis. Cell Tissue Res. 1997, 288, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Thangaraj, G.; Greif, A.; Bachmann, G.; Layer, P.G. Intricate Paths of Cells and Networks Becoming “Cholinergic” in the Embryonic Chicken Retina. J. Comp. Neurol. 2012, 520, 3181–3193. [Google Scholar] [CrossRef] [PubMed]
- Harahush, B.K.; Hart, N.S.; Green, K.; Collin, S.P. Retinal Neurogenesis and Ontogenetic Changes in the Visual System of the Brown Banded Bamboo Shark, Chiloscyllium punctatum (Hemiscyllidae, Elasmobranchii). J. Comp. Neurol. 2009, 513, 83–97. [Google Scholar] [CrossRef]
- Ferreiro-Galve, S.; Rodríguez-Moldes, I.; Anadón, R.; Candal, E. Patterns of Cell Proliferation and Rod Photoreceptor Differentiation in Shark Retinas. J. Chem. Neuroanat. 2010, 39, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bejarano-Escobar, R.; Blasco, M.; Durán, A.C.; Martín-Partido, G.; Francisco-Morcillo, J. Chronotopographical Distribution Patterns of Cell Death and of Lectin-Positive Macrophages/Microglial Cells during the Visual System Ontogeny of the Small-Spotted Catshark Scyliorhinus canicula. J. Anat. 2013, 223, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Bejarano-Escobar, R.; Blasco, M.; DeGrip, W.J.; Martín-Partido, G.; Francisco-Morcillo, J. Cell Differentiation in the Retina of an Epibenthonic Teleost, the Tench (Tinca tinca, Linneo 1758). Exp. Eye Res. 2009, 89, 398–415. [Google Scholar] [CrossRef] [PubMed]
- Bejarano-Escobar, R.; Blasco, M.; DeGrip, W.J.; Oyola-Velasco, J.A.; Martín-Partido, G.; Francisco-Morcillo, J. Eye Development and Retinal Differentiation in an Altricial Fish Species, the Senegalese sole (Solea senegalensis, Kaup 1858). J. Exp. Zool. 2010, 314B, 580–605. [Google Scholar] [CrossRef]
- Álvarez-Hernán, G.; de Mera-Rodríguez, J.A.; de la Gándara, F.; Ortega, A.; Barros-Gata, I.; Romero-Rodríguez, J.A.; Blasco, M.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Histogenesis and Cell Differentiation in the Retina of Thunnus Thynnus: A Morphological and Immunohistochemical Study. Tissue Cell 2022, 76, 101809. [Google Scholar] [CrossRef]
- Murray, J.R.; Varian-Ramos, C.W.; Welch, Z.S.; Saha, M.S. Embryological Staging of the Zebra Finch, Taeniopygia guttata. J. Morphol. 2013, 274, 1090–1110. [Google Scholar] [CrossRef] [Green Version]
- Blom, J.; Lilja, C. A Comparative Study of Growth, Skeletal Development and Eggshell Composition in Some Species of Birds. J. Zool. 2004, 262, 361–369. [Google Scholar] [CrossRef]
- Blom, J.; Lilja, C. A Comparative Study of Embryonic Development of Some Bird Species with Different Patterns of Postnatal Growth. Zoology 2005, 108, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.H.; Richards, D.E.; Rogers, J.H. Calretinin and Calbindin in the Retina of the Developing Chick. Cell Tissue Res. 1991, 264, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, K.; Ishii, A.; Shimada, N.; Yasuda, K. RaxL Regulates Chick Ganglion Cell Development. Mech. Dev. 2003, 120, 881–895. [Google Scholar] [CrossRef]
- Bradford, R.L.; Wang, C.; Zack, D.J.; Adler, R. Roles of Cell-Intrinsic and Microenvironmental Factors in Photoreceptor Cell Differentiation. Dev. Biol. 2005, 286, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edqvist, P.-H.D.; Hallböök, F. Newborn Horizontal Cells Migrate Bi-Directionally across the Neuroepithelium during Retinal Development. Development 2004, 131, 1343–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olea, G.B. Desarrollo Embrionario de Columba livia (Aves: Columbiformes) Desde Una Perspectiva Altricial-Precoz. Rev. Colomb. Cienc. Pecu. 2012, 25, 3–13. [Google Scholar]
- Starck, J.M.; Ricklefs, R.E. (Eds.) Avian Growth and Development: Evolution within the Altricial-Precocial Spectrum; Oxford Ornithology Series; Oxford University Press: New York, NY, USA, 1998; ISBN 978-0-19-510608-4. [Google Scholar]
- Evans, B.I.; Browman, H.I. Variation in the Development of the Fish Retina. Am. Fish. Soc. Symp. 2004, 40, 145–166. [Google Scholar]
- Scheiber, I.B.R.; Weiß, B.M.; Kingma, S.A.; Komdeur, J. The Importance of the Altricial—Precocial Spectrum for Social Complexity in Mammals and Birds—A Review. Front. Zool. 2017, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.J.; Reh, T.A. Identification of a Proliferating Marginal Zone of Retinal Progenitors in Postnatal Chickens. Dev. Biol. 2000, 220, 197–210. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Incubation Time | N |
|---|---|---|
| St19 | 72 h (3 days) | 5 |
| St 21 | 84 h (3.5 days) | 6 |
| St 24 | 108 h (4.5 days) | 6 |
| St 28 | 132 h (5.5 days) | 5 |
| St 31 | 6.5 days | 4 |
| St 32 | 7 days | 4 |
| St 35 | 8–9 days | 4 |
| St 42 | 13 days | 3 |
| St 46 (P0) | 16.5 days (hatching) | 3 |
| TOTAL | 40 | |
| Primary Antibodies | Working Dilution | Antibody Suppliers | Cell Type Specificity |
|---|---|---|---|
| Mouse anti-β-tubulin class III monoclonal antibody (TUJ1) | 1:200 | Abcam (Ref. ab14545) | Neuroblasts, ganglion and amacrine cells |
| Mouse anti-PCNA monoclonal antibody (clone PC10) | 1:200 | Santa Cruz Biotechnology (Ref. sc-56) | Proliferating cells |
| Mouse anti-visinin monoclonal antibody (clone 7G4) | 1:200 | Sigma Aldrich (Ref. P0089) | Photoreceptors |
| Mouse anti-islet1 monoclonal antibody (clone 40.2D6) | 1:50 | Developmental Studies Hybridoma Bank (Ref. 40.2D6) | Neuroblasts, ganglion, amacrine, bipolar and horizontal cells |
| Mouse anti-glutamine synthetase monoclonal antibody | 1:200 | Millipore (Ref. LV1412159) | Müller cells |
| Mouse anti-synaptic vesicle glycoprotein 2A (SV2) monoclonal antibody | 1:200 | Developmental Studies Hybridoma Bank (ref. AB_2315387) | Axons and some amacrine cells |
| Mouse anti-acetylated α-Tubulin monoclonal antibody | 1:200 | Santa Cruz Biotechnology (Ref. sc-3950) | Neuroblasts, ganglion and amacrine cells |
| Mouse anti-Parvalbumin monoclonal antibody | 1:200 | Millipore (Ref. MAB1572) | Subpopulation of amacrine cells |
| Rabbit anti-Prox1 polyclonal antibody | 1:200 | Millipore (Ref. AB5475) | Amacrine, horizontal cells and horizontal cell precursors |
| Rabbit anti-calretinin polyclonal antibody | 1:500 | Swant (Ref. 7697) | Ganglion and amacrine cells |
| Rabbit anti-phospho Histone H3 (Ser10) polyclonal antibody | 1:200 | Millipore (Ref. 06-570) | Mitotic cells |
| Secondary antibodies | Working dilution | Antibody suppliers | |
| Alexa Fluor 488 goat anti-mouse IgG antibody | 1:200 | Molecular Probes (Ref. A11029) | |
| Alexa Fluor 594 goat anti-mouse IgG antibody | 1:200 | Molecular Probes (Ref. A11032) | |
| Alexa Fluor 488 goat anti-rabbit IgG antibody | 1:200 | Molecular Probes (Ref. A11008) | |
| Alexa Fluor 594 goat anti-rabbit IgG antibody | 1:200 | Molecular Probes (Ref. A11037) | |
| Event | Coturnix coturnix | Gallus gallus | Taeniopygia guttata | References |
|---|---|---|---|---|
| Proliferative activity at hatching in the laminated retina | Absent | Absent * | Abundant ** | * [19,73] Fischer and Reh, (2000) ** [19,26] |
| Ganglion cell differentiation | PCNA-negative nuclei: St21-22 (96 h) | [3H]timidine HH13 (48 h) * | PCNA-negative nuclei: St24 (108 h) ** | * [49]** [26] |
| α-Tubulin expression: St21 (96 h) | Tuj1 expression: HH16 (51–56 h) * | Tuj1 expression: St24 (108 h) ** | * [74]** [26] | |
| Isl1 expression: St22 (96 h) | Isl1 expression: HH19-20 (72 h) * | Isl1 expression: St25 (120 h) ** | * [75]S * [1]** [26] | |
| Amacrine cell differentiation | Tuj1 expression: St35 (E8) | Tuj1 expression: HH34 (E8) * | Tuj1 expression: St41 (E10) ** | * [74] ** [26] |
| Isl1 expression: St35 (E8) | Isl1 expression: HH34 (E8) * | Isl1 expression: St41 (E10) ** | * [76]** [26] | |
| Photoreceptor differentiation | PCNA-negative nuclei: St28 (132 h) | [3H]timidin: HH24 (96 h) * | PCNA-negative nuclei: St28 (132 h) ** | * [49]** [26] |
| Visinin expression: St28 (132 h) | Visinin expression: HH27 (120 h) * | Visinin expression: St28 (132 h) ** | * [77]** [26] | |
| Horizontal cell differentiation | Prox1 expression: St28 (132 h) ** | Prox1 expression: HH30 (156 h)-HH32 (174 h) * | Prox1 expression: St34 (174 h) ** | * [78] ** [26,27] |
| IPL emergence | SV2 expression: St31 (156 h) | SV2 expression: HH31 (E7) * AChE-immunoreactive band (E7) ** PNA-histochemistry *** | SV2 expression: St38 (E8.5) **** | * [31] ** [63] *** [59] ** [25] |
| OPL emergence | SV2 expression: St35 (E8) | SV2 expression: HH34 (E8) * | SV2 expression: St39 (E9) ** | * [31]** [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez-Hernán, G.; de Mera-Rodríguez, J.A.; Calle-Guisado, V.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Retinal Development in a Precocial Bird Species, the Quail (Coturnix coturnix, Linnaeus 1758). Cells 2023, 12, 989. https://doi.org/10.3390/cells12070989
Álvarez-Hernán G, de Mera-Rodríguez JA, Calle-Guisado V, Martín-Partido G, Rodríguez-León J, Francisco-Morcillo J. Retinal Development in a Precocial Bird Species, the Quail (Coturnix coturnix, Linnaeus 1758). Cells. 2023; 12(7):989. https://doi.org/10.3390/cells12070989
Chicago/Turabian StyleÁlvarez-Hernán, Guadalupe, José Antonio de Mera-Rodríguez, Violeta Calle-Guisado, Gervasio Martín-Partido, Joaquín Rodríguez-León, and Javier Francisco-Morcillo. 2023. "Retinal Development in a Precocial Bird Species, the Quail (Coturnix coturnix, Linnaeus 1758)" Cells 12, no. 7: 989. https://doi.org/10.3390/cells12070989
APA StyleÁlvarez-Hernán, G., de Mera-Rodríguez, J. A., Calle-Guisado, V., Martín-Partido, G., Rodríguez-León, J., & Francisco-Morcillo, J. (2023). Retinal Development in a Precocial Bird Species, the Quail (Coturnix coturnix, Linnaeus 1758). Cells, 12(7), 989. https://doi.org/10.3390/cells12070989