
Protein Networks Associated with Native Metabotropic Glutamate 1 Receptors (mGlu1) in the Mouse Cerebellum

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Antibodies and Vectors
2.3. Immunoprecipitation of Proteins from Mouse Cerebellum (P2 Fraction)
2.4. Immunoblotting
2.5. Mass Spectrometry (LC–MS/MS) Analysis of Proteins
2.6. Generation of a Stable Cell Line Expressing KCTD12
2.7. Immunofluorescence
2.8. Transient Transfection and Immunoprecipitation
2.9. Freeze-Fracture Replica Immunogold Labeling (FRIL)
2.10. Network Construction
2.11. Network Analysis
3. Results
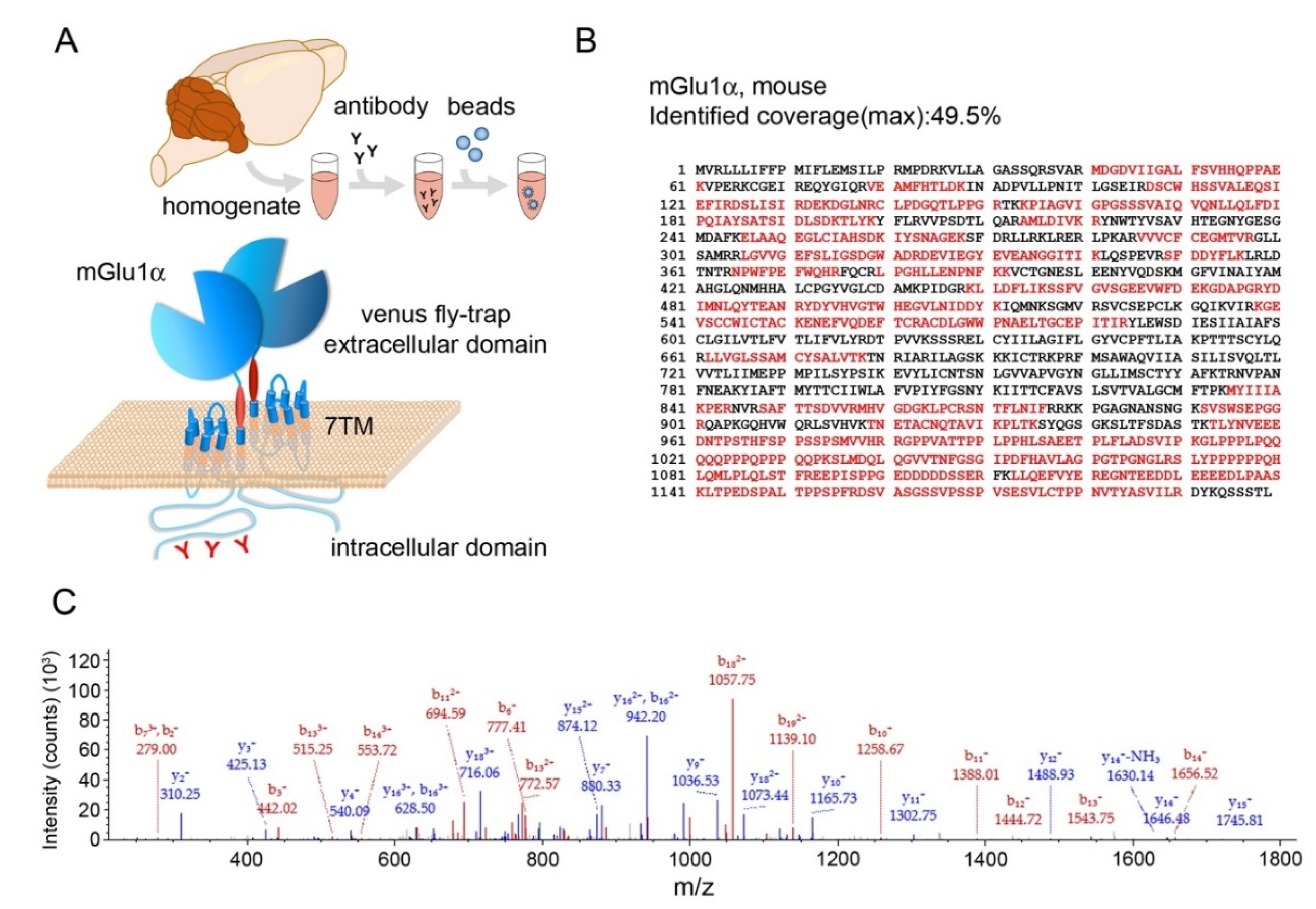
3.1. Immunoprecipitation of mGlu1α from Mouse Cerebellum
3.2. mGlu1α Co-Immunoprecipitated Proteins
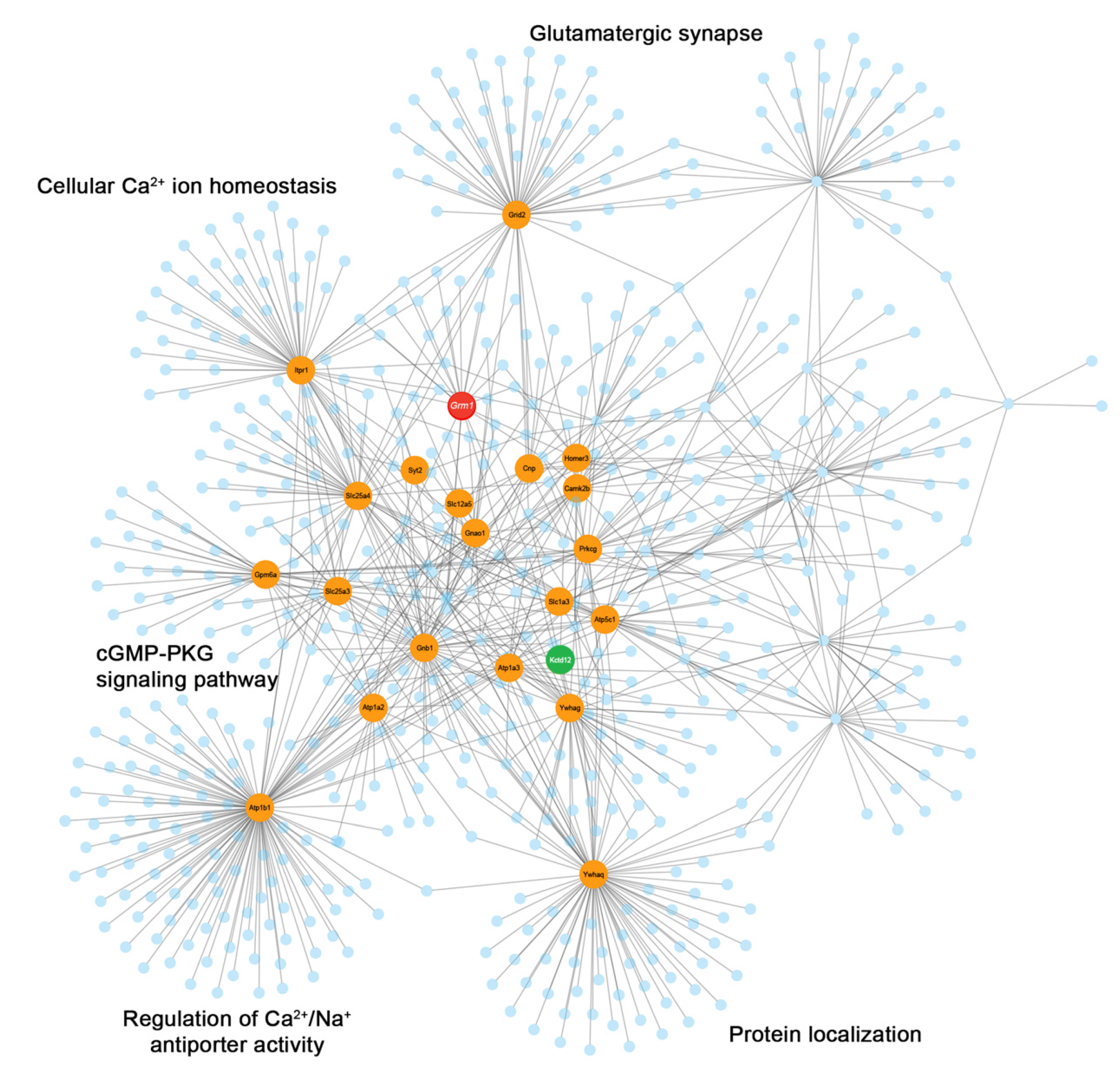
3.3. mGlu1 Interactome
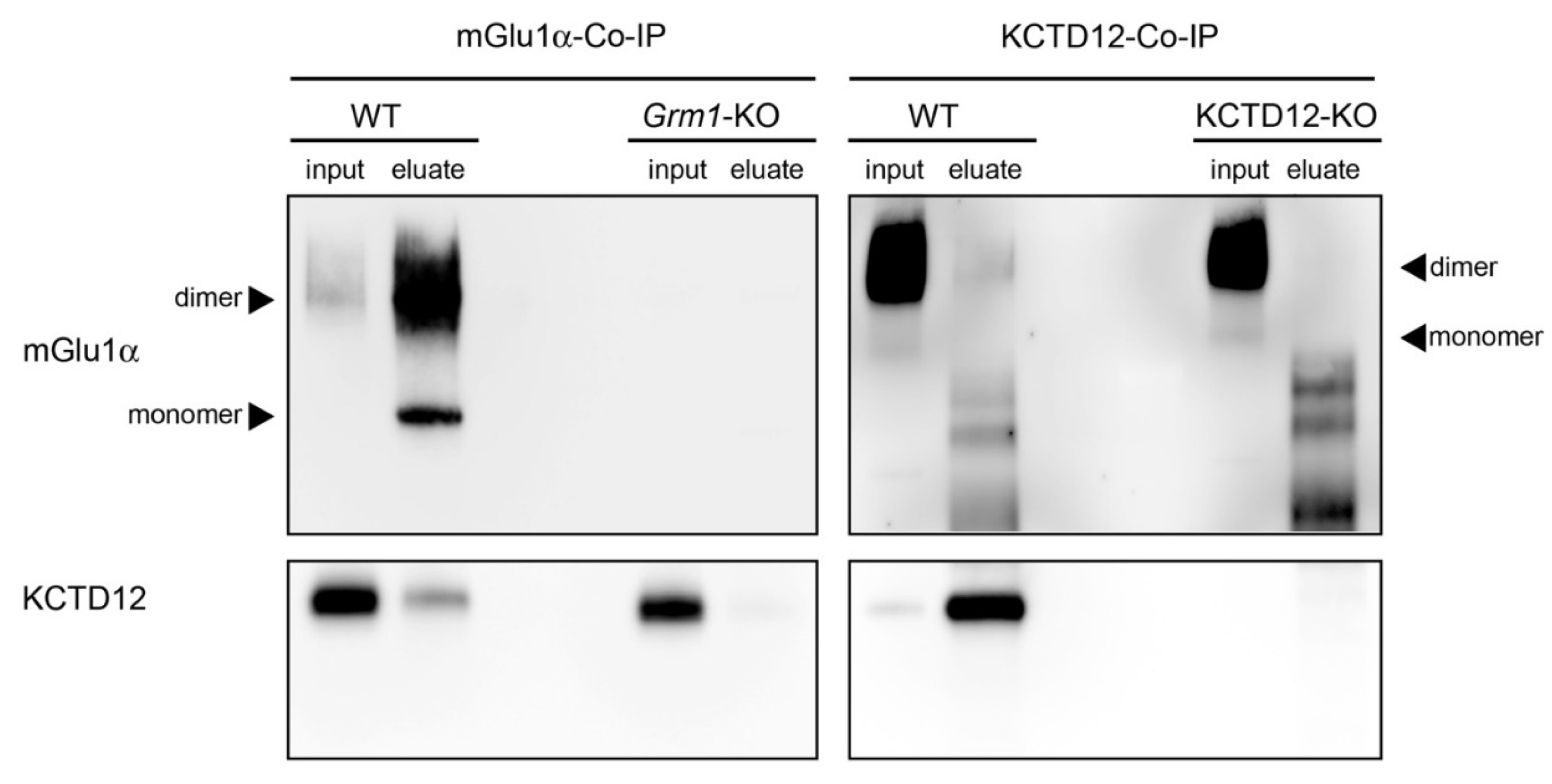
3.4. Interaction between mGlu1α and KCTD12 in the Cerebellum
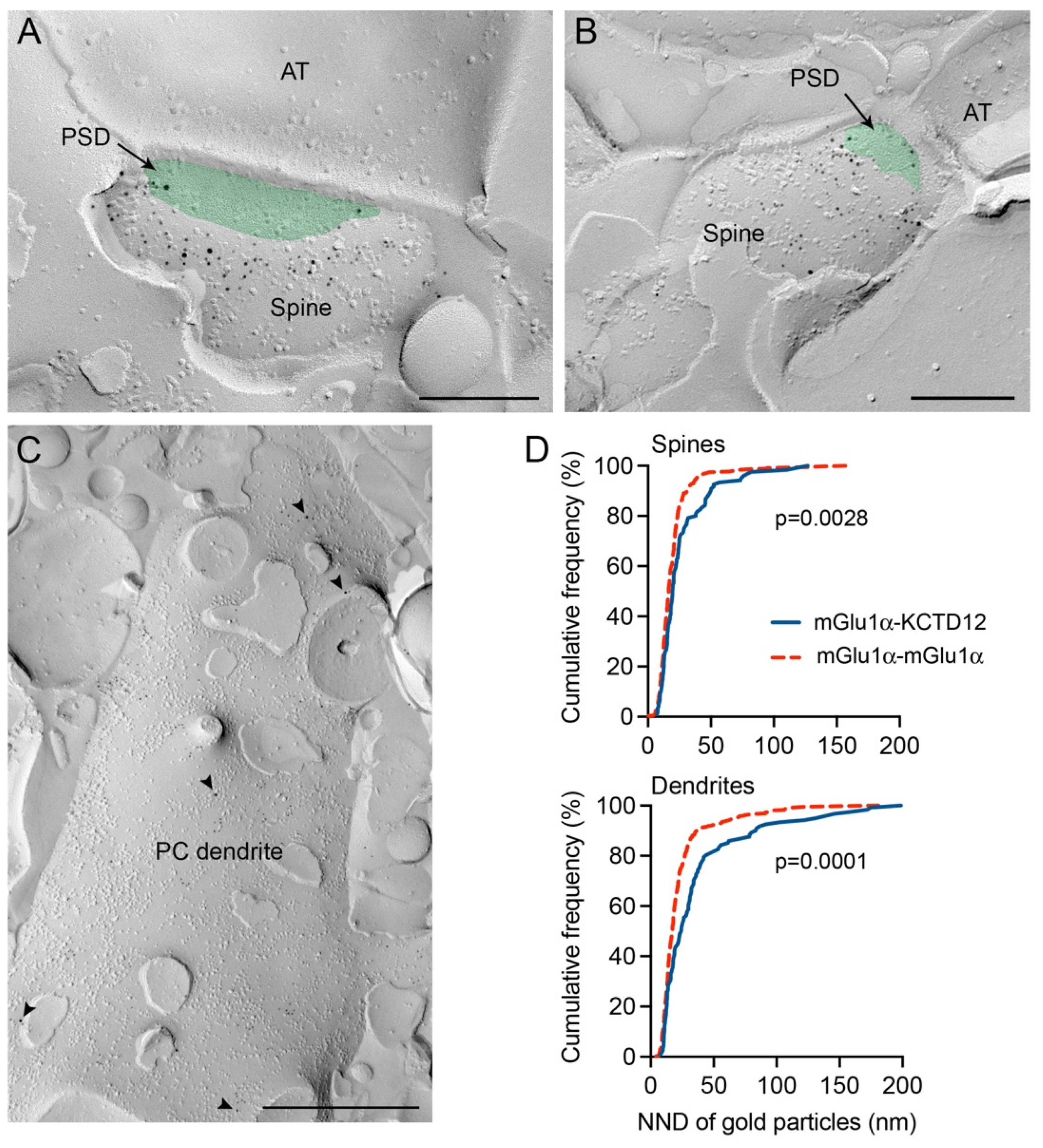
3.5. The Co-Existence of mGlu1α and KCTD12 in the Same Microdomain of Cerebellar Purkinje Cell Dendritic Spines
3.6. In Vitro Analysis of the mGlu1α–KCTD12 Interaction
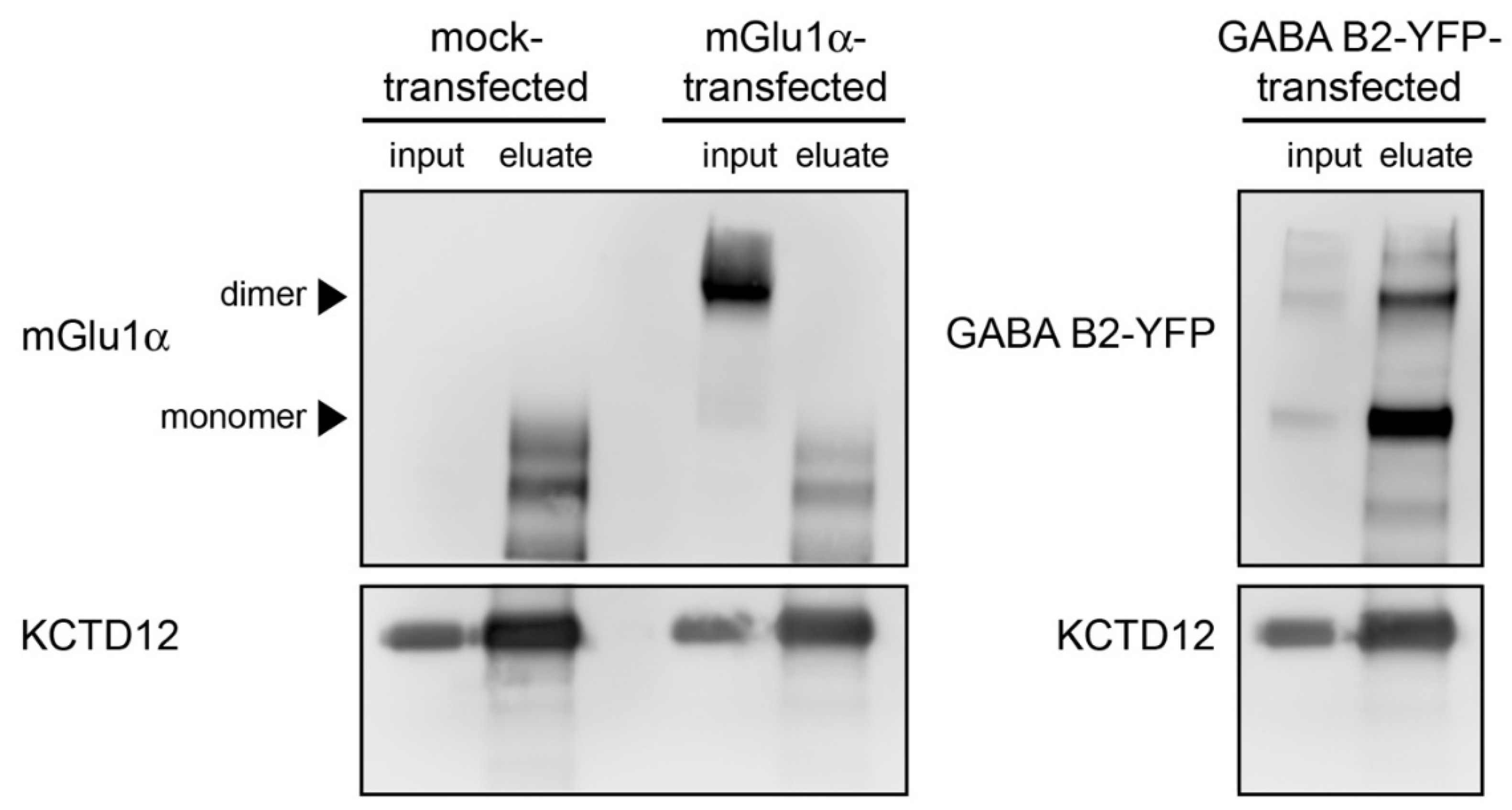
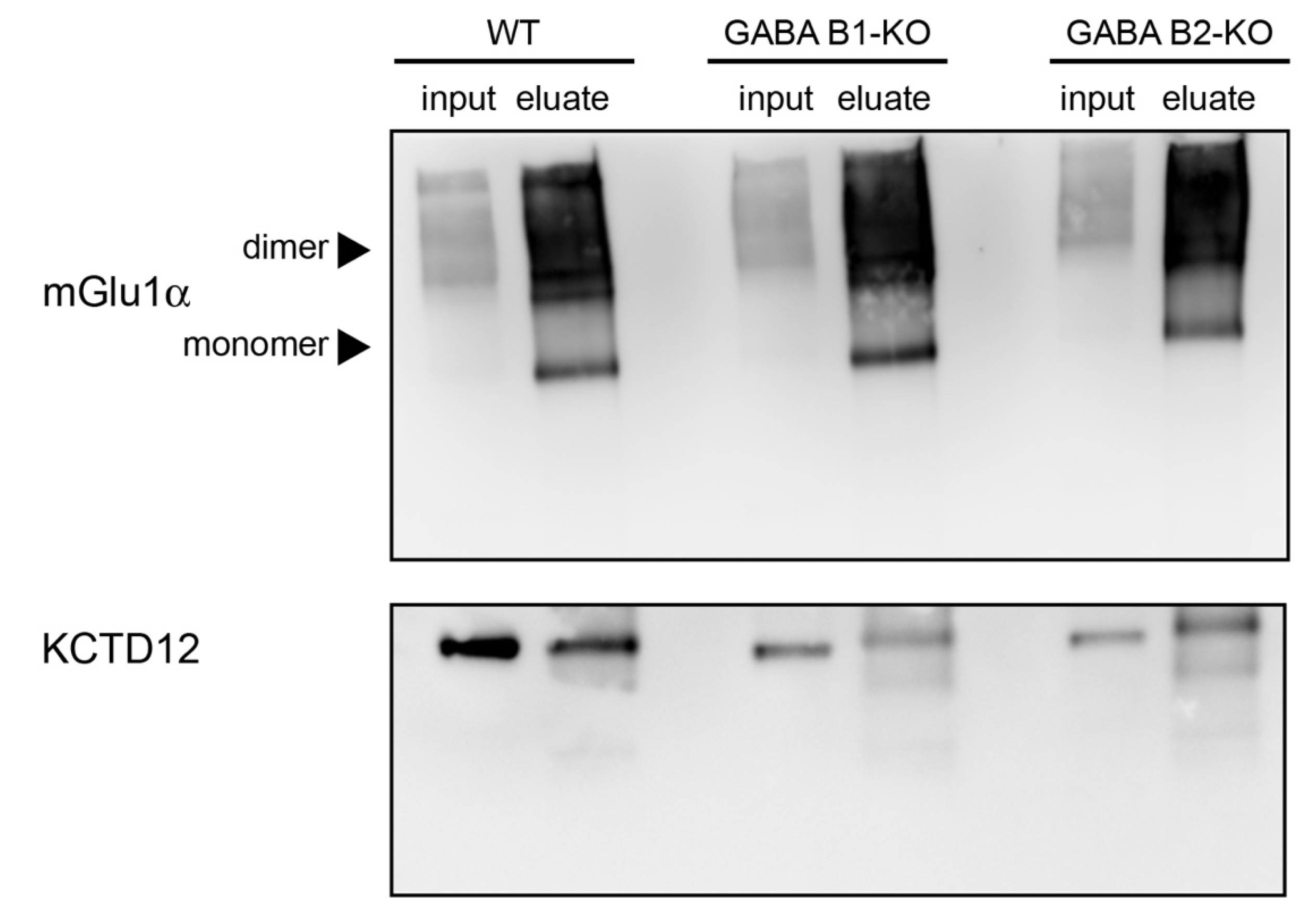
3.7. The mGlu1α Receptor–KCTD12 Interaction Is Not Mediated by GABAB Receptors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferraguti, F.; Crepaldi, L.; Nicoletti, F. Metabotropic glutamate 1 receptor: Current concepts and perspectives. Pharmacol. Rev. 2008, 60, 536–581. [Google Scholar] [CrossRef] [PubMed]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [PubMed]
- Ferraguti, F.; Shigemoto, R. Metabotropic glutamate receptors. Cell Tissue Res. 2006, 326, 483–504. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J.; Blackstone, C.D.; Huganir, R.L.; Price, D.L. Cellular localization of a metabotropic glutamate receptor in rat brain. Neuron 1992, 9, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Shigemoto, R.; Nakanishi, S.; Mizuno, N. Distribution of the mRNA for a metabotropic glutamate receptor (mGluR1) in the central nervous system: An in situ hybridization study in adult and developing rat. J. Comp. Neurol. 1992, 322, 121–135. [Google Scholar] [CrossRef]
- Shigemoto, R.; Mizuno, N. Chapter III Metabotropic glutamate receptors—immunocytochemical and in situ hybridization analyses. In Handbook of Chemical Neuroanatomy; Elsevier: Amsterdam, The Netherlands, 2000; pp. 63–98. [Google Scholar] [CrossRef]
- Négyessy, L.; Vidnyánszky, Z.; Kuhn, R.; Knöpfel, T.; Görcs, T.J.; Hámori, J. Light and electron microscopic demonstration of mGluR5 metabotropic glutamate receptor immunoreactive neuronal elements in the rat cerebellar cortex. J. Comp. Neurol. 1997, 385, 641–650. [Google Scholar] [CrossRef]
- Ito, M. Cerebellar circuitry as a neuronal machine. Prog. Neurobiol. 2006, 78, 272–303. [Google Scholar] [CrossRef]
- Aiba, A.; Chen, C.; Herrup, K.; Rosenmund, C.; Stevens, C.F.; Tonegawa, S. Reduced hippocampal long-term potentiation and context-specific deficit in associative learning in mGluR1 mutant mice. Cell 1994, 79, 365–375. [Google Scholar] [CrossRef]
- Conquet, F.; Bashir, Z.I.; Davies, C.H.; Daniel, H.; Ferraguti, F.; Bordi, F.; Franz-Bacon, K.; Reggiani, A.; Matarese, V.; Condé, F. Motor deficit and impairment of synaptic plasticity in mice lacking mGluR1. Nature 1994, 372, 237–243. [Google Scholar] [CrossRef]
- Kano, M.; Hashimoto, K.; Kurihara, H.; Watanabe, M.; Inoue, Y.; Aiba, A.; Tonegawa, S. Persistent multiple climbing fiber innervation of cerebellar Purkinje cells in mice lacking mGluR1. Neuron 1997, 18, 71–79. [Google Scholar] [CrossRef]
- Kano, M.; Hashimoto, K.; Tabata, T. Type-1 metabotropic glutamate receptor in cerebellar Purkinje cells: A key molecule responsible for long-term depression, endocannabinoid signalling and synapse elimination. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2008, 363, 2173–2186. [Google Scholar] [CrossRef]
- Ichise, T.; Kano, M.; Hashimoto, K.; Yanagihara, D.; Nakao, K.; Shigemoto, R.; Katsuki, M.; Aiba, A. mGluR1 in cerebellar Purkinje cells essential for long-term depression, synapse elimination, and motor coordination. Science 2000, 288, 1832–1835. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Kawaguchi, S.; Hirano, T. mGluR1-mediated facilitation of long-term potentiation at inhibitory synapses on a cerebellar Purkinje neuron. Eur. J. Neurosci. 2008, 27, 884–896. [Google Scholar] [CrossRef]
- Enz, R. Metabotropic glutamate receptors and interacting proteins: Evolving drug targets. Curr. Drug Targets 2012, 13, 145–156. [Google Scholar] [CrossRef]
- Kalinowska, M.; Francesconi, A. Group I Metabotropic Glutamate Receptor Interacting Proteins: Fine-Tuning Receptor Functions in Health and Disease. Curr. Neuropharmacol. 2016, 14, 494–503. [Google Scholar] [CrossRef]
- Pin, J.-P.; Bettler, B. Organization and functions of mGlu and GABAB receptor complexes. Nature 2016, 540, 60–68. [Google Scholar] [CrossRef]
- Suh, Y.H.; Chang, K.; Roche, K.W. Metabotropic glutamate receptor trafficking. Mol. Cell. Neurosci. 2018, 91, 10–24. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, S.; Xiong, Y.; Ma, Y.; Luo, D.; Jeromin, A.; Zhang, H.; He, J. A novel association of mGluR1a with the PDZ scaffold protein CAL modulates receptor activity. FEBS Lett. 2008, 582, 4117–4124. [Google Scholar] [CrossRef]
- Tu, J.C.; Xiao, B.; Yuan, J.P.; Lanahan, A.A.; Leoffert, K.; Li, M.; Linden, D.J.; Worley, P.F. Homer binds a novel proline-rich motif and links group 1 metabotropic glutamate receptors with IP3 receptors. Neuron 1998, 21, 717–726. [Google Scholar] [CrossRef]
- Xiao, B.; Tu, J.C.; Worley, P.F. Homer: A link between neural activity and glutamate receptor function. Curr. Opin. Neurobiol. 2000, 10, 370–374. [Google Scholar] [CrossRef]
- Wang, H.; Westin, L.; Nong, Y.; Birnbaum, S.; Bendor, J.; Brismar, H.; Nestler, E.; Aperia, A.; Flajolet, M.; Greengard, P. Norbin is an endogenous regulator of metabotropic glutamate receptor 5 signaling. Science 2009, 326, 1554–1557. [Google Scholar] [CrossRef] [PubMed]
- Ojha, P.; Pal, S.; Bhattacharyya, S. Regulation of Metabotropic Glutamate Receptor Internalization and Synaptic AMPA Receptor Endocytosis by the Postsynaptic Protein Norbin. J. Neurosci. 2022, 42, 731–748. [Google Scholar] [CrossRef] [PubMed]
- Croci, C.; Sticht, H.; Brandstätter, J.H.; Enz, R. Group I metabotropic glutamate receptors bind to protein phosphatase 1C. Mapping and modeling of interacting sequences. J. Biol. Chem. 2003, 278, 50682–50690. [Google Scholar] [CrossRef] [PubMed]
- Moriyoshi, K.; Iijima, K.; Fujii, H.; Ito, H.; Cho, Y.; Nakanishi, S. Seven in absentia homolog 1A mediates ubiquitination and degradation of group 1 metabotropic glutamate receptors. Proc. Natl. Acad. Sci. USA 2004, 101, 8614–8619. [Google Scholar] [CrossRef]
- Pandey, S.; Ramsakha, N.; Sharma, R.; Gulia, R.; Ojha, P.; Lu, W.; Bhattacharyya, S. The post-synaptic scaffolding protein tamalin regulates ligand-mediated trafficking of metabotropic glutamate receptors. J. Biol. Chem. 2020, 295, 8575–8588. [Google Scholar] [CrossRef]
- Uemura, T.; Mori, H.; Mishina, M. Direct interaction of GluRδ2 with Shank scaffold proteins in cerebellar Purkinje cells. Mol. Cell. Neurosci. 2004, 26, 330–341. [Google Scholar] [CrossRef]
- Hartmann, J.; Dragicevic, E.; Adelsberger, H.; Henning, H.A.; Sumser, M.; Abramowitz, J.; Blum, R.; Dietrich, A.; Freichel, M.; Flockerzi, V.; et al. TRPC3 channels are required for synaptic transmission and motor coordination. Neuron 2008, 59, 392–398. [Google Scholar] [CrossRef]
- Hartmann, J.; Henning, H.A.; Konnerth, A. mGluR1/TRPC3-mediated Synaptic Transmission and Calcium Signaling in Mammalian Central Neurons. Cold Spring Harb. Perspect. Biol. 2011, 3, a006726. [Google Scholar] [CrossRef]
- Bockaert, J.; Perroy, J.; Bécamel, C.; Marin, P.; Fagni, L. GPCR interacting proteins (GIPs) in the nervous system: Roles in physiology and pathologies. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 89–109. [Google Scholar] [CrossRef]
- Kato, A.S.; Knierman, M.D.; Siuda, E.R.; Isaac, J.T.R.; Nisenbaum, E.S.; Bredt, D.S. Glutamate Receptor 2 Associates with Metabotropic Glutamate Receptor 1 (mGluR1), Protein Kinase C, and Canonical Transient Receptor Potential 3 and Regulates mGluR1-Mediated Synaptic Transmission in Cerebellar Purkinje Neurons. J. Neurosci. 2012, 32, 15296–15308. [Google Scholar] [CrossRef]
- Fingleton, E.; Li, Y.; Roche, K.W. Advances in Proteomics Allow Insights Into Neuronal Proteomes. Front. Mol. Neurosci. 2021, 14, 647451. [Google Scholar] [CrossRef]
- Van Gelder, C.A.G.H.; Altelaar, M. Neuroproteomics of the synapse: Subcellular quantification of protein networks and signaling dynamics. Mol. Cell. Proteomics 2021, 20, 100087. [Google Scholar] [CrossRef]
- Schuler, V.; Lüscher, C.; Blanchet, C.; Klix, N.; Sansig, G.; Klebs, K.; Schmutz, M.; Heid, J.; Gentry, C.; Urban, L.; et al. Epilepsy, hyperalgesia, impaired memory, and loss of pre- and postsynaptic GABAB responses in mice lacking GABAB(1). Neuron 2001, 31, 47–58. [Google Scholar] [CrossRef]
- Gassmann, M.; Shaban, H.; Vigot, R.; Sansig, G.; Haller, C.; Barbieri, S.; Humeau, Y.; Schuler, V.; Müller, M.; Kinzel, B.; et al. Redistribution of GABAB(1) protein and atypical GABAB responses in GABAB(2)-deficient mice. J. Neurosci. 2004, 24, 6086–6097. [Google Scholar] [CrossRef]
- Cathomas, F.; Stegen, M.; Sigrist, H.; Schmid, L.; Seifritz, E.; Gassmann, M.; Bettler, B.; Pryce, C.R. Altered emotionality and neuronal excitability in mice lacking KCTD12, an auxiliary subunit of GABAB receptors associated with mood disorders. Transl. Psychiatry 2015, 5, e510. [Google Scholar] [CrossRef]
- Metz, M.; Gassmann, M.; Fakler, B.; Schaeren-Wiemers, N.; Bettler, B. Distribution of the auxiliary GABAB receptor subunits KCTD8, 12, 12b, and 16 in the mouse brain. J. Comp. Neurol. 2011, 519, 1435–1454. [Google Scholar] [CrossRef]
- Fritzius, T.; Turecek, R.; Seddik, R.; Kobayashi, H.; Tiao, J.; Rem, P.D.; Metz, M.; Kralikova, M.; Bouvier, M.; Gassmann, M.; et al. KCTD hetero-oligomers confer unique kinetic properties on hippocampal GABAB receptor-induced K+ currents. J. Neurosci. 2017, 37, 1162–1175. [Google Scholar] [CrossRef]
- Faserl, K.; Chetwynd, A.J.; Lynch, I.; Thorn, J.A.; Lindner, H.H. Corona Isolation Method Matters: Capillary Electrophoresis Mass Spectrometry Based Comparison of Protein Corona Compositions Following On-Particle versus In-Solution or In-Gel Digestion. Nanomaterials 2019, 9, 898. [Google Scholar] [CrossRef]
- Kasugai, Y.; Vogel, E.; Hörtnagl, H.; Schönherr, S.; Paradiso, E.; Hauschild, M.; Göbel, G.; Milenkovic, I.; Peterschmitt, Y.; Tasan, R.; et al. Structural and Functional Remodeling of Amygdala GABAergic Synapses in Associative Fear Learning. Neuron 2019, 104, 781–794.e4. [Google Scholar] [CrossRef]
- Schönherr, S.; Seewald, A.; Kasugai, Y.; Bosch, D.; Ehrlich, I.; Ferraguti, F. Combined Optogenetic and Freeze-fracture Replica Immunolabeling to Examine Input-specific Arrangement of Glutamate Receptors in the Mouse Amygdala. J. Vis. Exp. 2016. [Google Scholar] [CrossRef]
- Zotenko, E.; Mestre, J.; O’Leary, D.P.; Przytycka, T.M. Why do hubs in the yeast protein interaction network tend to be essential: Reexamining the connection between the network topology and essentiality. PLoS Comput. Biol. 2008, 4, e1000140. [Google Scholar] [CrossRef]
- Zhang, S.; Jin, G.; Zhang, X.-S.; Chen, L. Discovering functions and revealing mechanisms at molecular level from biological networks. Proteomics 2007, 7, 2856–2869. [Google Scholar] [CrossRef] [PubMed]
- Joy, M.P.; Brock, A.; Ingber, D.E.; Huang, S. High-betweenness proteins in the yeast protein interaction network. J. Biomed. Biotechnol. 2005, 2005, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 2009, 10, 48. [Google Scholar] [CrossRef]
- Kumpost, J.; Syrova, Z.; Kulihova, L.; Frankova, D.; Bologna, J.-C.; Hlavackova, V.; Prezeau, L.; Kralikova, M.; Hruskova, B.; Pin, J.-P.; et al. Surface expression of metabotropic glutamate receptor variants mGluR1a and mGluR1b in transfected HEK293 cells. Neuropharmacology 2008, 55, 409–418. [Google Scholar] [CrossRef]
- Techlovská, Š.; Chambers, J.N.; Dvořáková, M.; Petralia, R.S.; Wang, Y.-X.; Hájková, A.; Nová, A.; Franková, D.; Prezeau, L.; Blahos, J. Metabotropic glutamate receptor 1 splice variants mGluR1a and mGluR1b combine in mGluR1a/b dimers in vivo. Neuropharmacology 2014, 86, 329–336. [Google Scholar] [CrossRef]
- Naito, R.; Kassai, H.; Sakai, Y.; Schönherr, S.; Fukaya, M.; Schwarzer, C.; Sakagami, H.; Nakao, K.; Aiba, A.; Ferraguti, F. New Features on the Expression and Trafficking of mGluR1 Splice Variants Exposed by Two Novel Mutant Mouse Lines. Front. Mol. Neurosci. 2018, 11, 439. [Google Scholar] [CrossRef]
- Ohtani, Y.; Miyata, M.; Hashimoto, K.; Tabata, T.; Kishimoto, Y.; Fukaya, M.; Kase, D.; Kassai, H.; Nakao, K.; Hirata, T.; et al. The synaptic targeting of mGluR1 by its carboxyl-terminal domain is crucial for cerebellar function. J. Neurosci. 2014, 34, 2702–2712. [Google Scholar] [CrossRef]
- Mundell, S.J.; Matharu, A.-L.; Pula, G.; Holman, D.; Roberts, P.J.; Kelly, E. Metabotropic Glutamate Receptor 1 Internalization Induced by Muscarinic Acetylcholine Receptor Activation: Differential Dependency of Internalization of Splice Variants on Nonvisual Arrestins. Mol. Pharmacol. 2002, 61, 1114–1123. [Google Scholar] [CrossRef]
- Ady, V.; Perroy, J.; Tricoire, L.; Piochon, C.; Dadak, S.; Chen, X.; Dusart, I.; Fagni, L.; Lambolez, B.; Levenes, C. Type 1 metabotropic glutamate receptors (mGlu1) trigger the gating of GluD2 delta glutamate receptors. EMBO Rep. 2014, 15, 103–109. [Google Scholar] [CrossRef]
- Kitano, J.; Nishida, M.; Itsukaichi, Y.; Minami, I.; Ogawa, M.; Hirano, T.; Mori, Y.; Nakanishi, S. Direct interaction and functional coupling between metabotropic glutamate receptor subtype 1 and voltage-sensitive Cav2.1 Ca2+ channel. J. Biol. Chem. 2003, 278, 25101–25108. [Google Scholar] [CrossRef]
- Werthmann, R.C.; Tzouros, M.; Lamerz, J.; Augustin, A.; Fritzius, T.; Trovò, L.; Stawarski, M.; Raveh, A.; Diener, C.; Fischer, C.; et al. Symmetric signal transduction and negative allosteric modulation of heterodimeric mGlu1/5 receptors. Neuropharmacology 2021, 190, 108426. [Google Scholar] [CrossRef]
- Doumazane, E.; Scholler, P.; Zwier, J.M.; Trinquet, E.; Rondard, P.; Pin, J. A new approach to analyze cell surface protein complexes reveals specific heterodimeric metabotropic glutamate receptors. FASEB J. 2011, 25, 66–77. [Google Scholar] [CrossRef]
- Hanson, J.E.; Smith, Y. Group I metabotropic glutamate receptors at GABAergic synapses in monkeys. J. Neurosci. 1999, 19, 6488–6496. [Google Scholar] [CrossRef]
- Mansouri, M.; Kasugai, Y.; Fukazawa, Y.; Bertaso, F.; Raynaud, F.; Perroy, J.; Fagni, L.; Kaufmann, W.A.; Watanabe, M.; Shigemoto, R.; et al. Distinct subsynaptic localization of type 1 metabotropic glutamate receptors at glutamatergic and GABAergic synapses in the rodent cerebellar cortex. Eur. J. Neurosci. 2015, 41, 157–167. [Google Scholar] [CrossRef]
- Gassmann, M.; Bettler, B. Regulation of neuronal GABAB receptor functions by subunit composition. Nat. Rev. Neurosci. 2012, 13, 380–394. [Google Scholar] [CrossRef]
- Zheng, S.; Abreu, N.; Levitz, J.; Kruse, A.C. Structural basis for KCTD-mediated rapid desensitization of GABAB signalling. Nature 2019, 567, 127–131. [Google Scholar] [CrossRef]
- Zuo, H.; Glaaser, I.; Zhao, Y.; Kurinov, I.; Mosyak, L.; Wang, H.; Liu, J.; Park, J.; Frangaj, A.; Sturchler, E.; et al. Structural basis for auxiliary subunit KCTD16 regulation of the GABAB receptor. Proc. Natl. Acad. Sci. USA 2019, 116, 8370–8379. [Google Scholar] [CrossRef]
- Schwenk, J.; Metz, M.; Zolles, G.; Turecek, R.; Fritzius, T.; Bildl, W.; Tarusawa, E.; Kulik, A.; Unger, A.; Ivankova, K.; et al. Native GABAB receptors are heteromultimers with a family of auxiliary subunits. Nature 2010, 465, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, M.; Aiba, A.; Kano, M.; Watanabe, M. mGluR1 signaling in cerebellar Purkinje cells: Subcellular organization and involvement in cerebellar function and disease. Neuropharmacology 2021, 194, 108629. [Google Scholar] [CrossRef] [PubMed]
- Ferraguti, F.; Baldani-Guerra, B.; Corsi, M.; Nakanishi, S.; Corti, C. Activation of the extracellular signal-regulated kinase 2 by metabotropic glutamate receptors. Eur. J. Neurosci. 1999, 11, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Turecek, R.; Schwenk, J.; Fritzius, T.; Ivankova, K.; Zolles, G.; Adelfinger, L.; Jacquier, V.; Besseyrias, V.; Gassmann, M.; Schulte, U.; et al. Auxiliary GABAB receptor subunits uncouple G protein βγ subunits from effector channels to induce desensitization. Neuron 2014, 82, 1032–1044. [Google Scholar] [CrossRef] [PubMed]
- Baude, A.; Nusser, Z.; Roberts, J.D.; Mulvihill, E.; McIlhinney, R.A.; Somogyi, P. The metabotropic glutamate receptor (mGluR1 alpha) is concentrated at perisynaptic membrane of neuronal subpopulations as detected by immunogold reaction. Neuron 1993, 11, 771–787. [Google Scholar] [CrossRef] [PubMed]
- Kulik, A.; Nakadate, K.; Nyíri, G.; Notomi, T.; Malitschek, B.; Bettler, B.; Shigemoto, R. Distinct localization of GABAB receptors relative to synaptic sites in the rat cerebellum and ventrobasal thalamus. Eur. J. Neurosci. 2002, 15, 291–307. [Google Scholar] [CrossRef]
- Mao, L.-M.; Bodepudi, A.; Chu, X.-P.; Wang, J.Q. Group I Metabotropic Glutamate Receptors and Interacting Partners: An Update. Int. J. Mol. Sci. 2022, 23, 840. [Google Scholar] [CrossRef]
- Mundell, S.J.; Pula, G.; McIlhinney, R.A.J.; Roberts, P.J.; Kelly, E. Desensitization and internalization of metabotropic glutamate receptor 1a following activation of heterologous Gq/11-coupled receptors. Biochemistry 2004, 43, 7541–7551. [Google Scholar] [CrossRef]
- Jin, D.-Z.; Guo, M.-L.; Xue, B.; Fibuch, E.E.; Choe, E.S.; Mao, L.-M.; Wang, J.Q. Phosphorylation and feedback regulation of metabotropic glutamate receptor 1 by calcium/calmodulin-dependent protein kinase II. J. Neurosci. 2013, 33, 3402–3412. [Google Scholar] [CrossRef]
- van Woerden, G.M.; Hoebeek, F.E.; Gao, Z.; Nagaraja, R.Y.; Hoogenraad, C.C.; Kushner, S.A.; Hansel, C.; De Zeeuw, C.I.; Elgersma, Y. betaCaMKII controls the direction of plasticity at parallel fiber-Purkinje cell synapses. Nat. Neurosci. 2009, 12, 823–825. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, Y. 14-3-3 Proteins in Glutamatergic Synapses. Neural Plast. 2018, 2018, 8407609. [Google Scholar] [CrossRef]
- Abramow-Newerly, M.; Roy, A.A.; Nunn, C.; Chidiac, P. RGS proteins have a signalling complex: Interactions between RGS proteins and GPCRs, effectors, and auxiliary proteins. Cell. Signal. 2006, 18, 579–591. [Google Scholar] [CrossRef]
- Gabriel, L.; Lvov, A.; Orthodoxou, D.; Rittenhouse, A.R.; Kobertz, W.R.; Melikian, H.E. The acid-sensitive, anesthetic-activated potassium leak channel, KCNK3, is regulated by 14-3-3β-dependent, protein kinase C (PKC)-mediated endocytic trafficking. J. Biol. Chem. 2012, 287, 32354–32366. [Google Scholar] [CrossRef]
- Pandya, N.J.; Klaassen, R.V.; van der Schors, R.C.; Slotman, J.A.; Houtsmuller, A.; Smit, A.B.; Li, K.W. Group 1 metabotropic glutamate receptors 1 and 5 form a protein complex in mouse hippocampus and cortex. Proteomics 2016, 16, 2698–2705. [Google Scholar] [CrossRef]
- Shrivastava, A.N.; Triller, A.; Melki, R. Cell biology and dynamics of Neuronal Na+/K+-ATPase in health and diseases. Neuropharmacology 2020, 169, 107461. [Google Scholar] [CrossRef]
- Aittaleb, M.; Boguth, C.A.; Tesmer, J.J.G. Structure and function of heterotrimeric G protein-regulated Rho guanine nucleotide exchange factors. Mol. Pharmacol. 2010, 77, 111–125. [Google Scholar] [CrossRef]
- Ba, W.; Nadif Kasri, N. RhoGTPases at the synapse: An embarrassment of choice. Small GTPases 2017, 8, 106–113. [Google Scholar] [CrossRef]
- Hirono, M.; Yoshioka, T.; Konishi, S. GABA(B) receptor activation enhances mGluR-mediated responses at cerebellar excitatory synapses. Nat. Neurosci. 2001, 4, 1207–1216. [Google Scholar] [CrossRef]
- Tabata, T.; Araishi, K.; Hashimoto, K.; Hashimotodani, Y.; van der Putten, H.; Bettler, B.; Kano, M. Ca 2+ activity at GABA B receptors constitutively promotes metabotropic glutamate signaling in the absence of GABA. Proc. Natl. Acad. Sci. USA 2004, 101, 16952–16957. [Google Scholar] [CrossRef]
- Kamikubo, Y.; Tabata, T.; Kakizawa, S.; Kawakami, D.; Watanabe, M.; Ogura, A.; Iino, M.; Kano, M. Postsynaptic GABAB receptor signalling enhances LTD in mouse cerebellar Purkinje cells. J. Physiol. 2007, 585, 549–563. [Google Scholar] [CrossRef]
- Rives, M.-L.; Vol, C.; Fukazawa, Y.; Tinel, N.; Trinquet, E.; Ayoub, M.A.; Shigemoto, R.; Pin, J.-P.; Prézeau, L. Crosstalk between GABAB and mGlu1a receptors reveals new insight into GPCR signal integration. EMBO J. 2009, 28, 2195–2208. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, M.E.; Isope, P.; Miyazaki, T.; Nakaya, T.; Garcia, E.; Feltz, A.; Schneider, T.; Hescheler, J.; Kano, M.; Sakimura, K.; et al. Functional Coupling between mGluR1 and Cav3.1 T-Type Calcium Channels Contributes to Parallel Fiber-Induced Fast Calcium Signaling within Purkinje Cell Dendritic Spines. J. Neurosci. 2009, 29, 9668–9682. [Google Scholar] [CrossRef] [PubMed]
- Canepari, M.; Ogden, D. Kinetic, pharmacological and activity-dependent separation of two Ca 2+ signalling pathways mediated by type 1 metabotropic glutamate receptors in rat Purkinje neurones. J. Physiol. 2006, 573, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Netzeband, J.G.; Gruol, D.L. mGluR1 agonists elicit a Ca2+ signal and membrane hyperpolarization mediated by apamin-sensitive potassium channels in immature rat purkinje neurons. J. Neurosci. Res. 2008, 86, 293–305. [Google Scholar] [CrossRef]
- Bhandari, P.; Vandael, D.; Fernández-Fernández, D.; Fritzius, T.; Kleindienst, D.; Önal, C.; Montanaro, J.; Gassmann, M.; Jonas, P.; Kulik, A.; et al. GABAB receptor auxiliary subunits modulate Cav2.3-mediated release from medial habenula terminals. Elife 2021, 10, e68274. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coverage (%) | Unique Peptides 1 (Number) | Peptides (Number) |
|---|---|---|
| 35.11 | 11 | 33 |
| 31.61 | 7 | 29 |
| 43.20 | 10 | 34 |
| 49.54 | 11 | 39 |
| 46.79 | 11 | 37 |
| 31.61 | 8 | 27 |
| Protein ID | Protein Name | Gene ID | Unique Peptides (Number) | NCBI ID |
|---|---|---|---|---|
| P97772 | metabotropic glutamate receptor 1 | Grm1 | 48 | NP_058672.1 |
| P18872 | guanine nucleotide-binding protein G(o) subunit α | Gnao1 | 11 | NP_034438.1 |
| P62874 | guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit β-1 | Gnb1 | 6 | NP_001153488.1 |
| Q8BW86 | ρ guanine nucleotide exchange factor 33 | Arhgef33 | 14 | NP_001138924.1 |
| Q6PIC6 | sodium/potassium-transporting ATPase subunit α -3 | Atp1a3 | 26 | NP_001361556.1 |
| Q6PIE5 | sodium/potassium-transporting ATPase subunit α-2 | Atp1a2 | 14 | NP_848492.1 |
| P14094 | sodium/potassium-transporting ATPase subunit β-1 | Atp1b1 | 7 | NP_033851.1 |
| Q91V14 | solute carrier family 12 member 5 | Slc12a5 | 29 | NP_001342409.1 |
| P56564 | excitatory amino acid transporter 1 | Slc1a3 | 6 | NP_683740.1 |
| O35544 | excitatory amino acid transporter 4 | Slc1a6 | 7 | NP_033226.1 |
| Q61625 | glutamate receptor ionotropic, δ-2 | Grid2 | 20 | NP_032193.1 |
| Q99JP6 | homer protein homolog 3 | Homer3 | 14 | NP_001139625.1 |
| P16330 | 2′,3′-cyclic-nucleotide 3′-phosphodiesterase | Cnp | 10 | NP_001139790.1 |
| Q91VR2 | ATP synthase subunit γ, mitochondrial | Atp5c1 | 6 | NP_065640.2 |
| Q8VEM8 | phosphate carrier protein, mitochondrial | Slc25a3 | 5 | NP_598429.1 |
| P48962 | ADP/ATP translocase 1 | Slc25a4 | 3 | NP_031476.3 |
| P61982 | 14-3-3 protein γ | Ywhag | 7 | NP_061359.2 |
| P68254 | 14-3-3 protein θ | Ywhaq | 8 | NP_035869.1 |
| P11881 | inositol 1,4,5-trisphosphate receptor type 1 | Itpr1 | 62 | NP_034715.3 |
| Q7TNC9 | inositol polyphosphate-5-phosphatase A | Inpp5a | 10 | NP_898967.2 |
| P63318 | protein kinase C γ type | Prkcg | 16 | NP_035232.1 |
| P28652 | calcium/calmodulin-dependent protein kinase type II subunit β | Camk2b | 4 | NP_001167524.1 |
| P35802 | neuronal membrane glycoprotein M6-a | Gpm6a | 2 | NP_705809.1 |
| P46097 | synaptotagmin-2 | Syt2 | 4 | NP_001342655.1 |
| Q6WVG3 | BTB/POZ domain-containing protein KCTD12 | Kctd12 | 9 | NP_808383.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansouri, M.; Kremser, L.; Nguyen, T.-P.; Kasugai, Y.; Caberlotto, L.; Gassmann, M.; Sarg, B.; Lindner, H.; Bettler, B.; Carboni, L.; et al. Protein Networks Associated with Native Metabotropic Glutamate 1 Receptors (mGlu1) in the Mouse Cerebellum. Cells 2023, 12, 1325. https://doi.org/10.3390/cells12091325
Mansouri M, Kremser L, Nguyen T-P, Kasugai Y, Caberlotto L, Gassmann M, Sarg B, Lindner H, Bettler B, Carboni L, et al. Protein Networks Associated with Native Metabotropic Glutamate 1 Receptors (mGlu1) in the Mouse Cerebellum. Cells. 2023; 12(9):1325. https://doi.org/10.3390/cells12091325
Chicago/Turabian StyleMansouri, Mahnaz, Leopold Kremser, Thanh-Phuong Nguyen, Yu Kasugai, Laura Caberlotto, Martin Gassmann, Bettina Sarg, Herbert Lindner, Bernhard Bettler, Lucia Carboni, and et al. 2023. "Protein Networks Associated with Native Metabotropic Glutamate 1 Receptors (mGlu1) in the Mouse Cerebellum" Cells 12, no. 9: 1325. https://doi.org/10.3390/cells12091325
APA StyleMansouri, M., Kremser, L., Nguyen, T.-P., Kasugai, Y., Caberlotto, L., Gassmann, M., Sarg, B., Lindner, H., Bettler, B., Carboni, L., & Ferraguti, F. (2023). Protein Networks Associated with Native Metabotropic Glutamate 1 Receptors (mGlu1) in the Mouse Cerebellum. Cells, 12(9), 1325. https://doi.org/10.3390/cells12091325