Liposomal Delivery of miR-34b-5p Induced Cancer Cell Death in Thyroid Carcinoma

 and
and 
Abstract
:1. Introduction
2. Material and Method
2.1. Cell Culture
2.2. HFDM Formulated miRNA-Entrapped PEGylated Lipid Particle
2.3. Nanoparticle Characterization
2.4. Liposome-Loaded miR-34b Transfection
2.5. Quantification of miR-34b Expression
2.6. Western Blot Analysis for VEGF-A in Anaplastic Thyroid Carcinoma Cells
2.7. Enzyme-Linked Immunosorbent (ELISA) Assay
2.8. Cell Proliferation Assay
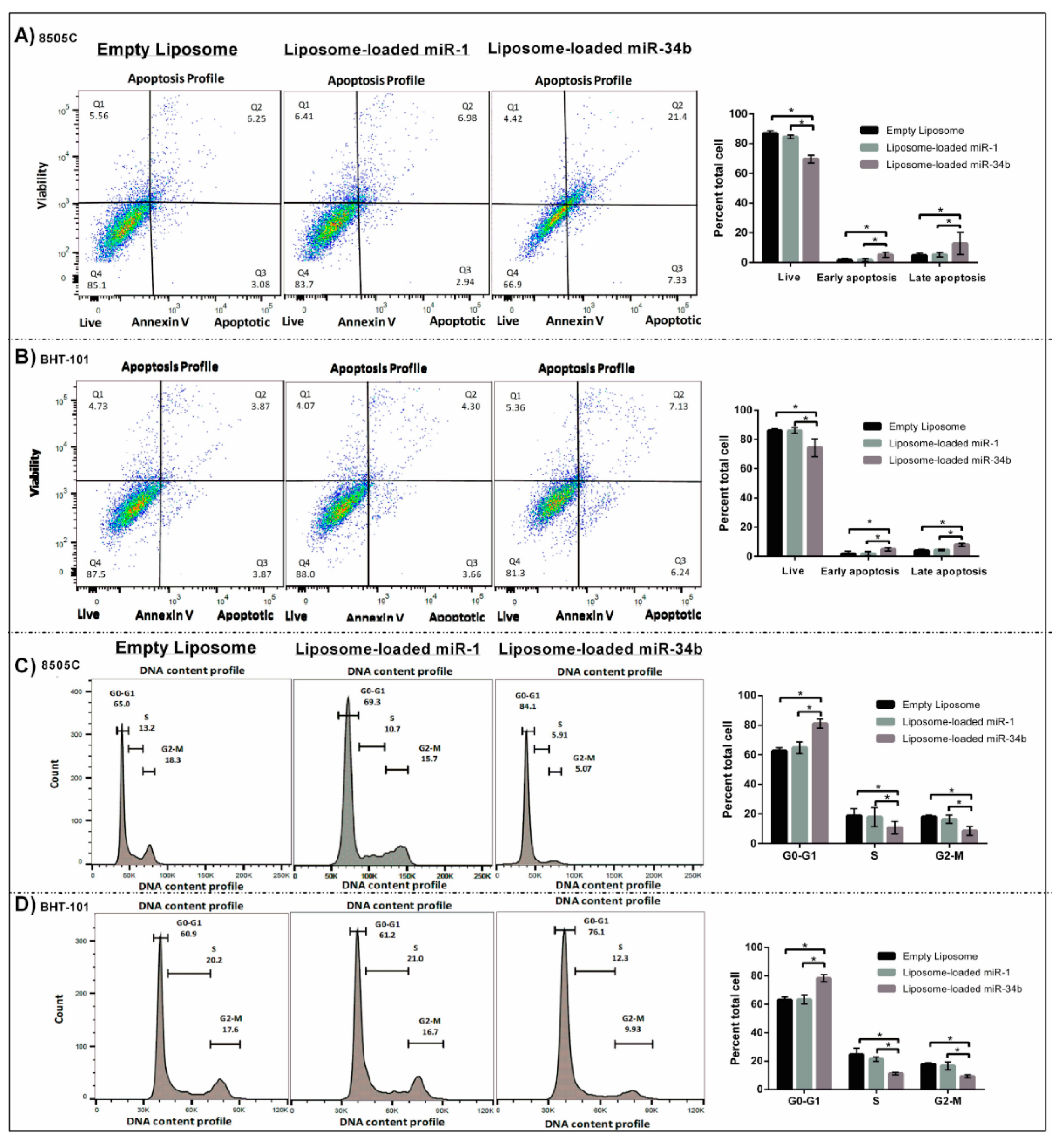
2.9. FACS Analysis for Cell Cycle Distribution
2.10. Quantitative Apoptosis Assay
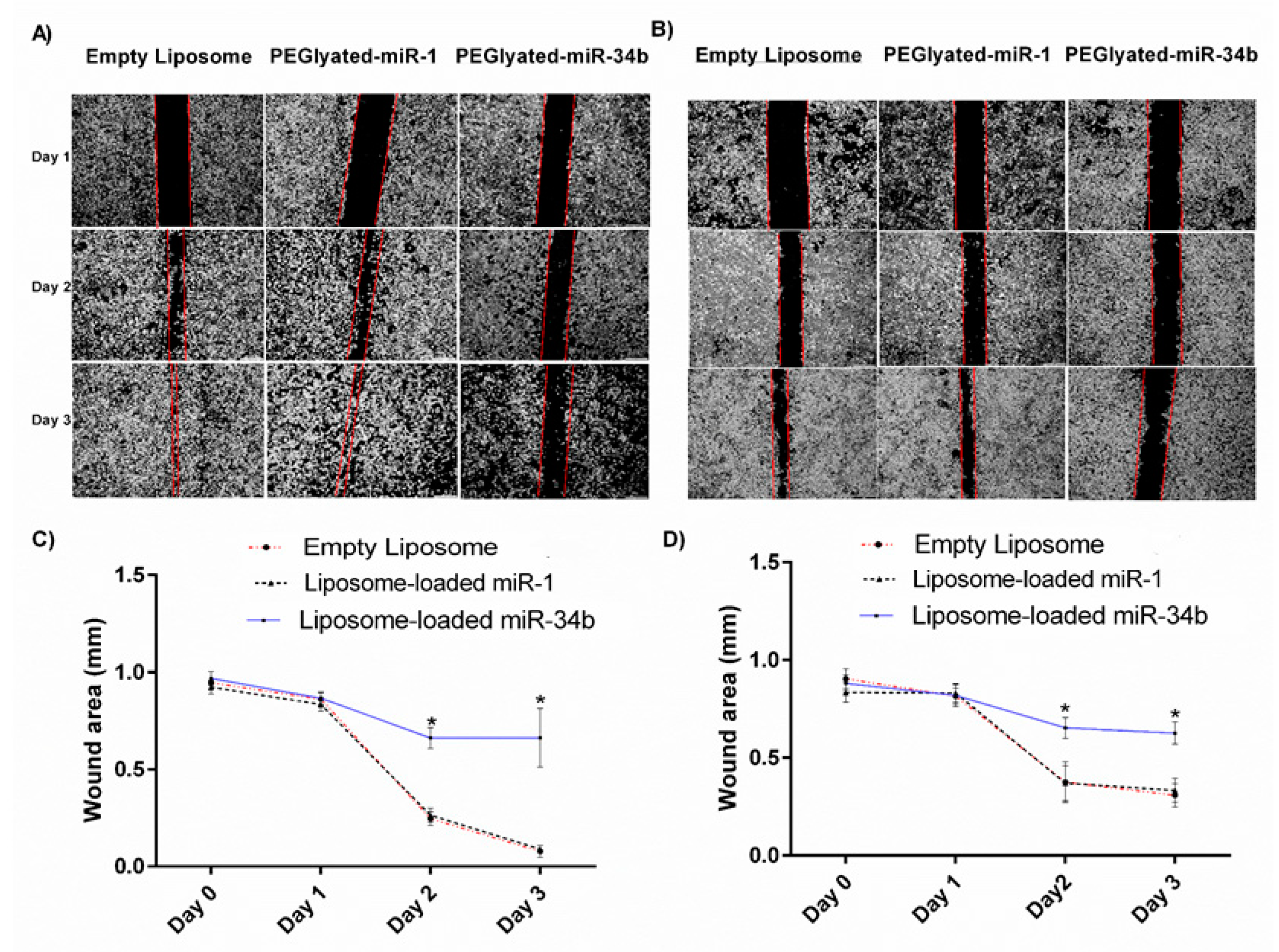
2.11. Wound Healing Assay
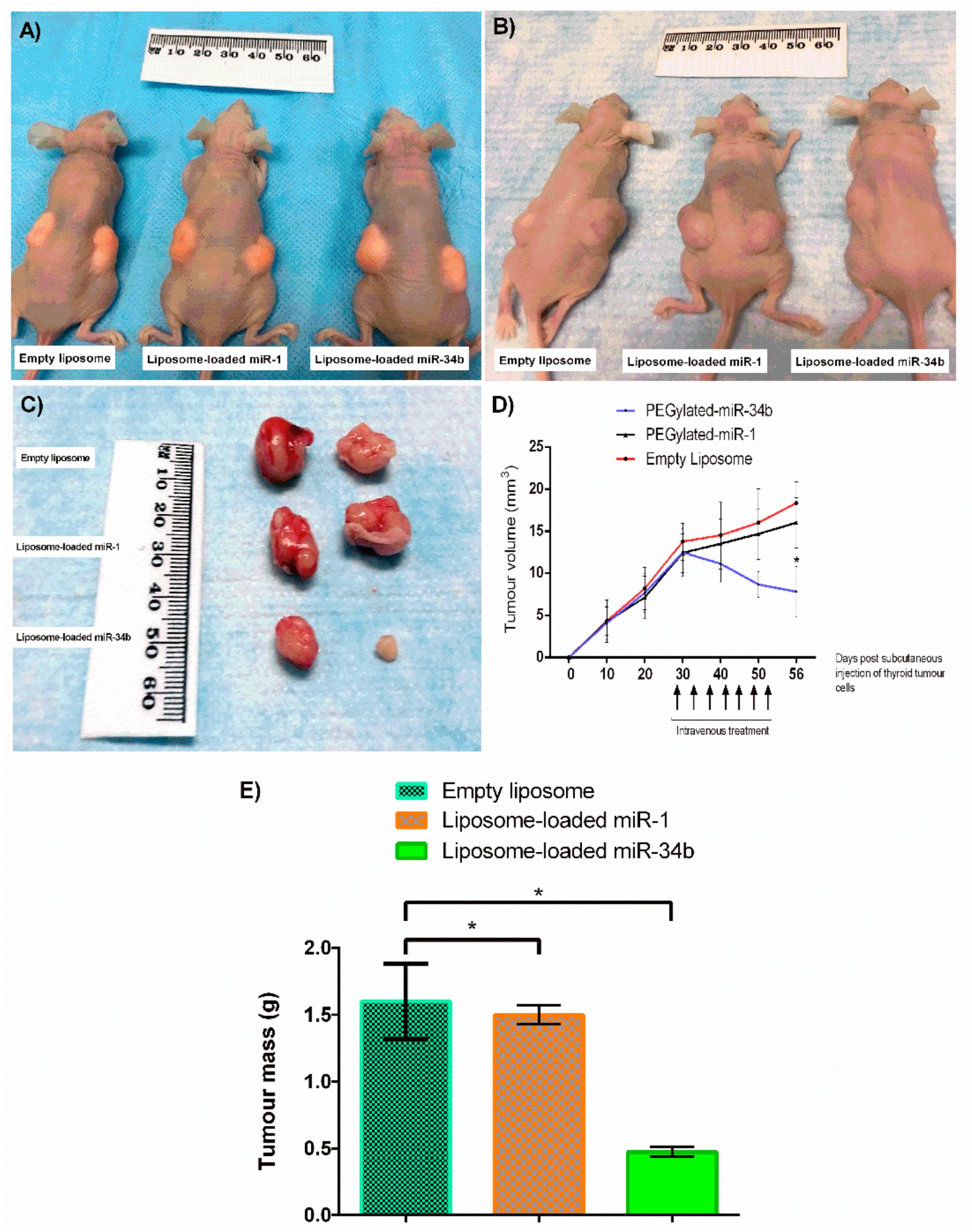
In Vivo Study
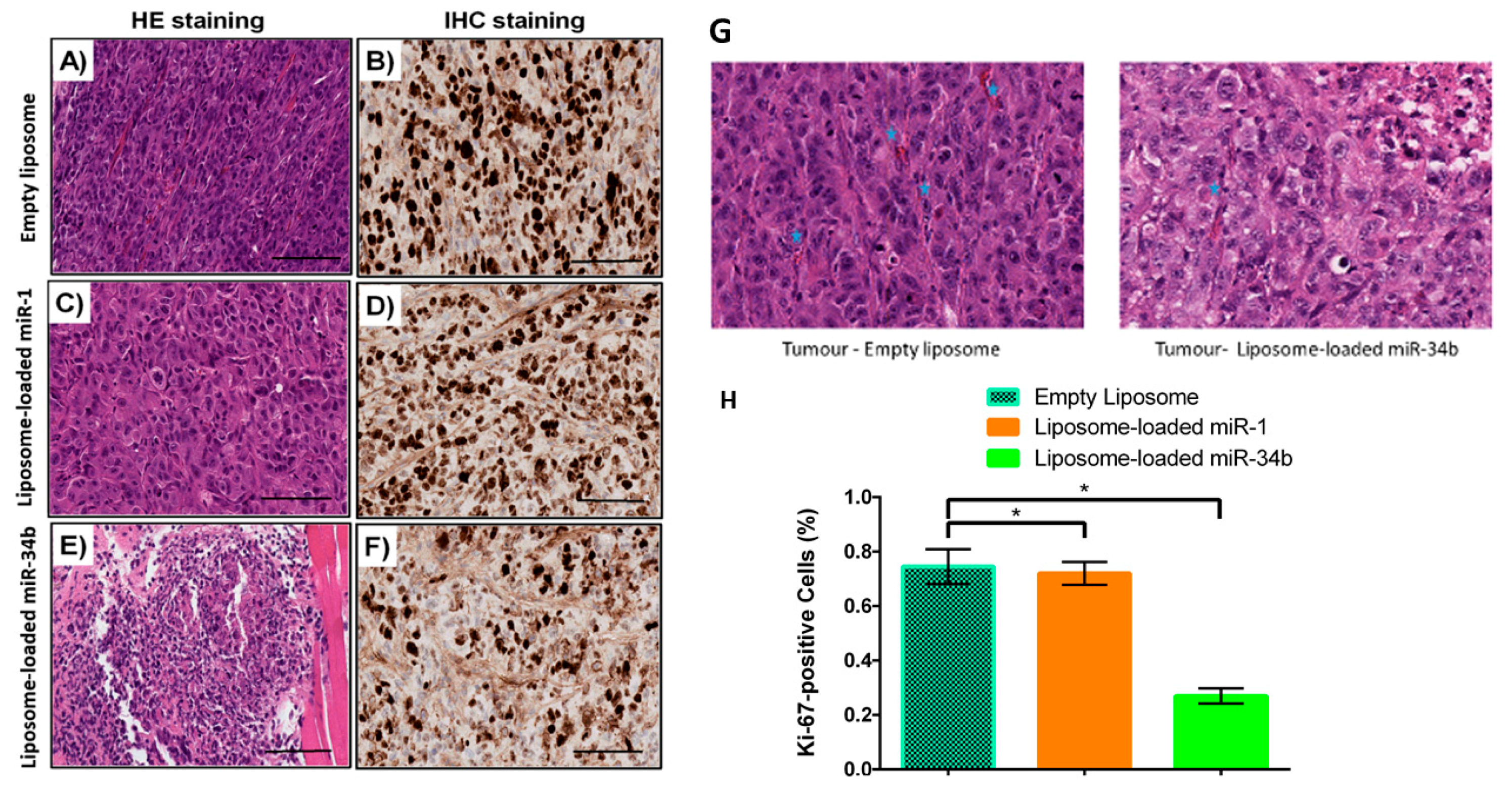
2.12. Histological Analysis and Proliferative Marker
2.13. miR-34b and VEGF-A mRNA Studies in Mice Xenografts
2.14. Western Blot Analysis for VEGF-A in Mice Xenografts
2.15. Data Analysis
3. Results
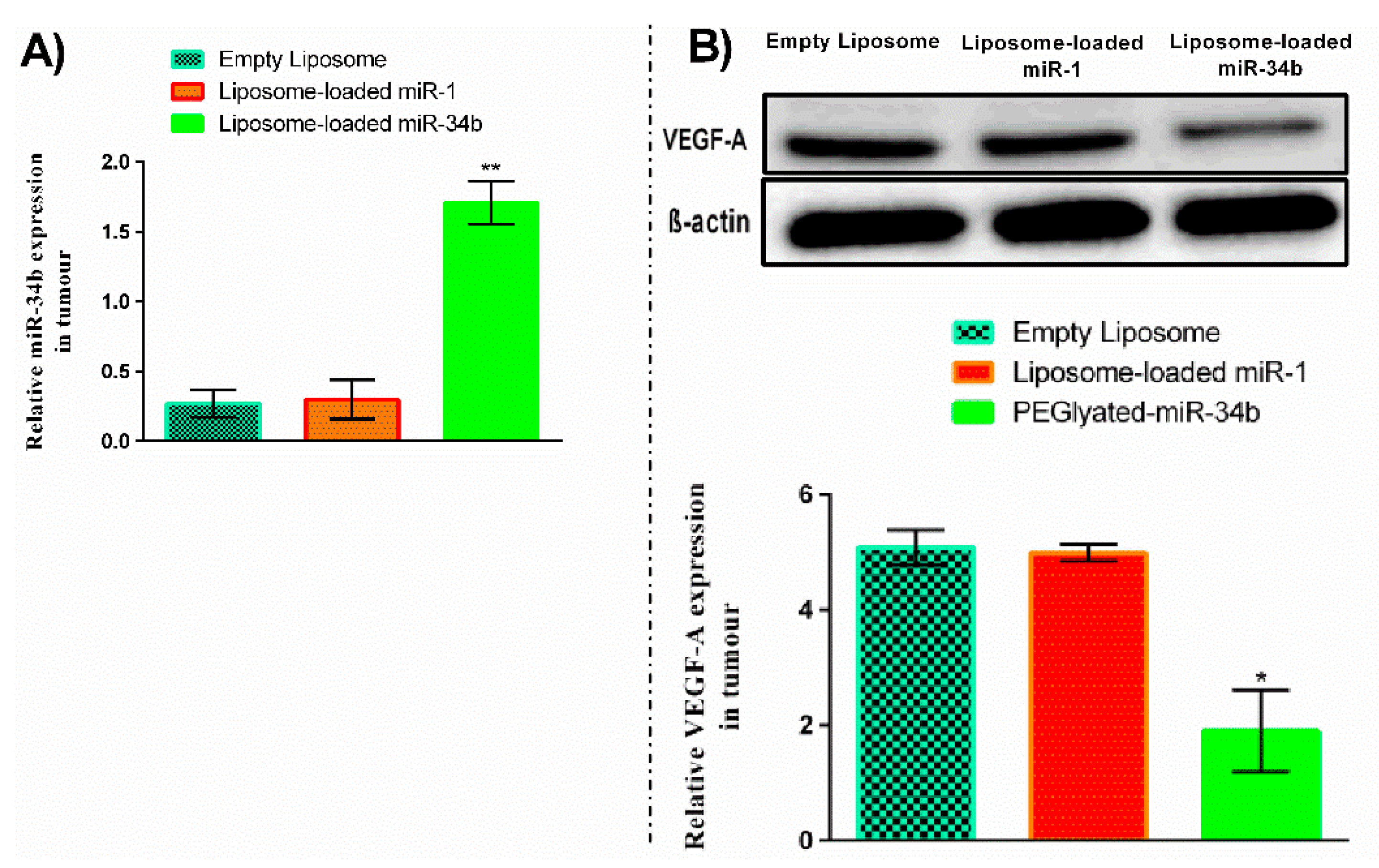
3.1. Characterization of Lipid Nanoparticles Entrapped miR-34b
3.2. Liposome-Loaded miR-34b Overexpression Caused Downregulation of VEGF-A Protein
3.3. Tumour Suppressor Properties of Liposome-Loaded miR-34b In Vitro
3.4. The Suppressive Role of miR-34b in Apoptosis and Cell Cycle Distribution
3.5. In vivo Confirmation of miR-34b Tumour Suppressor Properties
4. Discussion
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
Abbreviations
References
- Lam, A.K. Pathology of endocrine tumors update: World Health Organization new classification 2017-other thyroid tumors. AJSP Rev. Rep. 2017, 22, 209–216. [Google Scholar]
- Lam, A.K.; Lo, C.Y.; Lam, K.S. Papillary carcinoma of thyroid: A 30-yr clinicopathological review of the histological variants. Endocr. Pathol. 2005, 16, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.Y.; Lo, C.Y.; Chan, K.W. Insular and anaplastic carcinoma of the thyroid: A 45-year comparative study at a single institution and a review of the significance of p53 and p21. Ann. Surg. 2000, 231, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.Y.; Lam, K.Y.; Wan, K.Y. Anaplastic carcinoma of the thyroid. Am. J. Surg. 1999, 177, 337–339. [Google Scholar] [CrossRef]
- Rahman, M.A.; Salajegheh, A.; Smith, R.A. MicroRNA-126 suppresses proliferation of undifferentiated (BRAF(V600E) and BRAF(WT)) thyroid carcinoma through targeting PIK3R2 gene and repressing PI3K-AKT proliferation-survival signalling pathway. Exp. Cell. Res. 2015, 339, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Salajegheh, A.; Vosgha, H.; Md Rahman, A. Modulatory role of miR-205 in angiogenesis and progression of thyroid cancer. J. Mol. Endocrinol. 2015, 55, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Vosgha, H.; Ariana, A.; Smith, R.A. miR-205 targets angiogenesis and EMT concurrently in anaplastic thyroid carcinoma. Endocr. Relat. Cancer. 2018, 25, 323–337. [Google Scholar] [CrossRef]
- Maroof, H.; Salajegheh, A.; Smith, R.A. MicroRNA-34 family, mechanisms of action in cancer: A review. Curr. Cancer Drug Target. 2014, 14, 737–751. [Google Scholar] [CrossRef]
- Maroof, H.; Salajegheh, A.; Smith, R.A. Role of microRNA-34 family in cancer with particular reference to cancer angiogenesis. Exp. Mol. Pathol. 2014, 97, 298–304. [Google Scholar] [CrossRef]
- Maroof, H.; Islam, F.; Ariana, A. The roles of microRNA-34b-5p in angiogenesis of thyroid carcinoma. Endocrine 2017, 58, 153–166. [Google Scholar] [CrossRef]
- Yu, X.M.; Lo, C.Y.; Lam, A.K. The potential clinical relevance of serum vascular endothelial growth factor (VEGF) and VEGF-C in recurrent papillary thyroid carcinoma. Surgery 2008, 144, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.M.; Lo, C.Y.; Lam, A.K. Serum vascular endothelial growth factor C correlates with lymph node metastases and high-risk tumor profiles in papillary thyroid carcinoma. Ann. Surg. 2008, 247, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.M.; Lo, C.Y.; Chan, W.F. Increased expression of vascular endothelial growth factor C in papillary thyroid carcinoma correlates with cervical lymph node metastases. Clin. Cancer Res. 2005, 11, 8063–8069. [Google Scholar] [CrossRef] [PubMed]
- Salajegheh, A.; Vosgha, H.; Rahman, M.A. Interactive role of miR-126 on VEGF-A and progression of papillary and undifferentiated thyroid carcinoma. Hum. Pathol. 2016, 51, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Salajegheh, A.; Pakneshan, S.; Rahman, A. Co-regulatory potential of vascular endothelial growth factor-A and vascular endothelial growth factor-C in thyroid carcinoma. Hum. Pathol. 2013, 44, 2204–2212. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M.; Bernstein, E.; Beach, D. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature 2000, 404, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Judge, A.D.; Robbins, M.; Tavakoli, I. Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J. Clin. Investig. 2009, 119, 661–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.D.; Chen, Y.C.; Hackett, M.J. Tumor-targeted delivery of siRNA by self-assembled nanoparticles. Mol. Ther. 2008, 16, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Yagi, N.; Manabe, I.; Tottori, T. A nanoparticle system specifically designed to deliver short interfering RNA inhibits tumor growth in vivo. Cancer Res. 2009, 69, 6531–6538. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Putral, L.N.; Liang, M. Development of a novel method for formulating stable siRNA-loaded lipid particles for in vivo use. Pharm. Res. 2009, 26, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Gantier, M.P.; Tong, S.; Behlke, M.A. Rational design of immunostimulatory siRNAs. Mol. Ther. 2010, 18, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Singhania, A.; Wu, S.Y.; McMillan, N.A. Effective Delivery of PEGylated siRNA-containing lipoplexes to extraperitoneal tumours following intraperitoneal administration. J. Drug Deliv. 2011, 2011, 192562. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Singhania, A.; Burgess, M. Systemic delivery of E6/7 siRNA using novel lipidic particles and its application with cisplatin in cervical cancer mouse models. Gene Ther. 2011, 18, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Chang, H.I.; Burgess, M. Vaginal delivery of siRNA using a novel PEGylated lipoplex-entrapped alginate scaffold system. J. Control. Release 2011, 155, 418–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khairuddin, N.; Gantier, M.P.; Blake, S.J. siRNA-induced immunostimulation through TLR7 promotes antitumoral activity against HPV-driven tumors in vivo. Immunol. Cell. Biol. 2012, 90, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Plummer, P.N.; Freeman, R.; Taft, R.J. MicroRNAs regulate tumor angiogenesis modulated by endothelial progenitor cells. Cancer Res. 2013, 73, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.H.; He, Y.; Hasnain, S.Z. MUC13 protects colorectal cancer cells from death by activating the NF-κB pathway and is a potential therapeutic target. Oncogene 2017, 36, 700–713. [Google Scholar] [CrossRef] [PubMed]
- McCaskill, J.; Singhania, R.; Burgess, M. Efficient biodistribution and gene silencing in the lung epithelium via intravenous liposomal delivery of siRNA. Mol. Ther. Nucl. Acids. 2013, 2, e96. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Gopalan, V.; Vider, J. MicroRNA-186-5p overexpression modulates colon cancer growth by repressing the expression of the FAM134B tumour inhibitor. Exp. Cell. Res. 2017, 357, 260–270. [Google Scholar] [CrossRef]
- Islam, F.; Khanam, J.A.; Khatun, M.A. p-menth-1-ene-4,7-diol (EC-1) from Eucalyptus camaldulensis Dhnh. triggers apoptosis and cell cycle changes in Ehrlich ascites carcinoma cells. Phytother. Res. 2015, 29, 573–581. [Google Scholar] [CrossRef]
- Plonowski, A.; Schally, A.V.; Nagy, A. Inhibition of metastatic renal cell carcinomas expressing somatostatin receptors by a targeted cytotoxic analogue of somatostatin AN-238. Cancer Res. 2000, 60, 2996–3001. [Google Scholar] [PubMed]
- Wahab, R.; Gopalan, V.; Islam, F. Cellular expression, in-vitro and in-vivo confirmation of GAEC1 oncogenic properties in colon cancer. Eur. J. Cell. Biol. 2017, 96, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Gopalan, V.; Smith, R.A.; Lam, A.K. Downregulation of microRNA-498 in colorectal cancers and its cellular effects. Exp. Cell. Res. 2015, 330, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Qiao, B.; Cai, J.H.; King-Yin, L.A. MicroRNA-542-3p inhibits oral squamous cell carcinoma progression by inhibiting ILK/TGF-β1/Smad2/3 signaling. Oncotarget 2017, 8, 70761–70776. [Google Scholar] [CrossRef] [PubMed]
- Lobry, C.; Oh, P.; Aifantis, I. Oncogenic and tumor suppressor functions of Notch in cancer: It’s NOTCH what you think. J. Exp. Med. 2011, 26, 1931–1935. [Google Scholar] [CrossRef] [PubMed]
- Anai, S.; Sakamoto, N.; Sakai, Y. Dual targeting of Bcl-2 and VEGF: A potential strategy to improve therapy for prostate cancer. Urol. Oncol. 2011, 29, 421–429. [Google Scholar] [CrossRef]
- Ji, Q.; Mu, D. MicroRNAs and lung cancers: From pathogenesis to clinical implications. Front. Med. 2012, 6, 134–155. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size (nm) a | 135.3 ± 10.80 |
| Polydispersity index | 0.311 ± 0.06 |
| Zeta potential (mV) | 39.16 ± 0.451 |
| miR-34b-5p entrapment efficiency (%) | 96.9 ± 2.18 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maroof, H.; Islam, F.; Dong, L.; Ajjikuttira, P.; Gopalan, V.; McMillan, N.A.J.; Lam, A.K. Liposomal Delivery of miR-34b-5p Induced Cancer Cell Death in Thyroid Carcinoma. Cells 2018, 7, 265. https://doi.org/10.3390/cells7120265
Maroof H, Islam F, Dong L, Ajjikuttira P, Gopalan V, McMillan NAJ, Lam AK. Liposomal Delivery of miR-34b-5p Induced Cancer Cell Death in Thyroid Carcinoma. Cells. 2018; 7(12):265. https://doi.org/10.3390/cells7120265
Chicago/Turabian StyleMaroof, Hamidreza, Farhadul Islam, LanFeng Dong, Prabha Ajjikuttira, Vinod Gopalan, Nigel A.J. McMillan, and Alfred K. Lam. 2018. "Liposomal Delivery of miR-34b-5p Induced Cancer Cell Death in Thyroid Carcinoma" Cells 7, no. 12: 265. https://doi.org/10.3390/cells7120265
APA StyleMaroof, H., Islam, F., Dong, L., Ajjikuttira, P., Gopalan, V., McMillan, N. A. J., & Lam, A. K. (2018). Liposomal Delivery of miR-34b-5p Induced Cancer Cell Death in Thyroid Carcinoma. Cells, 7(12), 265. https://doi.org/10.3390/cells7120265