Phosphatidylethanolamine-Binding Protein 1 Ameliorates Ischemia-Induced Inflammation and Neuronal Damage in the Rabbit Spinal Cord

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
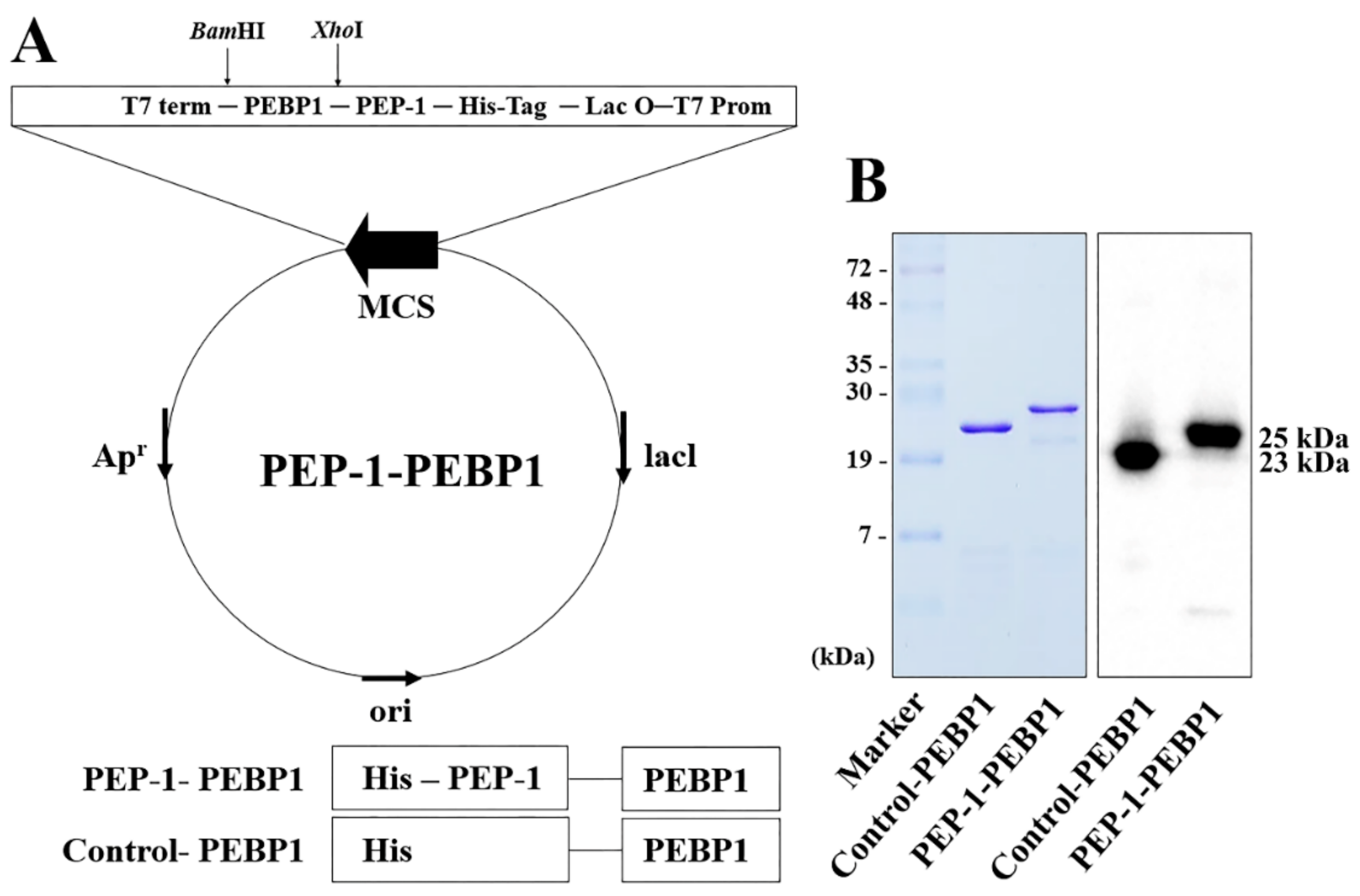
2.1. PEP-1-PEBP1 Fusion Protein Construction and Assessment Its Efficiency
2.1.1. Cell Preparation
2.1.2. Construction of PEP-1-PEBP1
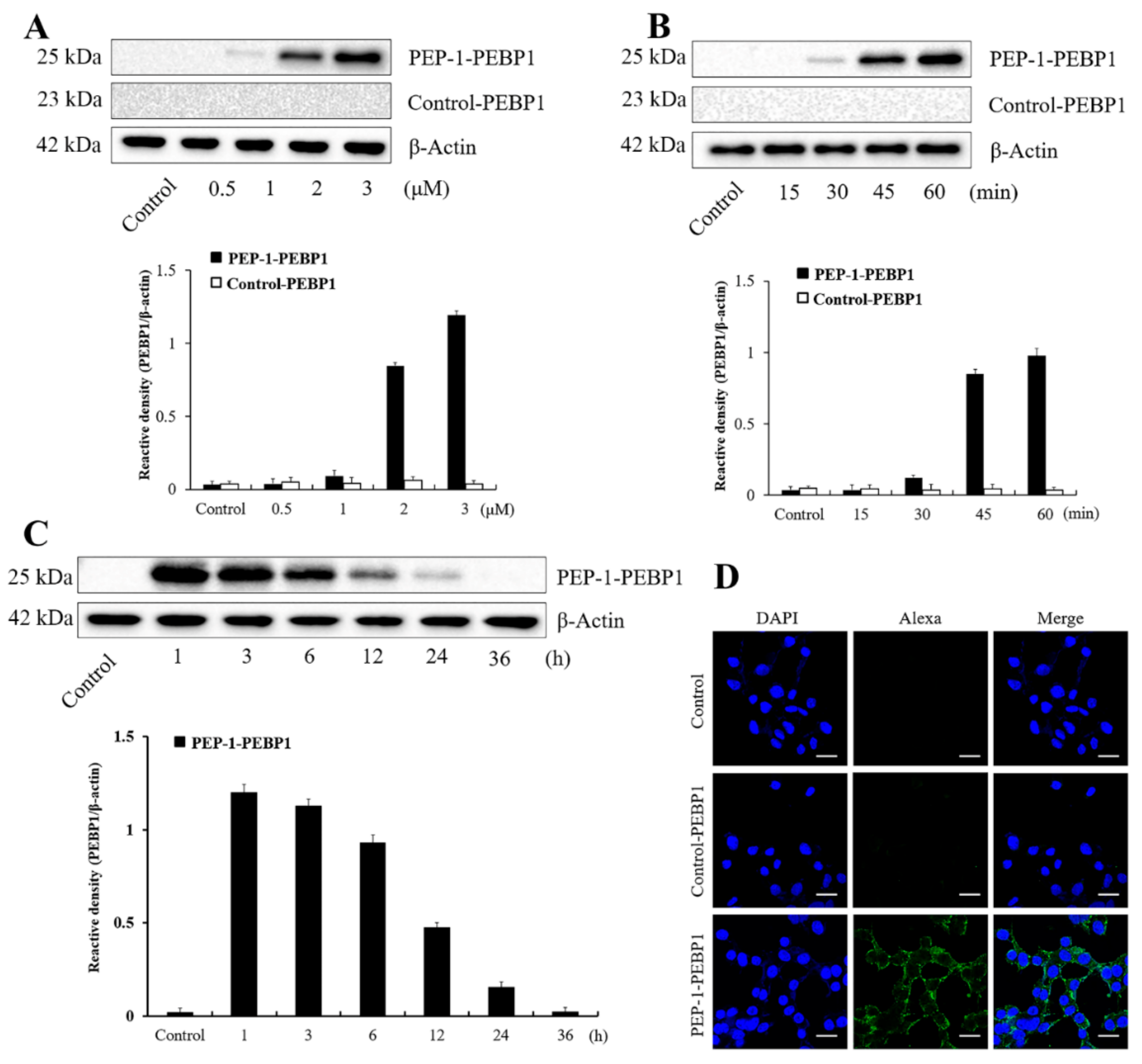
2.1.3. Transduction of PEP-1-PEBP1 Proteins into NSC34 Cells
2.1.4. Intracellular Location of PEP-1-PEBP1
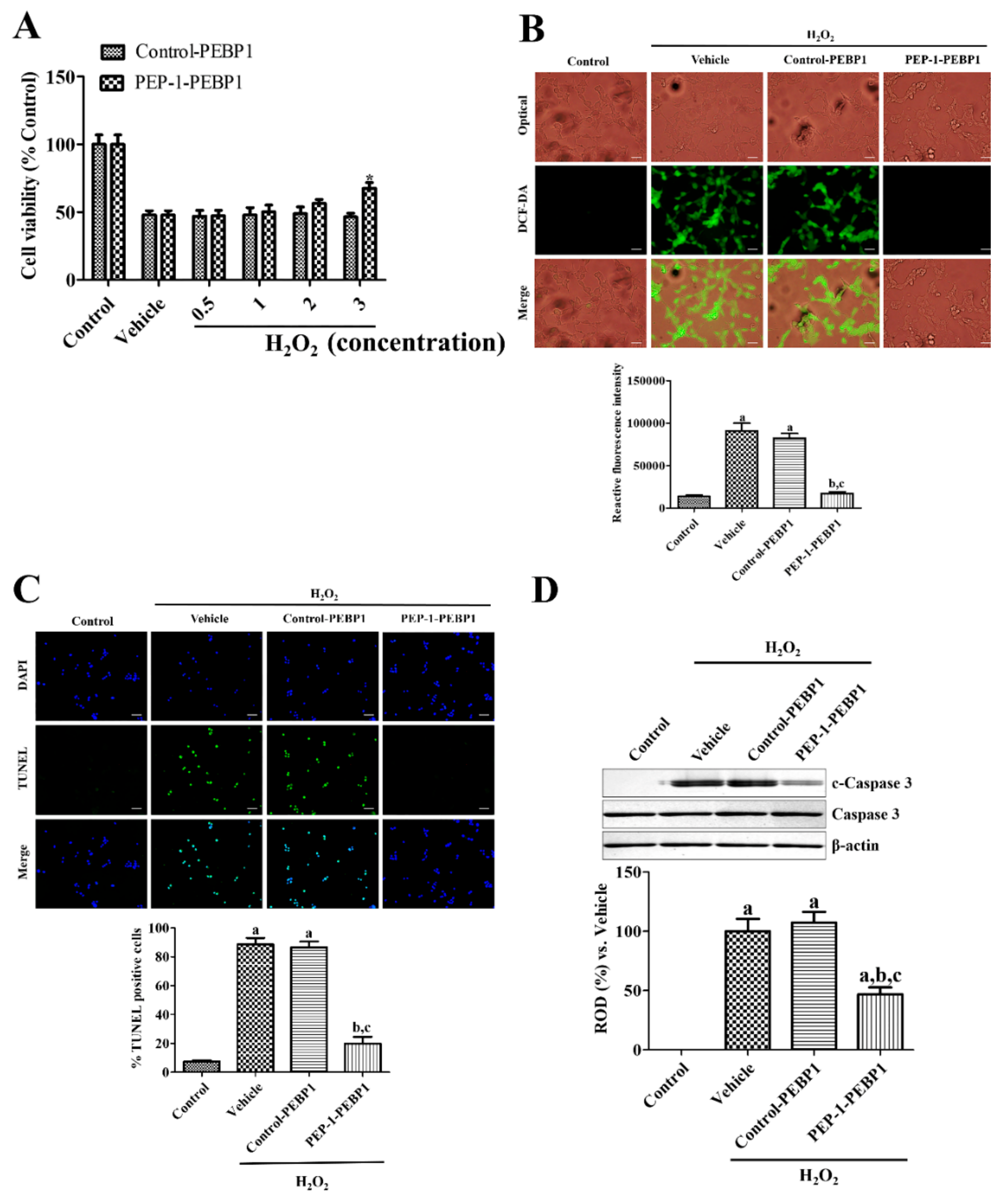
2.2. Protective Effects of PEP-1-PEBP1 against Oxidative Stress in NSC34 Cells
2.2.1. DNA Damage and Oxidative Stress
2.2.2. Measurement of Cell Viability
2.2.3. Western Blot Analysis
2.3. In vivo Neuroprotective Action of PEP-1-PEBP1 against Spinal Cord Ischemia
2.3.1. Experimental Animals
2.3.2. Determination of Effective PEP-1-PEBP1 Concentration against Ischemic Damage
2.3.3. Ischemic Surgery
2.3.4. Monitoring of Physiological Parameters
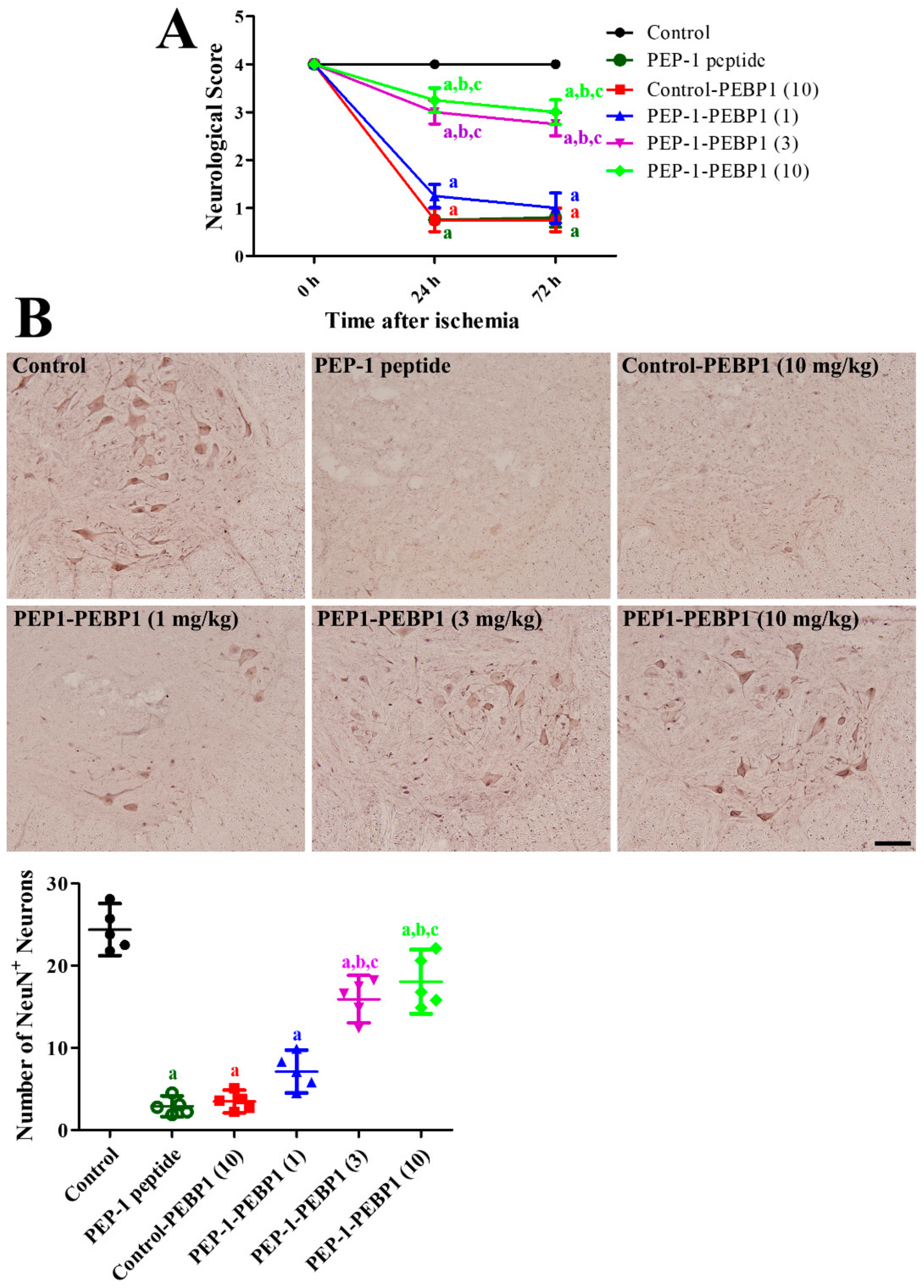
2.3.5. Neurological Assessment
2.3.6. Detection of Neuronal Survival and Cell Death
2.3.7. Biochemical Assessments in Spinal Cord Tissue
2.4. Statistical Analysis
3. Results
3.1. Production and Delivery of PEP-1-PEBP1 Protein into NSC34 Cells
3.2. Effects of PEP-1-PEBP1 on Oxidative Damage in NSC34 Cells
3.3. Effects of PEP-1-PEBP1 on Physiological Data against Spinal Cord Ischemia
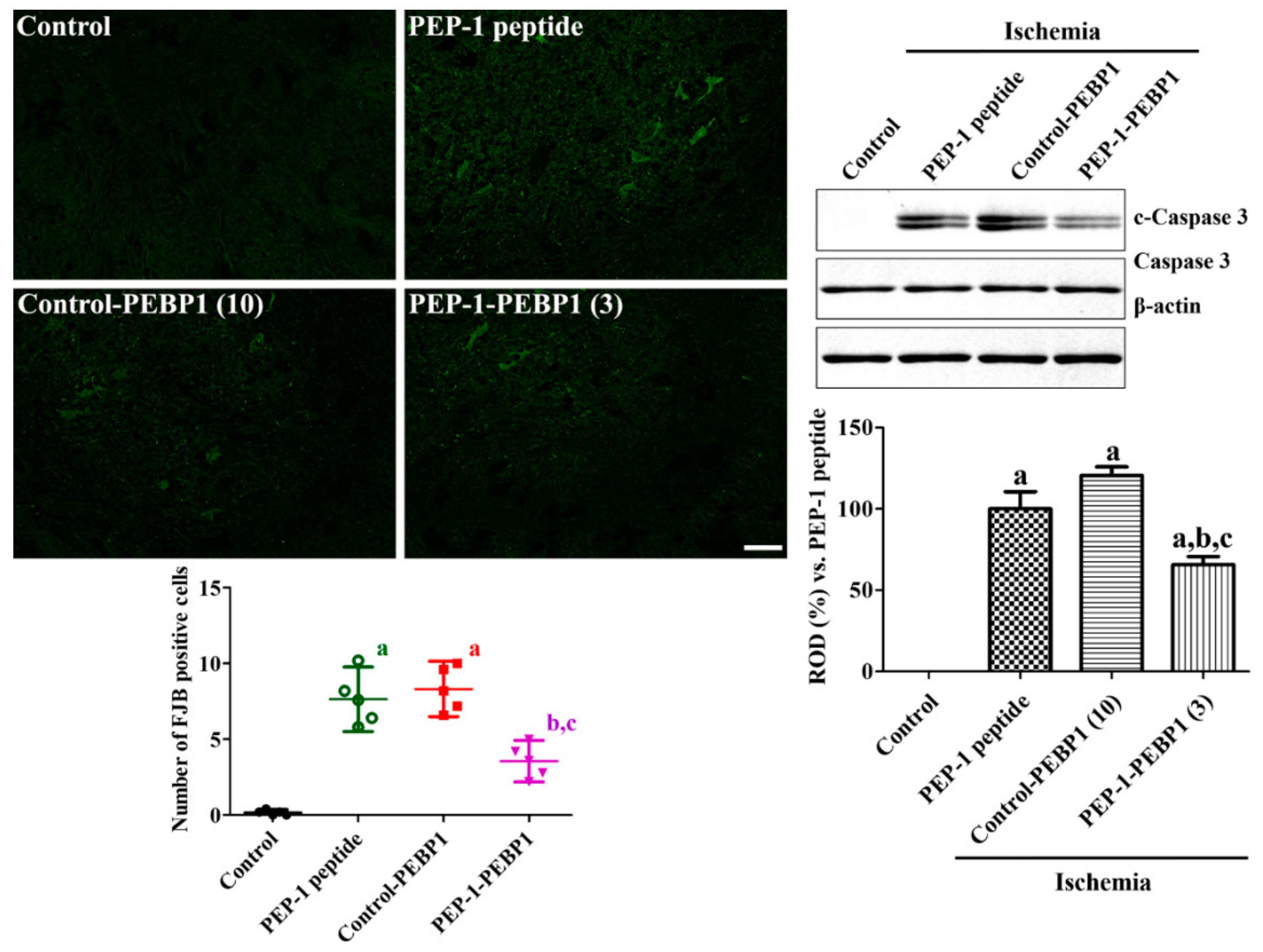
3.4. Effects of PEP-1-PEBP1 on Neuronal Death against Spinal Cord Ischemia
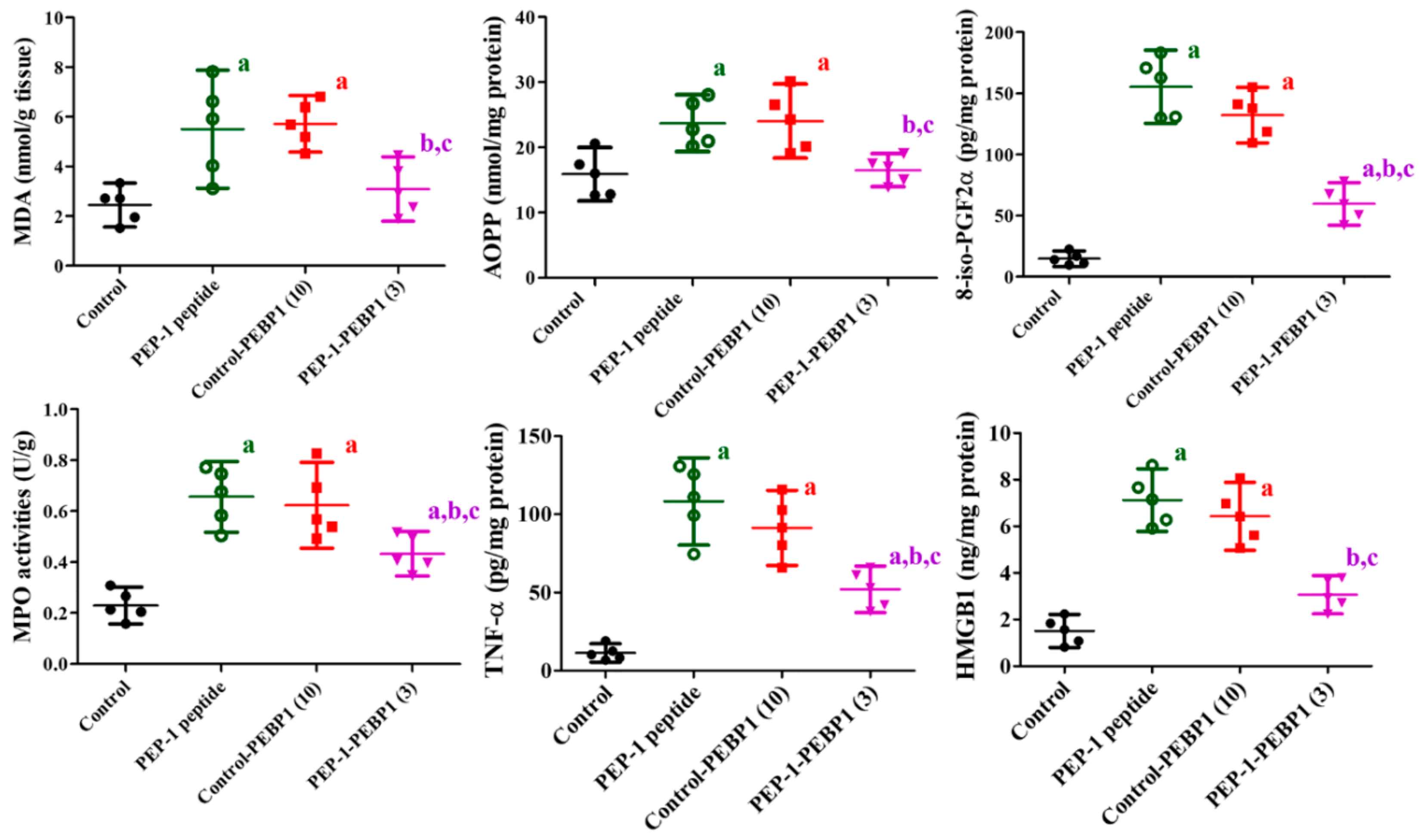
3.5. Effects of PEP-1-PEBP1 on Oxidative Stress and Inflammation after Spinal Cord Ischemia
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Yamauchi, T.; Takano, H.; Nishimura, M.; Matsumiya, G.; Sawa, Y. Paraplegia and paraparesis after descending thoracic aortic aneurysm repair: A risk factor analysis. Ann. Thorac. Cardiovasc. Surg. 2006, 12, 179–183. [Google Scholar]
- Disch, A.C.; Schaser, K.D.; Melcher, I.; Feraboli, F.; Schmoelz, W.; Druschel, C.; Luzzati, A. Oncosurgical results of multilevel thoracolumbar en-bloc spondylectomy and reconstruction with a carbon composite vertebral body replacement system. Spine 2011, 36, e647–e655. [Google Scholar] [CrossRef]
- Luzzati, A.D.; Shah, S.P.; Gagliano, F.S.; Perrucchini, G.G.; Fontanella, W.; Alloisio, M. Four- and five- level en bloc spondylectomy for malignant spinal tumors. Spine 2014, 39, e129–e139. [Google Scholar] [CrossRef]
- Etz, D.C.; Luehr, M.; Aspern, K.V.; Misfeld, M.; Gudehus, S.; Ender, J.; Koelbel, T.; Debus, E.S.; Mohr, F.W. Spinal cord ischemia in open and endovascular thoracoabdominal aortic aneurysm repair: New concepts. J. Cardiovasc. Surg. 2014, 55, 159–168. [Google Scholar]
- Mazensky, D.; Radonak, J.; Danko, J.; Petrovova, E.; Frankovicova, M. Anatomical study of blood supply to the spinal cord in the rabbit. Spinal Cord 2011, 49, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Plata-Salamán, C.R.; Kelly, G.; Agresta, C.; Taylor, K.; Salzman, S.K. Interleukin-1 beta enhances spinal cord blood flow after intrathecal administration in the normal rat. Am. J. Physiol. 1995, 269, R1032–R1037. [Google Scholar]
- Akuzawa, S.; Kazui, T.; Shi, E.; Yamashita, K.; Bashar, A.H.; Terada, H. Interleukin-1 receptor antagonist attenuates the severity of spinal cord ischemic injury in rabbits. J. Vasc. Surg. 2008, 48, 694–700. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Zhu, H.; Li, J.; Wang, X.; Misra, H.; Li, Y. Oxidative stress in spinal cord injury and antioxidant-based intervention. Spinal Cord 2012, 50, 264–274. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Hyde, J.S. Concentration of oxygen in lipid bilayers using a spin-label method. Biophys. J. 1983, 41, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Ojika, K.; Kojima, S.; Ueki, Y.; Fukushima, N.; Hayashi, K.; Yamamoto, M. Purification and structural analysis of hippocampal cholinergic neurostimulating peptide. Brain Res. 1992, 572, 164–171. [Google Scholar] [CrossRef]
- Maki, M.; Matsukawa, N.; Yuasa, H.; Otsuka, Y.; Yamamoto, T.; Akatsu, H.; Okamoto, T.; Ueda, R.; Ojika, K. Decreased expression of hippocampal cholinergic neurostimulating peptide precursor protein mRNA in the hippocampus in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2002, 61, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, R.E., Jr.; Maurer, M.H.; Hunzinger, C.; Lewicka, S.; Buergers, H.F.; Kalenka, A.; Hinkelbein, J.; Broemme, J.O.; Seidler, G.H.; Martin, E.; et al. Reduction in rat phosphatidylethanolamine binding protein-1 (PEBP1) after chronic corticosterone treatment may be paralleled by cognitive impairment: A first study. Stress 2008, 11, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, J.; Rommelspacher, H.; Mühlbauer, E.; Wernicke, C. Raf kinase inhibitor protein enhances neuronal differentiation in human SH-SY5Y cells. Dev. Neurosci. 2010, 32, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Sagisaka, T.; Matsukawa, N.; Toyoda, T.; Uematsu, N.; Kanamori, T.; Wake, H.; Borlongan, C.V.; Ojika, K. Directed neural lineage differentiation of adult hippocampal progenitor cells via modulation of hippocampal cholinergic neurostimulating peptide precursor expression. Brain Res. 2010, 1327, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Yoo, D.Y.; Cho, S.B.; Jung, H.Y.; Kim, W.; Choi, G.M.; Won, M.H.; Kim, D.W.; Hwang, I.K.; Choi, S.Y.; Moon, S.M. Tat-protein disulfide-isomerase A3: A possible candidate for preventing ischemic damage in the spinal cord. Cell Death Dis. 2017, 8, e3075. [Google Scholar] [CrossRef]
- Jung, H.Y.; Cho, S.B.; Kim, W.; Yoo, D.Y.; Won, M.H.; Choi, G.M.; Cho, T.G.; Kim, D.W.; Hwang, I.K.; Choi, S.Y.; et al. Phosphatidylethanolamine-binding protein 1 protects CA1 neurons against ischemic damage via ERK-CREB signaling in Mongolian gerbils. Neurochem. Int. 2018, 118, 265–274. [Google Scholar] [CrossRef]
- Eggett, C.J.; Crosier, S.; Manning, P.; Cookson, M.R.; Menzies, F.M.; McNeil, C.J.; Shaw, P.J. Development and characterisation of a glutamate-sensitive motor neurone cell line. J. Neurochem. 2000, 74, 1895–1902. [Google Scholar] [CrossRef]
- Eum, W.S.; Kim, D.W.; Hwang, I.K.; Yoo, K.Y.; Kang, T.C.; Jang, S.H.; Choi, H.S.; Choi, S.H.; Kim, Y.H.; Kim, S.Y.; et al. In vivo protein transduction: Biologically active intact pep-1-superoxide dismutase fusion protein efficiently protects against ischemic insult. Free Radic. Biol. Med. 2004, 37, 1656–1669. [Google Scholar]
- Jo, H.S.; Kim, D.W.; Shin, M.J.; Cho, S.B.; Park, J.H.; Lee, C.H.; Yeo, E.J.; Choi, Y.J.; Yeo, H.J.; Sohn, E.J.; et al. Tat-HSP22 inhibits oxidative stress-induced hippocampal neuronal cell death by regulation of the mitochondrial pathway. Mol. Brain 2017, 10, 1. [Google Scholar] [CrossRef]
- Yeo, H.J.; Shin, M.J.; Yeo, E.J.; Choi, Y.J.; Kim, D.W.; Kim, D.S.; Eum, W.S.; Choi, S.Y. Tat-CIAPIN1 inhibits hippocampal neuronal cell damage through the MAPK and apoptotic signaling pathways. Free Radic. Biol. Med. 2019, 135, 68–78. [Google Scholar] [CrossRef]
- Kim, W.; Kwon, H.J.; Jung, H.Y.; Yoo, D.Y.; Moon, S.M.; Kim, D.W.; Hwang, I.K. Tat-HSP70 protects neurons from oxidative damage in the NSC34 cells and ischemic damage in the ventral horn of rabbit spinal cord. Neurochem. Int. 2019, 129, 104477. [Google Scholar] [CrossRef] [PubMed]
- Moore, W.M., Jr.; Hollier, L.H. The influence of severity of spinal cord ischemia in the etiology of delayed-onset paraplegia. Ann. Surg. 1991, 213, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Wisselink, W.; Patetsios, P.; Panetta, T.F.; Ramirez, J.A.; Rodino, W.; Kirwin, J.D.; Zikria, B.A. Medium molecular weight pentastarch reduces reperfusion injury by decreasing capillary leak in an animal model of spinal cord ischemia. J. Vasc. Surg. 1998, 27, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Candelario-Jalil, E.; Alvarez, D.; Merino, N.; León, O.S. Delayed treatment with nimesulide reduces measures of oxidative stress following global ischemic brain injury in gerbils. Neurosci. Res. 2003, 47, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.J.; Jung, H.Y.; Hahn, K.R.; Kim, W.; Kim, J.W.; Yoo, D.Y.; Yoon, Y.S.; Hwang, I.K.; Kim, D.W. Bacopa monnieri extract improves novel object recognition, cell proliferation, neuroblast differentiation, brain-derived neurotrophic factor, and phosphorylation of cAMP response element-binding protein in the dentate gyrus. Lab. Anim. Res. 2018, 34, 239–247. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [Green Version]
- George, A.J.; Holsinger, R.M.; McLean, C.A.; Tan, S.S.; Scott, H.S.; Cardamone, T.; Cappai, R.; Masters, C.L.; Li, Q.X. Decreased phosphatidylethanolamine binding protein expression correlates with Aβ accumulation in the Tg2576 mouse model of Alzheimer’s disease. Neurobiol. Aging 2006, 27, 614–623. [Google Scholar] [CrossRef]
- Su, L.; Zhang, R.; Chen, Y.; Zhu, Z.; Ma, C. Raf kinase inhibitor protein attenuates ischemic-induced microglia cell apoptosis and activation through NF-κB pathway. Cell. Physiol. Biochem. 2017, 41, 1125–1134. [Google Scholar] [CrossRef]
- Aluise, C.D.; Robinson, R.A.; Cai, J.; Pierce, W.M.; Markesbery, W.R.; Butterfield, D.A. Redox proteomics analysis of brains from subjects with amnestic mild cognitive impairment compared to brains from subjects with preclinical Alzheimer’s disease: Insights into memory loss in MCI. J. Alzheimers Dis. 2011, 23, 257–269. [Google Scholar] [CrossRef]
- Gatta, V.; D’Aurora, M.; Granzotto, A.; Stuppia, L.; Sensi, S.L. Early and sustained altered expression of aging-related genes in young 3xTg-AD mice. Cell Death Dis. 2014, 5, e1054. [Google Scholar] [CrossRef]
- Tsai, M.C.; Shen, L.F.; Kuo, H.S.; Cheng, H.; Chak, K.F. Involvement of acidic fibroblast growth factor in spinal cord injury repair processes revealed by a proteomics approach. Mol. Cell. Proteomics 2008, 7, 1668–1687. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, S.; Ma, H.; Yang, S.; Liu, Z.; Wu, X.; Wang, S.; Zhang, Y.; Liu, Y. Chronic intermittent hypobaric hypoxia pretreatment ameliorates ischemia-induced cognitive dysfunction through activation of ERK1/2-CREB-BDNF pathway in anesthetized mice. Neurochem. Res. 2017, 42, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Benkler, C.; Ben-Zur, T.; Barhum, Y.; Offen, D. Altered astrocytic response to activation in SOD1G93A mice and its implications on amyotrophic lateral sclerosis pathogenesis. Glia 2013, 61, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Maier, O.; Böhm, J.; Dahm, M.; Brück, S.; Beyer, C.; Johann, S. Differentiated NSC-34 motoneuron-like cells as experimental model for cholinergic neurodegeneration. Neurochem. Int. 2013, 62, 1029–1038. [Google Scholar] [CrossRef]
- Valbuena, G.N.; Rizzardini, M.; Cimini, S.; Siskos, A.P.; Bendotti, C.; Cantoni, L.; Keun, H.C. Metabolomic analysis reveals increased aerobic glycolysis and amino acid deficit in a cellular model of amyotrophic lateral sclerosis. Mol. Neurobiol. 2016, 53, 2222–2240. [Google Scholar] [CrossRef]
- Jung, H.Y.; Kim, D.W.; Yim, H.S.; Yoo, D.Y.; Kim, J.W.; Won, M.H.; Yoon, Y.S.; Choi, S.Y.; Hwang, I.K. Heme oxygenase-1 protects neurons from ischemic damage by upregulating expression of Cu,Zn-superoxide dismutase, catalase, and brain-derived neurotrophic factor in the rabbit spinal cord. Neurochem. Res. 2016, 41, 869–879. [Google Scholar] [CrossRef]
- Seddiqi, N.; Segretain, D.; Bucquoy, S.; Pineau, C.; Jégou, B.; Jollès, P.; Schoentgen, F. Characterization and localization of the rat, mouse and human testicular phosphatidylethanolamine binding protein. Experientia 1996, 52, 101–110. [Google Scholar] [CrossRef]
- Hagan, S.; Al-Mulla, F.; Mallon, E.; Oien, K.; Ferrier, R.; Gusterson, B.; García, J.J.; Kolch, W. Reduction of Raf-1 kinase inhibitor protein expression correlates with breast cancer metastasis. Clin. Cancer Res. 2005, 11, 7392–7397. [Google Scholar] [CrossRef]
- Al-Mulla, F.; Hagan, S.; Behbehani, A.I.; Bitar, M.S.; George, S.S.; Going, J.J.; García, J.J.; Scott, L.; Fyfe, N.; Murray, G.I.; et al. Raf kinase inhibitor protein expression in a survival analysis of colorectal cancer patients. J. Clin. Oncol. 2006, 24, 5672–5679. [Google Scholar] [CrossRef]
- Madji Hounoum, B.; Vourc’h, P.; Felix, R.; Corcia, P.; Patin, F.; Guéguinou, M.; Potier-Cartereau, M.; Vandier, C.; Raoul, C.; Andres, C.R.; et al. NSC-34 motor neuron-like cells are unsuitable as experimental model for glutamate-mediated excitotoxicity. Front. Cell. Neurosci. 2016, 10, 118. [Google Scholar] [CrossRef]
- Al-Mulla, F.; Bitar, M.S.; Al-Maghrebi, M.; Behbehani, A.I.; Al-Ali, W.; Rath, O.; Doyle, B.; Tan, K.Y.; Pitt, A.; Kolch, W. Raf kinase inhibitor protein RKIP enhances signaling by glycogen synthase kinase-3β. Cancer Res. 2011, 71, 1334–1343. [Google Scholar] [CrossRef] [PubMed]
- Li, W.J.; Pan, S.Q.; Zeng, Y.S.; Su, B.G.; Li, S.M.; Ding, Y.; Li, Y.; Ruan, J.W. Ruan, Identification of acupuncture-specific proteins in the process of electro-acupuncture after spinal cord injury. Neurosci. Res. 2010, 67, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, T.; Matsukawa, N.; Sagisaka, T.; Uematsu, N.; Kanamori, T.; Kato, D.; Mizuno, M.; Wake, H.; Hida, H.; Borlongan, C.V.; et al. Suppression of astrocyte lineage in adult hippocampal progenitor cells expressing hippocampal cholinergic neurostimulating peptide precursor in an in vivo ischemic model. Cell Transplant. 2012, 21, 2159–2169. [Google Scholar] [CrossRef] [PubMed]
- Dinc, C.; Iplikcioglu, A.C.; Atabey, C.; Eroglu, A.; Topuz, K.; Ipcioglu, O.; Demirel, D. Comparison of deferoxamine and methylprednisolone: Protective effect of pharmacological agents on lipid peroxidation in spinal cord injury in rats. Spine 2013, 38, e1649–e1655. [Google Scholar] [CrossRef]
- Praticò, D. F2-isoprostanes: Sensitive and specific non-invasive indices of lipid peroxidation in vivo. Atherosclerosis 1999, 147, 1–10. [Google Scholar] [CrossRef]
- Basu, S. Radioimmunoassay of 8-iso-prostaglandin F2alpha: An index for oxidative injury via free radical catalysed lipid peroxidation. Prostaglandins Leukot. Essent. Fatty Acids 1988, 58, 319–325. [Google Scholar] [CrossRef]
- Emmez, H.; Yildirim, Z.; Kale, A.; Tönge, M.; Durdağ, E.; Börcek, A.O.; Uçankuş, L.N.; Doğulu, F.; Kiliç, N.; Baykaner, M.K. Anti-apoptotic and neuroprotective effects of α-lipoic acid on spinal cord ischemia-reperfusion injury in rabbits. Acta Neurochir. 2010, 152, 1591–1600. [Google Scholar] [CrossRef]
- Krawisz, J.E.; Sharon, P.; Stenson, W.F. Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity. Assessment of inflammation in rat and hamster models. Gastroenterology 1984, 87, 1344–1350. [Google Scholar] [CrossRef]
- Chen, X.G.; Hua, F.; Wang, S.G.; Xu, Y.Y.; Yue, H.T. Phosphodiesterase inhibitor ameliorates neuronal injury in spinal cord ischemia/reperfusion injured rat model. Biomed. Pharmacother. 2017, 93, 1213–1219. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Nguyen, D.H. Immunoglobulin G: A potential treatment to attenuate neuroinflammation following spinal cord injury. J. Clin. Immunol. 2010, 30 (Suppl. 1), S109–S112. [Google Scholar] [CrossRef]
- Hirose, K.; Okajima, K.; Taoka, Y.; Uchiba, M.; Tagami, H.; Nakano, K.; Utoh, J.; Okabe, H.; Kitamura, N. Activated protein C reduces the ischemia/reperfusion-induced spinal cord injury in rats by inhibiting neutrophil activation. Ann. Surg. 2010, 232, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, X.; Xue, W.; Xie, K.; Huang, Y.; Chen, H.; Gong, G.; Zeng, Y. Beneficial effects of hydrogen-rich saline against spinal cord ischemia-reperfusion injury in rabbits. Brain Res. 2013, 1517, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Kurt, G.; Yildirim, Z.; Cemil, B.; Celtikci, E.; Kaplanoglu, G.T. Effects of curcumin on acute spinal cord ischemia-reperfusion injury in rabbits. Laboratory investigation. J. Neurosurg. Spine 2014, 20, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.B.; Uchida, K.; Nakajima, H.; Yayama, T.; Hirai, T.; Rodriguez Guerrero, A.; Kobayashi, S.; Ma, W.Y.; Liu, S.Y.; Zhu, P.; et al. High-mobility group box-1 and its receptors contribute to proinflammatory response in the acute phase of spinal cord injury in rats. Spine 2011, 36, 2122–2129. [Google Scholar] [CrossRef]
- Kikuchi, K.; Uchikado, H.; Miura, N.; Morimoto, Y.; Ito, T.; Tancharoen, S.; Miyata, K.; Sakamoto, R.; Kikuchi, C.; Iida, N.; et al. HMGB1 as a therapeutic target in spinal cord injury: A hypothesis for novel therapy development. Exp. Ther. Med. 2012, 2, 767–770. [Google Scholar] [CrossRef]
- Zhu, X.L.; Chen, X.; Wang, W.; Li, X.; Huo, J.; Wang, Y.; Min, Y.Y.; Su, B.X.; Pei, J.M. Electroacupuncture pretreatment attenuates spinal cord ischemia-reperfusion injury via inhibition of high-mobility group box 1 production in a LXA4 receptor-dependent manner. Brain Res. 2017, 1659, 113–120. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Distal MAP (mmHg) | PaCO2 (mmHg) | PaO2 (mmHg) | Glu (mM) | |
|---|---|---|---|---|---|
| Pre-ischemia | |||||
| Control | 7.38 ± 0.03 | 83.6 ± 8.85 | 37.0 ± 3.92 | 105.2 ± 8.81 | 6.39 ± 1.23 |
| Vehicle | 7.39 ± 0.03 | 83.9 ± 8.53 | 36.7 ± 3.56 | 102.3 ± 8.35 | 6.34 ± 1.09 |
| Control-PEBP1 (10 mg/kg) | 7.43 ± 0.04 | 84.1 ± 9.10 | 37.2 ± 4.09 | 103.1 ± 9.02 | 6.42 ± 0.88 |
| PEP-1-PEBP1 (1 mg/kg) | 7.41 ± 0.04 | 83.4 ± 9.11 | 37.1 ± 4.55 | 104.2 ± 9.44 | 6.37 ± 1.03 |
| PEP-1-PEBP1 (3 mg/kg) | 7.43 ± 0.03 | 83.8 ± 8.95 | 36.8 ± 3.94 | 106.3 ± 9.21 | 6.48 ± 1.31 |
| PEP-1-PEBP1 (10 mg/kg) | 7.42 ± 0.04 | 83.5 ± 8.66 | 37.1 ± 4.45 | 106.9 ± 10.2 | 6.40 ± 1.07 |
| Reperfusion 10 min | |||||
| Control | 7.40 ± 0.04 | 84.8 ± 8.99 | 36.6 ± 3.96 | 107.4 ± 9.81 | 6.46 ± 0.98 |
| Vehicle | 7.37 ± 0.07 | 87.2 ± 10.2 | 39.4 ± 5.18 | 108.1 ± 9.60 | 6.93 ± 1.44 |
| Control-PEBP1 (10 mg/kg) | 7.35 ± 0.09 | 87.5 ± 11.3 | 38.8 ± 4.82 | 110.9 ± 10.8 | 7.05 ± 1.28 |
| PEP-1-PEBP1 (1 mg/kg) | 7.36 ± 0.06 | 88.2 ± 9.58 | 39.6 ± 6.02 | 106.5 ± 10.6 | 7.12 ± 1.36 |
| PEP-1-PEBP1 (3 mg/kg) | 7.37 ± 0.08 | 86.9 ± 9.83 | 38.4 ± 4.88 | 113.1 ± 12.2 | 7.07 ± 1.20 |
| PEP-1-PEBP1 (10 mg/kg) | 7.37 ± 0.10 | 85.2 ± 8.86 | 38.3 ± 5.03 | 109.8 ± 11.2 | 7.19 ± 1.24 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.; Cho, S.B.; Jung, H.Y.; Yoo, D.Y.; Oh, J.K.; Choi, G.-M.; Cho, T.-G.; Kim, D.W.; Hwang, I.K.; Choi, S.Y.; et al. Phosphatidylethanolamine-Binding Protein 1 Ameliorates Ischemia-Induced Inflammation and Neuronal Damage in the Rabbit Spinal Cord. Cells 2019, 8, 1370. https://doi.org/10.3390/cells8111370
Kim W, Cho SB, Jung HY, Yoo DY, Oh JK, Choi G-M, Cho T-G, Kim DW, Hwang IK, Choi SY, et al. Phosphatidylethanolamine-Binding Protein 1 Ameliorates Ischemia-Induced Inflammation and Neuronal Damage in the Rabbit Spinal Cord. Cells. 2019; 8(11):1370. https://doi.org/10.3390/cells8111370
Chicago/Turabian StyleKim, Woosuk, Su Bin Cho, Hyo Young Jung, Dae Young Yoo, Jae Keun Oh, Goang-Min Choi, Tack-Geun Cho, Dae Won Kim, In Koo Hwang, Soo Young Choi, and et al. 2019. "Phosphatidylethanolamine-Binding Protein 1 Ameliorates Ischemia-Induced Inflammation and Neuronal Damage in the Rabbit Spinal Cord" Cells 8, no. 11: 1370. https://doi.org/10.3390/cells8111370
APA StyleKim, W., Cho, S. B., Jung, H. Y., Yoo, D. Y., Oh, J. K., Choi, G. -M., Cho, T. -G., Kim, D. W., Hwang, I. K., Choi, S. Y., & Moon, S. M. (2019). Phosphatidylethanolamine-Binding Protein 1 Ameliorates Ischemia-Induced Inflammation and Neuronal Damage in the Rabbit Spinal Cord. Cells, 8(11), 1370. https://doi.org/10.3390/cells8111370