Isolation and Culture of Human Stem Cells from Apical Papilla under Low Oxygen Concentration Highlight Original Properties

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
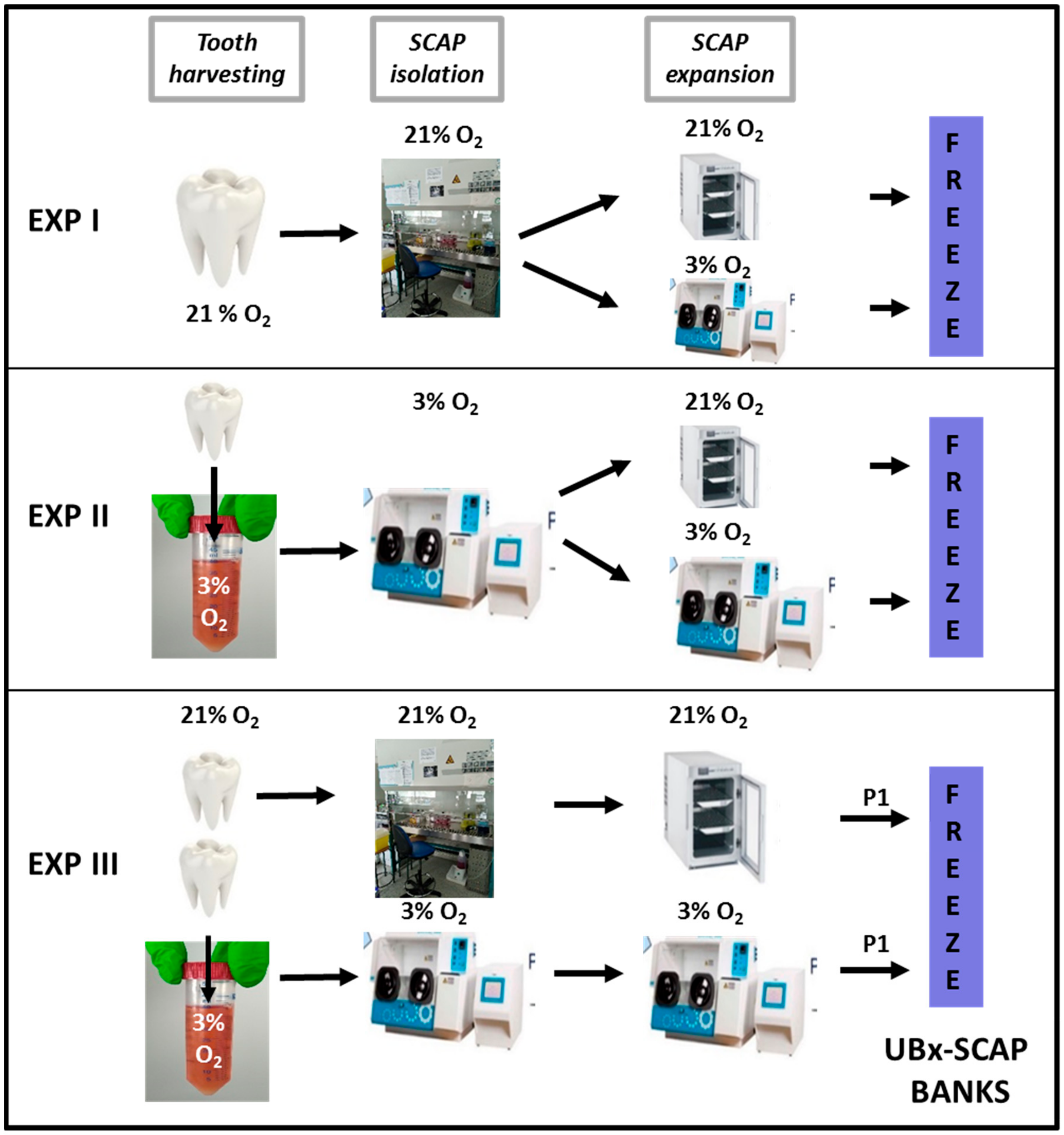
2.1. Tooth Recovery
2.2. Experimental Settings
2.3. Detailed UBx-SCAP Preparation
2.4. Cell Proliferation Assay
2.5. Colony Forming Unit (CFU) Assay
2.6. Cell Cycle Analysis
2.7. Flow Cytometry Analysis of Membrane Markers
2.8. Differentiation Procedures
2.9. Western Blotting and Immunolabelling
2.10. Statistical Analyses
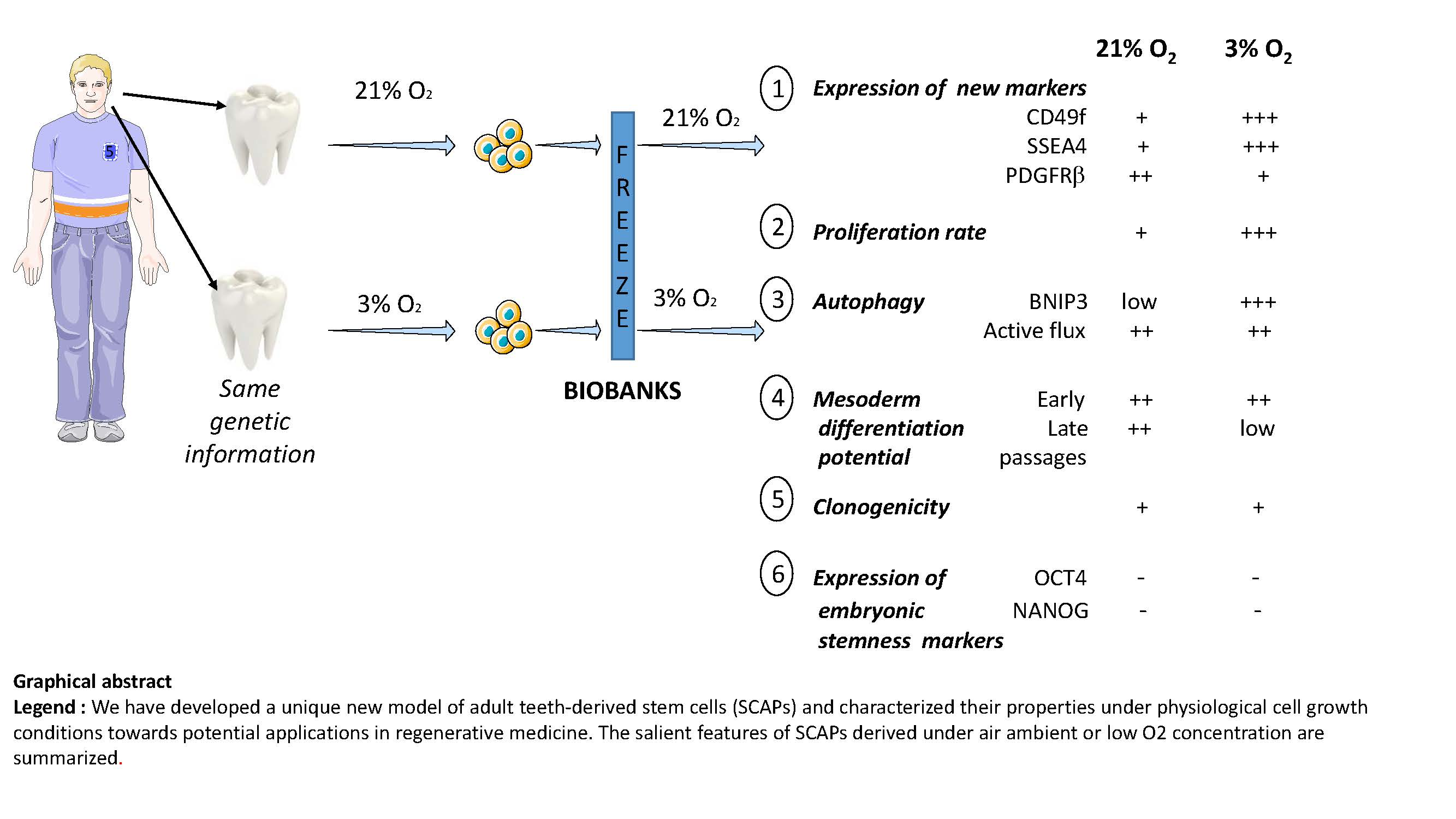
3. Results
3.1. SCAPs Display a Proliferative Advantage When Grown at 3% O2 Versus 21% O2
3.2. Clonogenicity of SCAPs In Vitro
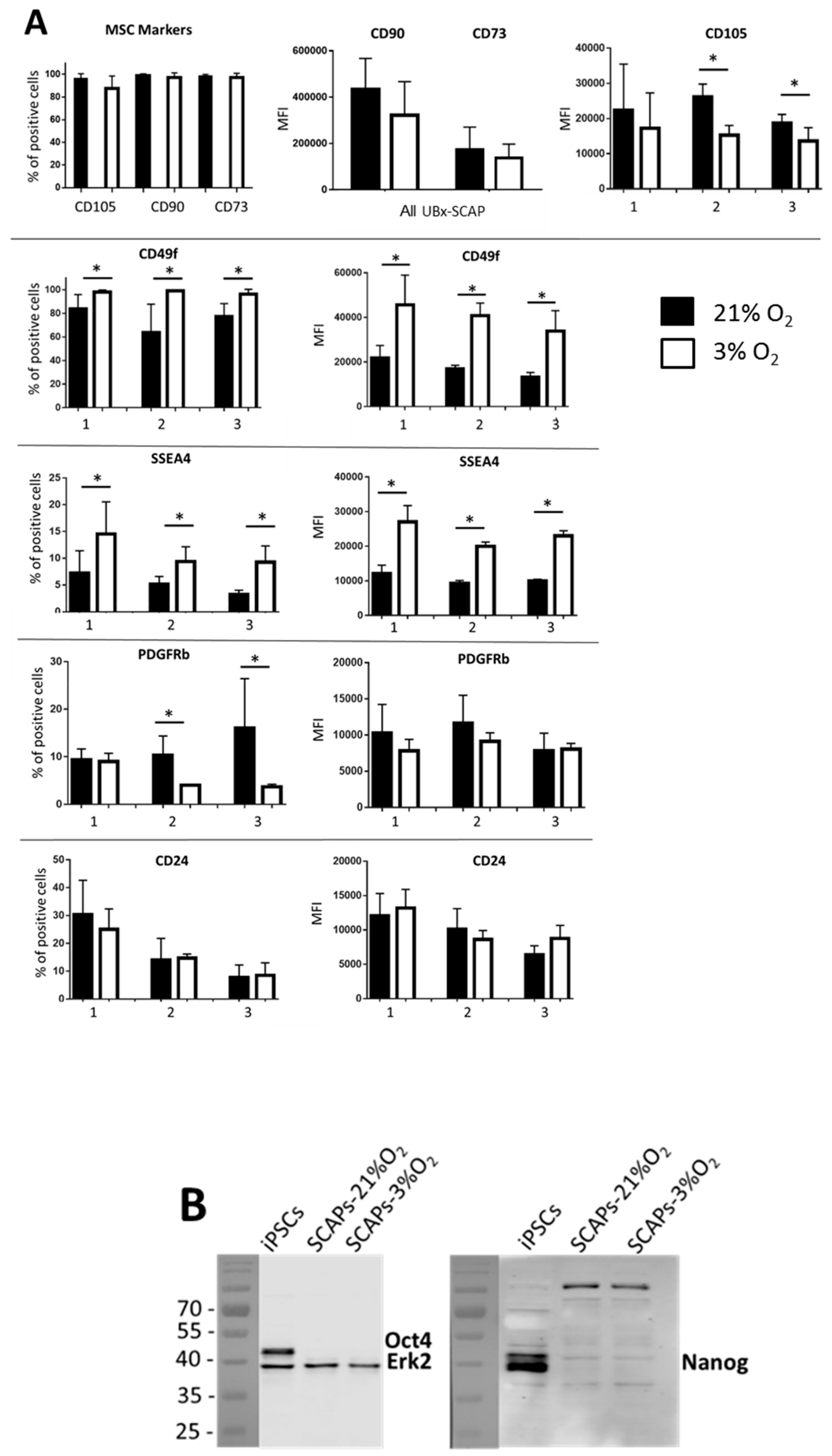
3.3. Differential Expression of Membrane Markers
3.4. SCAPs Differentiate into the Three Mesodermal Lineages but Lose Potential at Late Passages under Low [O2]
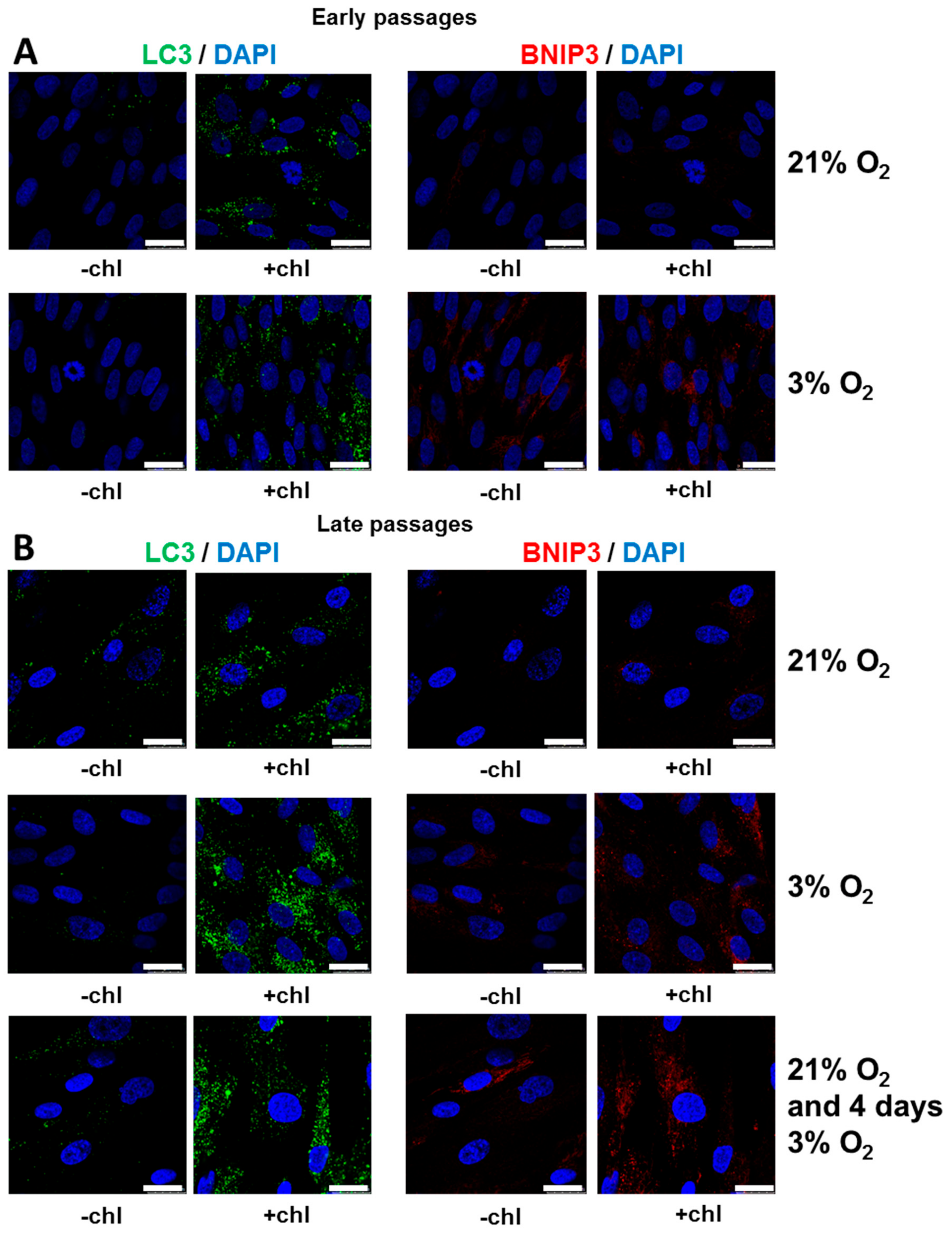
3.5. Autophagy is Increased under Low [O2] with Efficient Flux at Both 3% O2 and 21% O2
4. Discussion
4.1. High Proliferative Capacity but Not Better Clonogenicity under Low [O2]
4.2. Mesodermal Differentiation Potential of SCAPs is Maintained at Early but Not Late Passages under Low [O2]
4.3. Active Autophagy Remains for Long Periods in SCAPs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kobolak, J.; Dinnyes, A.; Memic, A.; Khademhosseini, A.; Mobasheri, A. Mesenchymal stem cells: Identification, phenotypic characterization, biological properties and potential for regenerative medicine through biomaterial micro-engineering of their niche. Methods San Diego Calif. 2016, 99, 62–68. [Google Scholar] [CrossRef]
- Liu, J.; Yu, F.; Sun, Y.; Jiang, B.; Zhang, W.; Yang, J.; Xu, G.-T.; Liang, A.; Liu, S. Concise reviews: Characteristics and potential applications of human dental tissue-derived mesenchymal stem cells. Stem Cells Dayt. Ohio 2015, 33, 627–638. [Google Scholar] [CrossRef]
- Rastegar, F.; Shenaq, D.; Huang, J.; Zhang, W.; Zhang, B.-Q.; He, B.-C.; Chen, L.; Zuo, G.-W.; Luo, Q.; Shi, Q.; et al. Mesenchymal stem cells: Molecular characteristics and clinical applications. World J. Stem Cells 2010, 2, 67–80. [Google Scholar]
- Hoogduijn, M.J.; Lombardo, E. Concise Review: Mesenchymal Stromal Cells Anno 2019: Dawn of the Therapeutic Era? Stem Cells Transl. Med. 2019, 8, 1126–1134. [Google Scholar] [CrossRef]
- Huang, G.T.-J.; Sonoyama, W.; Liu, Y.; Liu, H.; Wang, S.; Shi, S. The hidden treasure in apical papilla: The potential role in pulp/dentin regeneration and bioroot engineering. J. Endod. 2008, 34, 645–651. [Google Scholar] [CrossRef]
- Sonoyama, W.; Liu, Y.; Yamaza, T.; Tuan, R.S.; Wang, S.; Shi, S.; Huang, G.T.-J. Characterization of the apical papilla and its residing stem cells from human immature permanent teeth: A pilot study. J. Endod. 2008, 34, 166–171. [Google Scholar] [CrossRef]
- Bakopoulou, A.; About, I. Stem Cells of Dental Origin: Current Research Trends and Key Milestones towards Clinical Application. Stem Cells Int. 2016, 2016, 4209891. [Google Scholar] [CrossRef]
- Kang, J.; Fan, W.; Deng, Q.; He, H.; Huang, F. Stem Cells from the Apical Papilla: A Promising Source for Stem Cell-Based Therapy. BioMed. Res. Int. 2019, 2019, 6104738. [Google Scholar] [CrossRef]
- Nada, O.A.; El Backly, R.M. Stem Cells from the Apical Papilla (SCAP) as a Tool for Endogenous Tissue Regeneration. Front. Bioeng. Biotechnol. 2018, 6, 103. [Google Scholar] [CrossRef]
- Leyendecker Junior, A.; Gomes Pinheiro, C.C.; Lazzaretti Fernandes, T.; Franco Bueno, D. The use of human dental pulp stem cells for in vivo bone tissue engineering: A systematic review. J. Tissue Eng. 2018, 9, 2041731417752766. [Google Scholar] [CrossRef]
- Grayson, W.L.; Zhao, F.; Bunnell, B.; Ma, T. Hypoxia enhances proliferation and tissue formation of human mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2007, 358, 948–953. [Google Scholar] [CrossRef]
- Ivanovic, Z. Hypoxia or in situ normoxia: The stem cell paradigm. J. Cell. Physiol. 2009, 219, 271–275. [Google Scholar] [CrossRef]
- Ivanovic, Z.; Vlaski-Lafarge, M. Harnessing anaerobic nature of stem cells for use in regenerative medicine. In Anaerobiosis and Stemness: An Evolutionary Paradigm; Academic Press: Cambridge, MA, USA, 2016; pp. 258–286. [Google Scholar]
- Gorin, C.; Rochefort, G.Y.; Bascetin, R.; Ying, H.; Lesieur, J.; Sadoine, J.; Beckouche, N.; Berndt, S.; Novais, A.; Lesage, M.; et al. Priming Dental Pulp Stem Cells with Fibroblast Growth Factor-2 Increases Angiogenesis of Implanted Tissue-Engineered Constructs Through Hepatocyte Growth Factor and Vascular Endothelial Growth Factor Secretion. Stem Cells Transl. Med. 2016, 5, 392–404. [Google Scholar] [CrossRef]
- Shi, R.; Yang, H.; Lin, X.; Cao, Y.; Zhang, C.; Fan, Z.; Hou, B. Analysis of the characteristics and expression profiles of coding and noncoding RNAs of human dental pulp stem cells in hypoxic conditions. Stem Cell Res. Ther. 2019, 10, 89. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Diabira, S.; Howard, G.A.; Menei, P.; Roos, B.A.; Schiller, P.C. Marrow-isolated adult multilineage inducible (MIAMI) cells, a unique population of postnatal young and old human cells with extensive expansion and differentiation potential. J. Cell Sci. 2004, 117, 2971–2981. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Diabira, S.; Howard, G.A.; Roos, B.A.; Schiller, P.C. Low oxygen tension inhibits osteogenic differentiation and enhances stemness of human MIAMI cells. Bone 2006, 39, 513–522. [Google Scholar] [CrossRef]
- Rios, C.; D’Ippolito, G.; Curtis, K.M.; Delcroix, G.J.-R.; Gomez, L.A.; El Hokayem, J.; Rieger, M.; Parrondo, R.; de Las Pozas, A.; Perez-Stable, C.; et al. Low Oxygen Modulates Multiple Signaling Pathways, Increasing Self-Renewal, While Decreasing Differentiation, Senescence, and Apoptosis in Stromal MIAMI Cells. Stem Cells Dev. 2016, 25, 848–860. [Google Scholar] [CrossRef]
- Lennon, D.P.; Edmison, J.M.; Caplan, A.I. Cultivation of rat marrow-derived mesenchymal stem cells in reduced oxygen tension: Effects on in vitro and in vivo osteochondrogenesis. J. Cell. Physiol. 2001, 187, 345–355. [Google Scholar] [CrossRef]
- Ren, H.; Cao, Y.; Zhao, Q.; Li, J.; Zhou, C.; Liao, L.; Jia, M.; Zhao, Q.; Cai, H.; Han, Z.C.; et al. Proliferation and differentiation of bone marrow stromal cells under hypoxic conditions. Biochem. Biophys. Res. Commun. 2006, 347, 12–21. [Google Scholar] [CrossRef]
- Ivanovic, Z.; Vlaski-Lafarge, M. In situ normoxia versus “Hypoxia”. In Anaerobiosis and Stemness: An Evolutionary Paradigm; Academic Press: Cambridge, MA, USA, 2016; pp. 17–21. [Google Scholar]
- Schäfer, R.; Spohn, G.; Baer, P.C. Mesenchymal Stem/Stromal Cells in Regenerative Medicine: Can Preconditioning Strategies Improve Therapeutic Efficacy? Transfus. Med. Hemother. 2016, 43, 256–267. [Google Scholar] [CrossRef]
- Choi, J.R.; Pingguan-Murphy, B.; Wan Abas, W.A.B.; Noor Azmi, M.A.; Omar, S.Z.; Chua, K.H.; Wan Safwani, W.K.Z. Impact of low oxygen tension on stemness, proliferation and differentiation potential of human adipose-derived stem cells. Biochem. Biophys. Res. Commun. 2014, 448, 218–224. [Google Scholar] [CrossRef]
- Fotia, C.; Massa, A.; Boriani, F.; Baldini, N.; Granchi, D. Prolonged exposure to hypoxic milieu improves the osteogenic potential of adipose derived stem cells. J. Cell. Biochem. 2015, 116, 1442–1453. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Sanz-Ros, J.; Román-Domínguez, A.; Inglés, M.; Gimeno-Mallench, L.; El Alami, M.; Viña-Almunia, J.; Gambini, J.; Viña, J.; Borrás, C. Relevance of Oxygen Concentration in Stem Cell Culture for Regenerative Medicine. Int. J. Mol. Sci. 2019, 20, 1195. [Google Scholar] [CrossRef]
- Boya, P.; Codogno, P.; Rodriguez-Muela, N. Autophagy in stem cells: Repair, remodelling and metabolic reprogramming. Dev. Camb. Engl. 2018, 145, dev146506. [Google Scholar] [CrossRef]
- Meijer, A.J.; Codogno, P. Regulation and role of autophagy in mammalian cells. Int. J. Biochem. Cell Biol. 2004, 36, 2445–2462. [Google Scholar] [CrossRef]
- Sotthibundhu, A.; Promjuntuek, W.; Liu, M.; Shen, S.; Noisa, P. Roles of autophagy in controlling stem cell identity: A perspective of self-renewal and differentiation. Cell Tissue Res. 2018, 374, 205–216. [Google Scholar] [CrossRef]
- Guan, J.-L.; Simon, A.K.; Prescott, M.; Menendez, J.A.; Liu, F.; Wang, F.; Wang, C.; Wolvetang, E.; Vazquez-Martin, A.; Zhang, J. Autophagy in stem cells. Autophagy 2013, 9, 830–849. [Google Scholar] [CrossRef]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 conjugation system in mammalian autophagy. Int. J. Biochem. Cell Biol. 2004, 36, 2503–2518. [Google Scholar] [CrossRef]
- Semenza, G.L. Mitochondrial autophagy: Life and breath of the cell. Autophagy 2008, 4, 534–536. [Google Scholar] [CrossRef]
- Ney, P.A. Mitochondrial autophagy: Origins, significance, and role of BNIP3 and NIX. Biochim. Biophys. Acta 2015, 1853, 2775–2783. [Google Scholar] [CrossRef]
- Lee, H.J.; Jung, Y.H.; Choi, G.E.; Ko, S.H.; Lee, S.-J.; Lee, S.H.; Han, H.J. BNIP3 induction by hypoxia stimulates FASN-dependent free fatty acid production enhancing therapeutic potential of umbilical cord blood-derived human mesenchymal stem cells. Redox Biol. 2017, 13, 426–443. [Google Scholar] [CrossRef]
- Martin, D.E.; De Almeida, J.F.A.; Henry, M.A.; Khaing, Z.Z.; Schmidt, C.E.; Teixeira, F.B.; Diogenes, A. Concentration-dependent effect of sodium hypochlorite on stem cells of apical papilla survival and differentiation. J. Endod. 2014, 40, 51–55. [Google Scholar] [CrossRef]
- Devillard, R.; Rémy, M.; Kalisky, J.; Bourget, J.-M.; Kérourédan, O.; Siadous, R.; Bareille, R.; Amédée-Vilamitjana, J.; Chassande, O.; Fricain, J.-C. In vitro assessment of a collagen/alginate composite scaffold for regenerative endodontics. Int. Endod. J. 2017, 50, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.H.; Campbell, G.R. Culture techniques and their applications to studies of vascular smooth muscle. Clin. Sci. 1993, 85, 501–513. [Google Scholar] [CrossRef]
- Greenwood, S.K.; Hill, R.B.; Sun, J.T.; Armstrong, M.J.; Johnson, T.E.; Gara, J.P.; Galloway, S.M. Population doubling: A simple and more accurate estimation of cell growth suppression in the in vitro assay for chromosomal aberrations that reduces irrelevant positive results. Environ. Mol. Mutagen. 2004, 43, 36–44. [Google Scholar] [CrossRef]
- Duval, D.; Trouillas, M.; Thibault, C.; Dembele, D.; Diemunsch, F.; Reinhardt, B.; Mertz, A.L.; Dierich, A.; Boeuf, H. Apoptosis and differentiation commitment: Novel insights revealed by gene profiling studies in mouse embryonic stem cells. Cell Death Differ. 2006, 13, 564–575. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, A.A.; Kirstein, N.; Mournetas, V.; Darracq, A.; Broc, S.; Blanchard, C.; Zeineddine, D.; Mortada, M.; Boeuf, H. Murine Embryonic Stem Cell Plasticity Is Regulated through Klf5 and Maintained by Metalloproteinase MMP1 and Hypoxia. PLoS ONE 2016, 11, e0146281. [Google Scholar] [CrossRef] [Green Version]
- Smyrek, I.; Stelzer, E.H.K. Quantitative three-dimensional evaluation of immunofluorescence staining for large whole mount spheroids with light sheet microscopy. Biomed. Opt. Express 2017, 8, 484–499. [Google Scholar] [CrossRef] [Green Version]
- Coronado, D.; Godet, M.; Bourillot, P.-Y.; Tapponnier, Y.; Bernat, A.; Petit, M.; Afanassieff, M.; Markossian, S.; Malashicheva, A.; Iacone, R.; et al. A short G1 phase is an intrinsic determinant of naïve embryonic stem cell pluripotency. Stem Cell Res. 2013, 10, 118–131. [Google Scholar] [CrossRef]
- Elabd, C.; Ichim, T.E.; Miller, K.; Anneling, A.; Grinstein, V.; Vargas, V.; Silva, F.J. Comparing atmospheric and hypoxic cultured mesenchymal stem cell transcriptome: Implication for stem cell therapies targeting intervertebral discs. J. Transl. Med. 2018, 16, 222. [Google Scholar] [CrossRef]
- Antebi, B.; Rodriguez, L.A.; Walker, K.P.; Asher, A.M.; Kamucheka, R.M.; Alvarado, L.; Mohammadipoor, A.; Cancio, L.C. Short-term physiological hypoxia potentiates the therapeutic function of mesenchymal stem cells. Stem Cell Res. Ther. 2018, 9, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, P.; Lertchirakarn, V. Comparison of stem cell behaviors between indigenous high and low-CD24 percentage expressing cells of stem cells from apical papilla (SCAPs). Tissue Cell 2016, 48, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Mo, M.; Wang, J.; Sadia, S.; Shi, B.; Fu, X.; Yu, L.; Tredget, E.E.; Wu, Y. Platelet-derived growth factor receptor beta identifies mesenchymal stem cells with enhanced engraftment to tissue injury and pro-angiogenic property. Cell. Mol. Life Sci. 2018, 75, 547–561. [Google Scholar] [CrossRef]
- Henderson, J.K.; Draper, J.S.; Baillie, H.S.; Fishel, S.; Thomson, J.A.; Moore, H.; Andrews, P.W. Preimplantation human embryos and embryonic stem cells show comparable expression of stage-specific embryonic antigens. Stem Cells 2002, 20, 329–337. [Google Scholar] [CrossRef]
- Yu, K.-R.; Yang, S.-R.; Jung, J.-W.; Kim, H.; Ko, K.; Han, D.W.; Park, S.-B.; Choi, S.W.; Kang, S.-K.; Schöler, H.; et al. CD49f enhances multipotency and maintains stemness through the direct regulation of OCT4 and SOX2. Stem Cells Dayt. Ohio 2012, 30, 876–887. [Google Scholar] [CrossRef]
- Alvarez, R.; Lee, H.-L.; Hong, C.; Wang, C.-Y. Single CD271 marker isolates mesenchymal stem cells from human dental pulp. Int. J. Oral Sci. 2015, 7, 205–212. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Sbrana, F.V.; Cortini, M.; Avnet, S.; Perut, F.; Columbaro, M.; De Milito, A.; Baldini, N. The Role of Autophagy in the Maintenance of Stemness and Differentiation of Mesenchymal Stem Cells. Stem Cell Rev. 2016, 12, 621–633. [Google Scholar] [CrossRef]
- Cossu, G.; Birchall, M.; Brown, T.; De Coppi, P.; Culme-Seymour, E.; Gibbon, S.; Hitchcock, J.; Mason, C.; Montgomery, J.; Morris, S.; et al. Lancet Commission: Stem cells and regenerative medicine. Lancet 2018, 391, 883–910. [Google Scholar] [CrossRef] [Green Version]
- Cipolleschi, M.G.; Rovida, E.; Ivanovic, Z.; Praloran, V.; Olivotto, M.; Sbarba, P.D. The expansion of murine bone marrow cells preincubated in hypoxia as an in vitro indicator of their marrow-repopulating ability. Leukemia 2000, 14, 735–739. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, M.; Vlaski, M.; Duchez, P.; Chevaleyre, J.; Lafarge, X.; Boiron, J.-M.; Praloran, V.; Brunet De La Grange, P.; Ivanovic, Z. Combination of low O2 concentration and mesenchymal stromal cells during culture of cord blood CD34(+) cells improves the maintenance and proliferative capacity of hematopoietic stem cells. J. Cell. Physiol. 2012, 227, 2750–2758. [Google Scholar] [CrossRef] [PubMed]
- Leroux, L.; Descamps, B.; Tojais, N.F.; Séguy, B.; Oses, P.; Moreau, C.; Daret, D.; Ivanovic, Z.; Boiron, J.-M.; Lamazière, J.-M.D.; et al. Hypoxia preconditioned mesenchymal stem cells improve vascular and skeletal muscle fiber regeneration after ischemia through a Wnt4-dependent pathway. Mol. Ther. J. Am. Soc. Gene Ther. 2010, 18, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Grau-Monge, C.; Delcroix, G.J.-R.; Bonnin-Marquez, A.; Valdes, M.; Awadallah, E.L.M.; Quevedo, D.F.; Armour, M.R.; Montero, R.B.; Schiller, P.C.; Andreopoulos, F.M.; et al. Marrow-isolated adult multilineage inducible cells embedded within a biologically-inspired construct promote recovery in a mouse model of peripheral vascular disease. Biomed. Mater. 2017, 12, 015024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niculescu, C.; Ganguli-Indra, G.; Pfister, V.; Dupé, V.; Messaddeq, N.; De Arcangelis, A.; Georges-Labouesse, E. Conditional ablation of integrin alpha-6 in mouse epidermis leads to skin fragility and inflammation. Eur. J. Cell Biol. 2011, 90, 270–277. [Google Scholar] [CrossRef]
- De Arcangelis, A.; Hamade, H.; Alpy, F.; Normand, S.; Bruyère, E.; Lefebvre, O.; Méchine-Neuville, A.; Siebert, S.; Pfister, V.; Lepage, P.; et al. Hemidesmosome integrity protects the colon against colitis and colorectal cancer. Gut 2017, 66, 1748–1760. [Google Scholar] [CrossRef]
- Qiryaqoz, Z.; Timilsina, S.; Czarnowski, D.; Krebsbach, P.H.; Villa-Diaz, L.G. Identification of biomarkers indicative of functional skeletal stem cells. Orthod. Craniofac. Res. 2019, 22, 192–198. [Google Scholar] [CrossRef]
- Yang, Z.; Dong, P.; Fu, X.; Li, Q.; Ma, S.; Wu, D.; Kang, N.; Liu, X.; Yan, L.; Xiao, R. CD49f Acts as an Inflammation Sensor to Regulate Differentiation, Adhesion, and Migration of Human Mesenchymal Stem Cells. Stem Cells Dayt. Ohio 2015, 33, 2798–2810. [Google Scholar] [CrossRef]
- Inoue, H.; Nagata, N.; Kurokawa, H.; Yamanaka, S. iPS cells: A game changer for future medicine. EMBO J. 2014, 33, 409–417. [Google Scholar] [CrossRef]
- Takahashi, K.; Okita, K.; Nakagawa, M.; Yamanaka, S. Induction of pluripotent stem cells from fibroblast cultures. Nat. Protoc. 2007, 2, 3081–3089. [Google Scholar] [CrossRef]
- Pattappa, G.; Thorpe, S.D.; Jegard, N.C.; Heywood, H.K.; de Bruijn, J.D.; Lee, D.A. Continuous and uninterrupted oxygen tension influences the colony formation and oxidative metabolism of human mesenchymal stem cells. Tissue Eng. Part. C Methods 2013, 19, 68–79. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.; Fatima, F.; Vallabhaneni, K.C.; Penfornis, P.; Valadi, H.; Ekström, K.; Kholia, S.; Whitt, J.D.; Fernandes, J.D.; Pochampally, R.; et al. Extracellular Vesicles: Evolving Factors in Stem Cell Biology. Stem Cells Int. 2016, 2016, 1073140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. Regulation of mammalian O2 homeostasis by hypoxia-inducible factor 1. Annu. Rev. Cell Dev. Biol. 1999, 15, 551–578. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Oxygen sensing, hypoxia-inducible factors, and disease pathophysiology. Annu. Rev. Pathol. 2014, 9, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; He, Y.; Lu, F. Autophagy in Stem Cell Biology: A Perspective on Stem Cell Self-Renewal and Differentiation. Stem Cells Int. 2018, 2018, 9131397. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Li, R.; Tang, H.; Zhu, T.; Xu, F.; Zhu, J. Regulation of autophagy in mesenchymal stem cells modulates therapeutic effects on spinal cord injury. Brain Res. 2019, 146321. [Google Scholar] [CrossRef]
- Esteban-Martínez, L.; Boya, P. BNIP3L/NIX-dependent mitophagy regulates cell differentiation via metabolic reprogramming. Autophagy 2017, 14, 915–917. [Google Scholar] [CrossRef]
- Singh, A.; Azad, M.; Shymko, M.D.; Henson, E.S.; Katyal, S.; Eisenstat, D.D.; Gibson, S.B. The BH3 only Bcl-2 family member BNIP3 regulates cellular proliferation. PLoS ONE 2018, 13, e0204792. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Antigen Targeted | Supplier | Host Isotype | Clone Reference | Concentration (Stock Sol.) |
|---|---|---|---|---|---|
| Antibodies for Flow Cytometry Analyses | |||||
| Isotype PE | None | eBioscience | Mouse IgG1, κ | 12-4714-42 | 0.1 µg/µL |
| Isotype APC | None | eBioscience | Mouse IgG1, κ | 17-4714-42 | 0.1 µg/µL |
| CD24 APC | CD24 GPI-anchored glycoprotein | eBioscience | Mouse IgG1, κ | eBioSN3 17-0247-42 | 0.05 µg/µL |
| CD49f PE | CD49f Integrin α6 | eBioscience | Rat IgG2a, κ | eBioGoH3 12-0495-81 | 0.2 µg/µL |
| CD73 PE | CD73 ecto-5′-nucléotidase | eBioscience | Mouse IgG1, κ | AD2 12-0739-42 | 0.025 µg/µL |
| CD90 PE | CD90 Thy1 | eBioscience | Mouse IgG1, κ | eBio5E10 12-0909-42 | 0.05 µg/µL |
| CD105 APC | CD105 endoglin | eBioscience | Mouse IgG1, κ | SN6 17-1057-42 | 0.05 µg/µL |
| PDGFβ PE | PDGFRβ Platelet Derived Growth Factorβ | Biolegend | Mouse IgG1, κ | 18A2 323605 | 400 µg/mL |
| CD271 PE | CD271 nerve growth factor receptor | eBioscience | Mouse IgG1, κ | ME20.4 12-9400-42 | 0.05 µg/µL |
| SSEA4 APC | SSEA-4, stage specific embryonic antigen 4 | BioLegend | Mouse IgG3, κ | MC-813-70 330417 | 50 µg/mL |
| CD45 APC | LCA, leucocyte common antigen | eBioscience | Mouse IgG1, κ | HI30-17-0459-42 | 0.012 µg/µL |
| CD34 APC | Mucosialin, CD34 antigen | eBioscience | Mouse IgG1, κ | 4H11-17-0349-42 | 0.05 µg/µL |
| Antibodies for Immunolabelling Studies | |||||
| LC3 | Microtubule-associated protein light chain 3 | MBL | Mouse hybridoma IgG1, κ | M152-3 | 2 µg/µL |
| BNIP3 | Bcl2/adenovirus 19-kDa-interacting protein 3 | Abcam | Rabbit monoclonal | Ab109362 | 1.2 µg/µL |
| Alexa 488 Goat | Mouse IgG (H + L) | Invitrogen | Goat Polyclonal | A11001 | 2 µg/µL |
| Alexa 568 Goat | Rabbit IgG (H + L) | Invitrogen | Goat Polyclonal | A11036 | 2 µg/µL |
| Antibodies for Western Blot | |||||
| Nanog | Nanog | Abcam | Rabbit Monoclonal | EPR2027 | 0.3 mg/mL |
| Oct4 | Oct4 Transcription factor that binds to an octamer motif | Abcam | IgG Rabbit Polyclonal | Ab19857 | 1 mg/mL |
| ERK2 | Extracellular Regulated Kinase 2 | Santa Cruz Biotechnol. | IgG Rabbit Polyclonal | Sc154 | 0.2 mg/mL |
| Ac II-HRP | Rabbit IgG (H+L) | Jackson Immuno Research | Goat Peroxidase Conjugated | 111-035-144 | 0.8 mg/mL |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rémy, M.; Ferraro, F.; Le Salver, P.; Rey, S.; Genot, E.; Djavaheri-Mergny, M.; Thébaud, N.; Boiziau, C.; Boeuf, H. Isolation and Culture of Human Stem Cells from Apical Papilla under Low Oxygen Concentration Highlight Original Properties. Cells 2019, 8, 1485. https://doi.org/10.3390/cells8121485
Rémy M, Ferraro F, Le Salver P, Rey S, Genot E, Djavaheri-Mergny M, Thébaud N, Boiziau C, Boeuf H. Isolation and Culture of Human Stem Cells from Apical Papilla under Low Oxygen Concentration Highlight Original Properties. Cells. 2019; 8(12):1485. https://doi.org/10.3390/cells8121485
Chicago/Turabian StyleRémy, Murielle, Francesca Ferraro, Pierre Le Salver, Sylvie Rey, Elisabeth Genot, Mojgan Djavaheri-Mergny, Noélie Thébaud, Claudine Boiziau, and Hélène Boeuf. 2019. "Isolation and Culture of Human Stem Cells from Apical Papilla under Low Oxygen Concentration Highlight Original Properties" Cells 8, no. 12: 1485. https://doi.org/10.3390/cells8121485
APA StyleRémy, M., Ferraro, F., Le Salver, P., Rey, S., Genot, E., Djavaheri-Mergny, M., Thébaud, N., Boiziau, C., & Boeuf, H. (2019). Isolation and Culture of Human Stem Cells from Apical Papilla under Low Oxygen Concentration Highlight Original Properties. Cells, 8(12), 1485. https://doi.org/10.3390/cells8121485