Dietary Immunostimulant CpG Modulates MicroRNA Biomarkers Associated with Immune Responses in Atlantic Salmon (Salmo salar)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Feed Production
2.2. Feeding Trial, Immune Challenge, and Fish Sampling
2.3. RNA Isolation
2.4. Library Preparation and Deep Sequencing
2.5. Analysis of Deep Sequencing Data
2.6. Prediction of Target Genes and Their Functional Annotations
2.7. qPCR Analysis of miRNA Expression
2.8. Statistical Analyses
3. Results
3.1. Deep Sequencing and Identification of Differentially Expressed miRNAs
3.2. qPCR Validation of DESeq2-Identified pIC- and/or ASAL-Responsive miRNAs
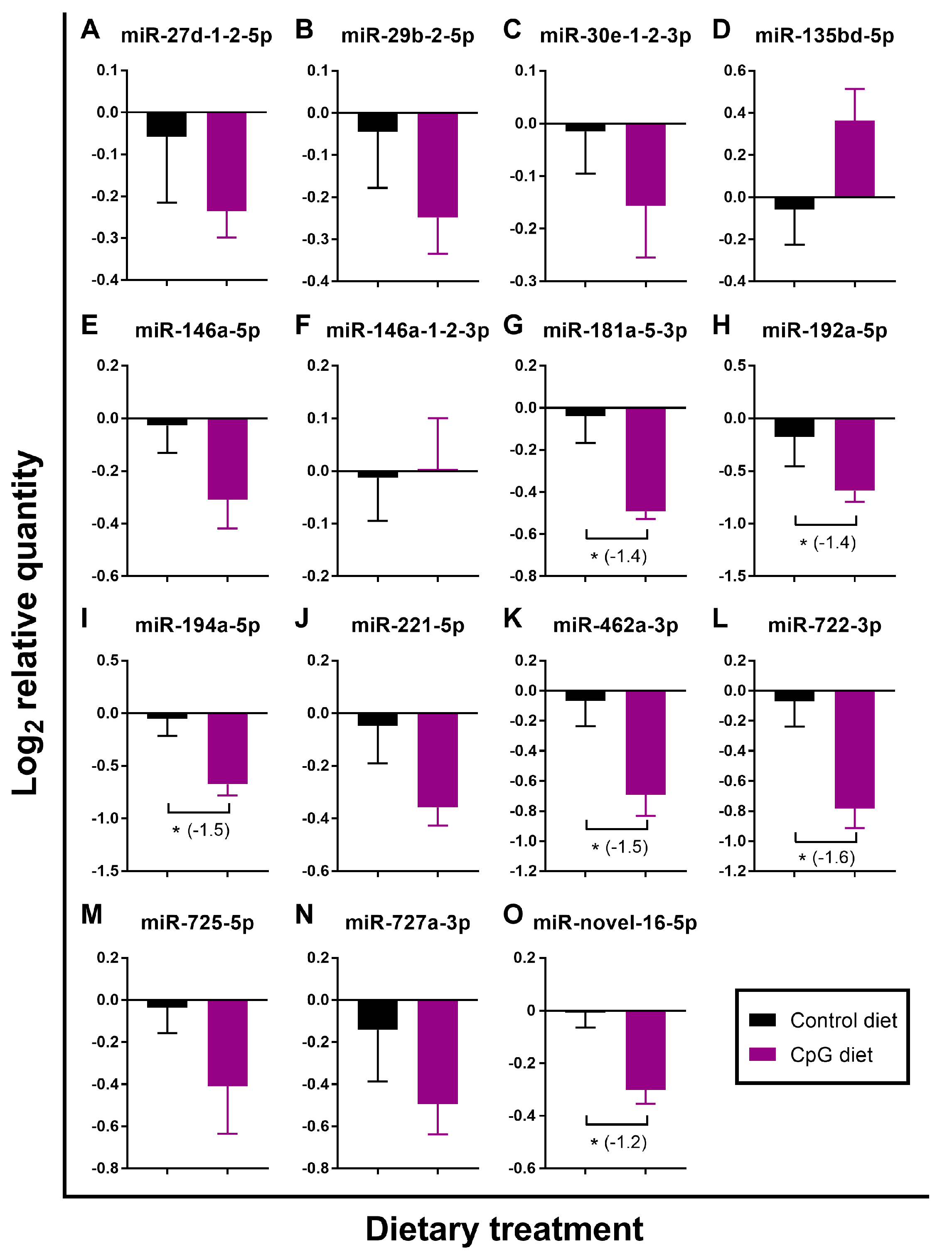
3.3. Impact of Diets on the Expression of pIC- and/or ASAL-Responsive miRNAs
3.4. Multivariate Statistical Analyses
4. Discussion
4.1. Deep Sequencing and Identification of Differentially Expressed miRNAs
4.2. miRNAs Associated with Both Antiviral and Antibacterial Immune Responses in Atlantic Salmon Head Kidney
4.3. miRNAs Only Associated with Antiviral Immune Response in Atlantic Salmon Head Kidney
4.4. miRNAs Only Associated with Antibacterial Immune Response in Atlantic Salmon Head Kidney
4.5. CpG Supplement Modulated the Expression of Immune-Relevant miRNAs in ASAL-Treated Atlantic Salmon
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agaba, M.K.; Tocher, D.R.; Zheng, X.; Dickson, C.A.; Dick, J.R.; Teale, A.J. Cloning and functional characterisation of polyunsaturated fatty acid elongases of marine and freshwater teleost fish. Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2005, 142, 342–352. [Google Scholar] [CrossRef]
- Tocher, D.R.; Zheng, X.; Schlechtriem, C.; Hastings, N.; Dick, J.R.; Teale, A.J. Highly unsaturated fatty acid synthesis in marine fish: Cloning, functional characterization, and nutritional regulation of fatty acyl Δ6 desaturase of Atlantic cod (Gadus morhua L.). Lipids 2006, 41, 1003–1016. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goal; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018. [Google Scholar]
- Kiron, V. Fish immune system and its nutritional modulation for preventive health care. Anim. Feed Sci. Technol. 2012, 173, 111–133. [Google Scholar] [CrossRef]
- Caballero-Solares, A.; Hall, J.R.; Xue, X.; Eslamloo, K.; Taylor, R.G.; Parrish, C.C.; Rise, M.L. The dietary replacement of marine ingredients by terrestrial animal and plant alternatives modulates the antiviral immune response of Atlantic salmon (Salmo salar). Fish Shellfish Immunol. 2017, 64, 24–38. [Google Scholar] [CrossRef]
- Lang, A.S.; Rise, M.L.; Culley, A.I.; Steward, G.F. RNA viruses in the sea. FEMS Microbiol. Rev. 2009, 33, 295–323. [Google Scholar] [CrossRef] [Green Version]
- Rise, M.L.; Jones, S.R.M.; Brown, G.D.; von Schalburg, K.R.; Davidson, W.S.; Koop, B.F. Microarray analyses identify molecular biomarkers of Atlantic salmon macrophage and hematopoietic kidney responses to Piscirickettsia salmonis infection. Physiol. Genom. 2004, 20, 21–35. [Google Scholar] [CrossRef]
- Ewart, K.V.; Belanger, J.C.; Williams, J.; Karakach, T.; Penny, S.; Tsoi, S.C.; Richards, R.C.; Douglas, S.E. Identification of genes differentially expressed in Atlantic salmon (Salmo salar) in response to infection by Aeromonas salmonicida using cDNA microarray technology. Dev. Comp. Immunol. 2005, 29, 333–347. [Google Scholar] [CrossRef]
- Murray, A.G.; Munro, L.A.; Wallace, I.S.; Allan, C.E.T.; Peeler, E.J.; Thrush, M.A. Epidemiology of Renibacterium salmoninarum in Scotland and the potential for compartmentalised management of salmon and trout farming areas. Aquaculture 2012, 324, 1–13. [Google Scholar] [CrossRef]
- Løvoll, M.; Wiik-Nielsen, C.R.; Tunsjø, H.S.; Colquhoun, D.; Lunder, T.; Sørum, H.; Grove, S. Atlantic salmon bath challenged with Moritella viscosa–Pathogen invasion and host response. Fish Shellfish Immunol. 2009, 26, 877–884. [Google Scholar] [CrossRef]
- Vallejos-Vidal, E.; Reyes-López, F.; Teles, M.; MacKenzie, S. The response of fish to immunostimulant diets. Fish Shellfish Immunol. 2016, 56, 34–69. [Google Scholar] [CrossRef]
- Secombes, C.J.; Wang, T. The innate and adaptive immune system of fish. In Infectious Disease in Aquaculture; Austin, B., Ed.; Woodhead Publishing: Sawston, UK, 2012; pp. 3–68. [Google Scholar]
- Robertsen, B. The interferon system of teleost fish. Fish Shellfish Immunol. 2006, 20, 172–191. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Eslamloo, K.; Xue, X.; Booman, M.; Smith, N.C.; Rise, M.L. Transcriptome profiling of the antiviral immune response in Atlantic cod macrophages. Dev. Comp. Immunol. 2016, 63, 187–205. [Google Scholar] [CrossRef]
- Feng, C.Y.; Johnson, S.C.; Hori, T.S.; Rise, M.; Hall, J.R.; Gamperl, A.K.; Hubert, S.; Kimball, J.; Bowman, S.; Rise, M.L. Identification and analysis of differentially expressed genes in immune tissues of Atlantic cod stimulated with formalin-killed, atypical Aeromonas salmonicida. Physiol. Genom. 2009, 37, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.S.; Gamperl, A.K.; Nash, G.; Booman, M.; Barat, A.; Rise, M.L. The impact of a moderate chronic temperature increase on spleen immune-relevant gene transcription depends on whether Atlantic cod (Gadus morhua) are stimulated with bacterial versus viral antigens. Genome 2013, 56, 567–576. [Google Scholar] [CrossRef]
- Andreassen, R.; Høyheim, B. miRNAs associated with immune response in teleost fish. Dev. Comp. Immunol. 2017, 75, 77–85. [Google Scholar] [CrossRef]
- Andreassen, R.; Woldemariam, N.T.; Egeland, I.Ø.; Agafonov, O.; Sindre, H.; Høyheim, B. Identification of differentially expressed Atlantic salmon miRNAs responding to salmonid alphavirus (SAV) infection. BMC Genom. 2017, 18, 349. [Google Scholar] [CrossRef]
- Herkenhoff, M.E.; Oliveira, A.C.; Nachtigall, P.G.; Costa, J.M.; Campos, V.F.; Hilsdorf, A.W.; Pinhal, D. Fishing into the MicroRNA transcriptome. Front. Genet. 2018, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Eslamloo, K.; Inkpen, S.M.; Rise, M.L.; Andreassen, R. Discovery of microRNAs associated with the antiviral immune response of Atlantic cod macrophages. Mol. Immunol. 2018, 93, 152–161. [Google Scholar] [CrossRef] [Green Version]
- Woldemariam, N.T.; Agafonov, O.; Høyheim, B.; Houston, R.D.; Taggart, J.B.; Andreassen, R. Expanding the miRNA repertoire in Atlantic salmon; discovery of isomiRs and miRNAs highly expressed in different tissues and developmental stages. Cells 2019, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela-Miranda, D.; Valenzuela-Muñoz, V.; Farlora, R.; Gallardo-Escárate, C. MicroRNA-based transcriptomic responses of Atlantic salmon during infection by the intracellular bacterium Piscirickettsia salmonis. Dev. Comp. Immunol. 2017, 77, 287–296. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, D.; Li, S.; Zhao, J.; Xu, L.; Liu, H.; Lu, T.; Mou, Z. A transcriptome analysis focusing on splenic immune-related mciroRNAs of rainbow trout upon Aeromonas salmonicida subsp. salmonicida infection. Fish Shellfish Immunol. 2019, 91, 350–357. [Google Scholar] [CrossRef]
- Tacchi, L.; Bickerdike, R.; Douglas, A.; Secombes, C.J.; Martin, S.A.M. Transcriptomic responses to functional feeds in Atlantic salmon (Salmo salar). Fish Shellfish Immunol. 2011, 31, 704–715. [Google Scholar] [CrossRef]
- Martin, S.A.M.; Król, E. Nutrigenomics and immune function in fish: New insights from omics technologies. Dev. Comp. Immunol. 2017, 75, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Covello, J.M.; Friend, S.E.; Purcell, S.L.; Burka, J.F.; Markham, R.J.F.; Donkin, A.W.; Groman, D.B.; Fast, M.D. Effects of orally administered immunostimulants on inflammatory gene expression and sea lice (Lepeophtheirus salmonis) burdens on Atlantic salmon (Salmo salar). Aquaculture 2012, 366, 9–16. [Google Scholar] [CrossRef]
- Carrington, A.C.; Secombes, C.J. A review of CpGs and their relevance to aquaculture. Vet. Immunol. Immunopathol. 2006, 112, 87–101. [Google Scholar] [CrossRef]
- Cuesta, A.; Esteban, M.A.; Meseguer, J. The expression profile of TLR9 mRNA and CpG ODNs immunostimulatory actions in the teleost gilthead seabream points to a major role of lymphocytes. Cell. Mol. Life Sci. 2008, 65, 2091–2104. [Google Scholar] [CrossRef]
- Liu, C.-s.; Sun, Y.; Hu, Y.-h.; Sun, L. Identification and analysis of a CpG motif that protects turbot (Scophthalmus maximus) against bacterial challenge and enhances vaccine-induced specific immunity. Vaccine 2010, 28, 4153–4161. [Google Scholar] [CrossRef]
- Cárdenas-Reyna, T.; Angulo, C.; Hori-Oshima, S.; Velázquez-Lizárraga, E.; Reyes-Becerril, M. B-cell activating CpG ODN 1668 enhance the immune response of Pacific red snapper (Lutjanus peru) exposed to Vibrio parahaemolitycus. Dev. Comp. Immunol. 2016, 62, 72–81. [Google Scholar] [CrossRef]
- Jung, M.-H.; Jung, S.-J. CpG ODN 1668 induce innate and adaptive immune responses in rock bream (Oplegnathus fasciatus) against rock bream iridovirus (RBIV) infection. Fish Shellfish Immunol. 2017, 69, 247–257. [Google Scholar] [CrossRef]
- Purcell, S.L.; Friend, S.E.; Covello, J.M.; Donkin, A.; Groman, D.B.; Poley, J.; Fast, M.D. CpG inclusion in feed reduces sea lice, Lepeophtheirus salmonis, numbers following re-infection. J. Fish. Dis. 2013, 36, 229–240. [Google Scholar] [CrossRef]
- Chen, J.; Li, C.; Huang, R.; Du, F.; Liao, L.; Zhu, Z.; Wang, Y. Transcriptome analysis of head kidney in grass carp and discovery of immune-related genes. BMC Vet. Res. 2012, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Press, C.M.; Evensen, Ø. The morphology of the immune system in teleost fishes. Fish Shellfish Immunol. 1999, 9, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Caballero-Solares, A.; Xue, X.; Parrish, C.C.; Foroutani, M.B.; Taylor, R.G.; Rise, M.L. Changes in the liver transcriptome of farmed Atlantic salmon (Salmo salar) fed experimental diets based on terrestrial alternatives to fish meal and fish oil. BMC Genom. 2018, 19, 796. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Rehmsmeier, M.; Steffen, P.; Höchsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Johansen, I.; Andreassen, R. Validation of miRNA genes suitable as reference genes in qPCR analyses of miRNA gene expression in Atlantic salmon (Salmo salar). BMC Res. Notes 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booman, M.; Xu, Q.H.; Rise, M.L. Evaluation of the impact of camelina oil-containing diets on the expression of genes involved in the innate anti-viral immune response in Atlantic cod (Gadus morhua). Fish Shellfish Immunol. 2014, 41, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Hixson, S.M.; Hori, T.S.; Booman, M.; Parrish, C.C.; Anderson, D.M.; Rise, M.L. Atlantic salmon (Salmo salar) liver transcriptome response to diets containing Camelina sativa products. Comp. Biochem. Physiol. Part D Genom. Proteom. 2015, 14, 1–15. [Google Scholar] [CrossRef]
- Chu, Q.; Sun, Y.; Bi, D.; Cui, J.; Xu, T. Up-regulated of miR-8159-5p and miR-217-5p by LPS stimulation negatively co-regulate TLR1 in miiuy croaker. Dev. Comp. Immunol. 2017, 67, 117–125. [Google Scholar] [CrossRef]
- Chu, Q.; Sun, Y.; Cui, J.; Xu, T. MicroRNA-3570 modulates the NF-κB pathway in teleost fish by targeting MyD88. J. Immunol. 2017, 198, 3274–3282. [Google Scholar] [CrossRef] [Green Version]
- Gong, G.; Sha, Z.; Chen, S.; Li, C.; Yan, H.; Chen, Y.; Wang, T. Expression profiling analysis of the microRNA response of Cynoglossus semilaevis to Vibrio anguillarum and other stimuli. Mar. Biotechnol. 2015, 17, 338–352. [Google Scholar] [CrossRef]
- Woldemariam, N.T.; Agafonov, O.; Sindre, H.; Høyheim, B.; Houston, R.D.; Robledo, D.; Bron, J.E.; Andreassen, R. miRNAs predicted to regulate host anti-viral gene pathways in IPNV-challenged Atlantic salmon are affected by viral load, and associated with the major IPN resistance QTL genotypes in late infection. Front. Immunol. 2019. Submitted. [Google Scholar]
- Buck, A.H.; Perot, J.; Chisholm, M.A.; Kumar, D.S.; Tuddenham, L.; Cognat, V.; Marcinowski, L.; Dölken, L.; Pfeffer, S. Post-transcriptional regulation of miR-27 in murine cytomegalovirus infection. RNA 2010, 16, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.H.; He, X.P.; Bai, Z.Y.; Yue, G.H. Identification and characterization of 63 MicroRNAs in the Asian seabass Lates calcarifer. PLoS ONE 2011, 6, e17537. [Google Scholar] [CrossRef]
- Ordas, A.; Kanwal, Z.; Lindenberg, V.; Rougeot, J.; Mink, M.; Spaink, H.P.; Meijer, A.H. MicroRNA-146 function in the innate immune transcriptome response of zebrafish embryos to Salmonella typhimurium infection. BMC Genom. 2013, 14, 696. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Gan, Z.; Cai, S.; Wang, Z.; Yu, D.; Lin, Z.; Lu, Y.; Wu, Z.; Jian, J. Comprehensive identification and profiling of Nile tilapia (Oreochromis niloticus) microRNAs response to Streptococcus agalactiae infection through high-throughput sequencing. Fish Shellfish Immunol. 2016, 54, 93–106. [Google Scholar] [CrossRef]
- Ma, F.; Xu, S.; Liu, X.; Zhang, Q.; Xu, X.; Liu, M.; Hua, M.; Li, N.; Yao, H.; Cao, X. The microRNA miR-29 controls innate and adaptive immune responses to intracellular bacterial infection by targeting interferon-γ. Nat. Immunol. 2011, 12, 861. [Google Scholar] [CrossRef]
- Pedersen, I.; David, M. MicroRNAs in the immune response. Cytokine 2008, 43, 391–394. [Google Scholar] [CrossRef] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Chang, K.-J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [Green Version]
- Najib, A.; Kim, M.S.; Choi, S.H.; Kang, Y.J.; Kim, K.H. Changes in microRNAs expression profile of olive flounder (Paralichthys olivaceus) in response to viral hemorrhagic septicemia virus (VHSV) infection. Fish Shellfish Immunol. 2016, 51, 384–391. [Google Scholar] [CrossRef]
- Sha, Z.; Gong, G.; Wang, S.; Lu, Y.; Wang, L.; Wang, Q.; Chen, S. Identification and characterization of Cynoglossus semilaevis microRNA response to Vibrio anguillarum infection through high-throughput sequencing. Dev. Comp. Immunol. 2014, 44, 59–69. [Google Scholar] [CrossRef]
- Galardi, S.; Mercatelli, N.; Farace, M.G.; Ciafre, S.A. NF-kB and c-Jun induce the expression of the oncogenic miR-221 and miR-222 in prostate carcinoma and glioblastoma cells. Nucleic Acids Res. 2011, 39, 3892–3902. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Zhao, P.; Zhang, C.; Fu, Z.; Chen, Y.; Lu, A.; Liu, N.; You, Y.; Pu, P.; Kang, C. Analysis of miR-221 and p27 expression in human gliomas. Mol. Med. Rep. 2009, 2, 651–656. [Google Scholar]
- Li, J.; Wang, Y.; Yu, W.; Chen, J.; Luo, J. Expression of serum miR-221 in human hepatocellular carcinoma and its prognostic significance. Biochem. Biophys. Res. Commun. 2011, 406, 70–73. [Google Scholar] [CrossRef]
- Yan, H.; Chen, Y.; Zhou, S.; Li, C.; Gong, G.; Chen, X.; Wang, T.; Chen, S.; Sha, Z. Expression profile analysis of miR-221 and miR-222 in different tissues and head kidney cells of Cynoglossus semilaevis, following pathogen infection. Mar. Biotechnol. 2016, 18, 37–48. [Google Scholar] [CrossRef]
- Sun, X.; Sit, A.; Feinberg, M.W. Role of miR-181 family in regulating vascular inflammation and immunity. Trends Cardiovasc. Med. 2014, 24, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.-c.; Zhang, J.; Sun, L. In-depth profiling and analysis of host and viral microRNAs in Japanese flounder (Paralichthys olivaceus) infected with megalocytivirus reveal involvement of microRNAs in host-virus interaction in teleost fish. BMC Genom. 2014, 15, 878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schyth, B.D.; Bela-ong, D.B.; Jalali, S.A.H.; Kristensen, L.B.J.; Einer-Jensen, K.; Pedersen, F.S.; Lorenzen, N. Two virus-induced MicroRNAs known only from teleost fishes are orthologues of MicroRNAs involved in cell cycle control in humans. PLoS ONE 2015, 10, e0132434. [Google Scholar] [CrossRef] [PubMed]
- Yuhong, J.; Leilei, T.; Fuyun, Z.; Hongyang, J.; Xiaowen, L.; Liying, Y.; Lei, Z.; Jingrong, M.; Jinpeng, Y. Identification and characterization of immune-related microRNAs in blunt snout bream, Megalobrama amblycephala. Fish Shellfish Immunol. 2016, 49, 470–492. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-P.; Xue, W.; Wang, J.-T.; Wan, Y.-M.; Wang, S.-L.; Xu, P.; Zhang, Y.; Li, J.-T.; Sun, X.-W. Identification of common carp (Cyprinus carpio) microRNAs and microRNA-related SNPs. BMC Genom. 2012, 13, 413. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.J.; Kim, K.H. Effect of CpG-ODNs belonging to different classes on resistance of olive flounder (Paralichthys olivaceus) against viral hemorrhagic septicemia virus (VHSV) and Miamiensis avidus (Ciliata; Scuticociliatia) infections. Aquaculture 2012, 324, 39–43. [Google Scholar] [CrossRef]
- Kerkmann, M.; Rothenfusser, S.; Hornung, V.; Towarowski, A.; Wagner, M.; Sarris, A.; Giese, T.; Endres, S.; Hartmann, G. Activation with CpG-A and CpG-B oligonucleotides reveals two distinct regulatory pathways of type I IFN synthesis in human plasmacytoid dendritic cells. J. Immunol. 2003, 170, 4465–4474. [Google Scholar] [CrossRef] [Green Version]
- Strandskog, G.; Ellingsen, T.; Jørgensen, J.B. Characterization of three distinct CpG oligonucleotide classes which differ in ability to induce IFN α/β activity and cell proliferation in Atlantic salmon (Salmo salar L.) leukocytes. Dev. Comp. Immunol. 2007, 31, 39–51. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID 1 | Total Number of Raw Reads 2 | Trimmed and Filtered Reads 3 | Reads Mapped to miRNAs (%) 4 | Accession Number 5 |
|---|---|---|---|---|
| 1-PBS-T30-2 | 9,609,300 | 4,800,941 | 89.1 | SRR9709006 |
| 2-PBS-T30-3 | 15,384,722 | 9,966,411 | 90.4 | SRR9709007 |
| 3-PBS-T33-3 | 12,674,361 | 8,124,580 | 89.9 | SRR9709008 |
| 4-ASAL-T33-1 | 11,715,675 | 7,190,332 | 89.4 | SRR9709009 |
| 5-ASAL-T30-4 | 14,028,188 | 9,134,360 | 89.7 | SRR9709002 |
| 6-ASAL-T33-3 | 34,491,682 | 8,654,401 | 91.3 | SRR9709003 |
| 7-pIC-T33-2 | 18,600,224 | 8,519,434 | 80.6 | SRR9709004 |
| 8-pIC-T30-3 | 13,957,705 | 9,864,824 | 93.4 | SRR9709005 |
| 9-pIC-T33-3 | 16,877,729 | 5,453,442 | 68.2 | SRR9709010 |
| miRNAs 1 | Base Mean 2 | Fold-Change 3 | Adjusted p-Values 4 | qPCR Fold-Change 5 |
|---|---|---|---|---|
| Up-regulated by pIC | ||||
| miR-27d-1-5p 6 | 868.11 | 2.17 | 0.049 | 2.69 |
| miR-27d-2-5p 6 | 861.10 | 2.17 | 0.049 | 2.69 |
| miR-30e-1-2-3p | 404.73 | 2.16 | 0.049 | 2.57 |
| miR-135bd-5p | 52.80 | 14.03 | 0.036 | 1.54 |
| miR-146a-5p | 152066.51 | 1.71 | 0.028 | 1.62 |
| miR-146a-1-2-3p | 6322.03 | 7.94 | 3.85 × 10-4 | 7.89 |
| miR-181a-5-3p | 11080.58 | 1.59 | 0.051 | 1.97 |
| miR-221-5p | 221.52 | 4.44 | 0.036 | 7.55 |
| miR-462a-3p | 1554.09 | 2.69 | 0.098 | 5.72 |
| miR-462b-3p | 941.82 | 5.46 | 8.0 × 10-12 | N/A |
| miR-8159-5p | 97.72 | 9.92 | 0.055 | N/A |
| Down-regulated by pIC | ||||
| miR-106a-3p | 960.11 | −1.93 | 0.036 | N/A |
| miRNAs 1 | Base Mean 2 | Fold-Change 3 | Adjusted p-Values 4 | qPCR Fold-Change 5 |
|---|---|---|---|---|
| Up-regulated by ASAL | ||||
| miR-21a-1-3p | 267.99 | 1.52 | 0.099 | N/A |
| miR-29b-2-5p | 516.80 | 1.55 | 0.091 | 1.47 |
| miR-146a-5p | 202688.90 | 2.01 | 8.9 × 10−6 | 1.89 |
| miR-146a-1-2-3p | 8503.53 | 9.06 | 9.06 × 10−37 | 5.87 |
| miR-146a-3-3p | 12146.14 | 3.43 | 2.62 × 10−11 | N/A |
| miR-146d-1-3p | 421.43 | 2.60 | 2.45 × 10−4 | N/A |
| miR-183-1-3-3p | 19.55 | 4.17 | 0.018 | N/A |
| miR-183-2-3p | 19.24 | 4.08 | 0.021 | N/A |
| miR-192a-5p | 391.09 | 6.68 | 0.016 | 1.64 |
| miR-194a-5p | 82.25 | 6.68 | 0.016 | 1.37 |
| miR-200b-3p | 275.30 | 8.00 | 0.009 | N/A |
| miR-221-5p | 141.35 | 1.92 | 0.018 | 2.23 |
| miR-429ab-3p | 11.34 | 8.40 | 0.071 | N/A |
| miR-725-5p | 26.55 | 17.15 | 2.27 × 10−6 | 1.55 |
| miR-725-3p | 163.89 | 4.63 | 6.75 × 10−6 | N/A |
| miR-novel-16-5p | 1066.67 | 2.01 | 1.81 × 10−4 | 1.09 |
| Down-regulated by ASAL | ||||
| miR-722-3p | 410.45 | −2.08 | 0.021 | −1.64 |
| miR-727a-3p | 886.63 | −1.82 | 0.085 | −1.94 |
| Pathway | Name | Assigned Target Genes of pIC-Responsive miRNAs 1 | Assigned Target Genes of ASAL-Responsive miRNAs 2 |
|---|---|---|---|
| sasa04621 | NOD-like receptor signaling pathway | 8 | 12 |
| sasa04060 | Cytokine-cytokine receptor interaction | 7 | 13 |
| sasa04217 | Necroptosis | 7 | 11 |
| sasa04620 | Toll-like receptor signaling pathway | 6 | 10 |
| sasa04210 | Apoptosis | 6 | 7 |
| sasa04625 | C-type lectin receptor signaling pathway | 5 | 5 |
| sasa04622 | RIG-I-like receptor signaling pathway | 4 | 7 |
| sasa04514 | Cell adhesion molecules (CAMs) | 4 | 2 |
| Control vs. CpG 3 | ||||
|---|---|---|---|---|
| Pre-Injection | PBS | pIC | ASAL | |
| p (perm) 1 | 0.0015 | 0.0041 | 0.4461 | 0.0015 |
| Average dissimilarity (%) 2 | 17.52 | 19.00 | - | 26.81 |
| Contributing variables (top 70%) | miR-722-3p | miR-194a-5p | - | miR-146a-1-2-3p |
| miR-192a-5p | miR-725-5p | - | miR-192a-5p | |
| miR-462a-3p | miR-192a-5p | - | miR-221-5p | |
| miR-727a-3p | miR-722-3p | - | miR-725-5p | |
| miR-194a-5p | miR-27d-1-2-5p | - | miR-29b-2-5p | |
| miR-725-5p | miR-727a-3p | - | miR-194a-5p | |
| miR-181a-5-3p | miR-146a-1-2-3p | - | miR-27d-1-2-5p | |
| miR-135bd-5p | miR-novel-16-5p | - | - | |
| - | miR-181a-5-3p | - | - | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, X.; Woldemariam, N.T.; Caballero-Solares, A.; Umasuthan, N.; Fast, M.D.; Taylor, R.G.; Rise, M.L.; Andreassen, R. Dietary Immunostimulant CpG Modulates MicroRNA Biomarkers Associated with Immune Responses in Atlantic Salmon (Salmo salar). Cells 2019, 8, 1592. https://doi.org/10.3390/cells8121592
Xue X, Woldemariam NT, Caballero-Solares A, Umasuthan N, Fast MD, Taylor RG, Rise ML, Andreassen R. Dietary Immunostimulant CpG Modulates MicroRNA Biomarkers Associated with Immune Responses in Atlantic Salmon (Salmo salar). Cells. 2019; 8(12):1592. https://doi.org/10.3390/cells8121592
Chicago/Turabian StyleXue, Xi, Nardos Tesfaye Woldemariam, Albert Caballero-Solares, Navaneethaiyer Umasuthan, Mark D. Fast, Richard G. Taylor, Matthew L. Rise, and Rune Andreassen. 2019. "Dietary Immunostimulant CpG Modulates MicroRNA Biomarkers Associated with Immune Responses in Atlantic Salmon (Salmo salar)" Cells 8, no. 12: 1592. https://doi.org/10.3390/cells8121592
APA StyleXue, X., Woldemariam, N. T., Caballero-Solares, A., Umasuthan, N., Fast, M. D., Taylor, R. G., Rise, M. L., & Andreassen, R. (2019). Dietary Immunostimulant CpG Modulates MicroRNA Biomarkers Associated with Immune Responses in Atlantic Salmon (Salmo salar). Cells, 8(12), 1592. https://doi.org/10.3390/cells8121592