Multi-Omics Integration Reveals Short and Long-Term Effects of Gestational Hypoxia on the Heart Development



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Proteomics Analysis
2.3. Metabolomics Analysis
2.4. Bioinformatics and Statistics
3. Results
3.1. Study Design
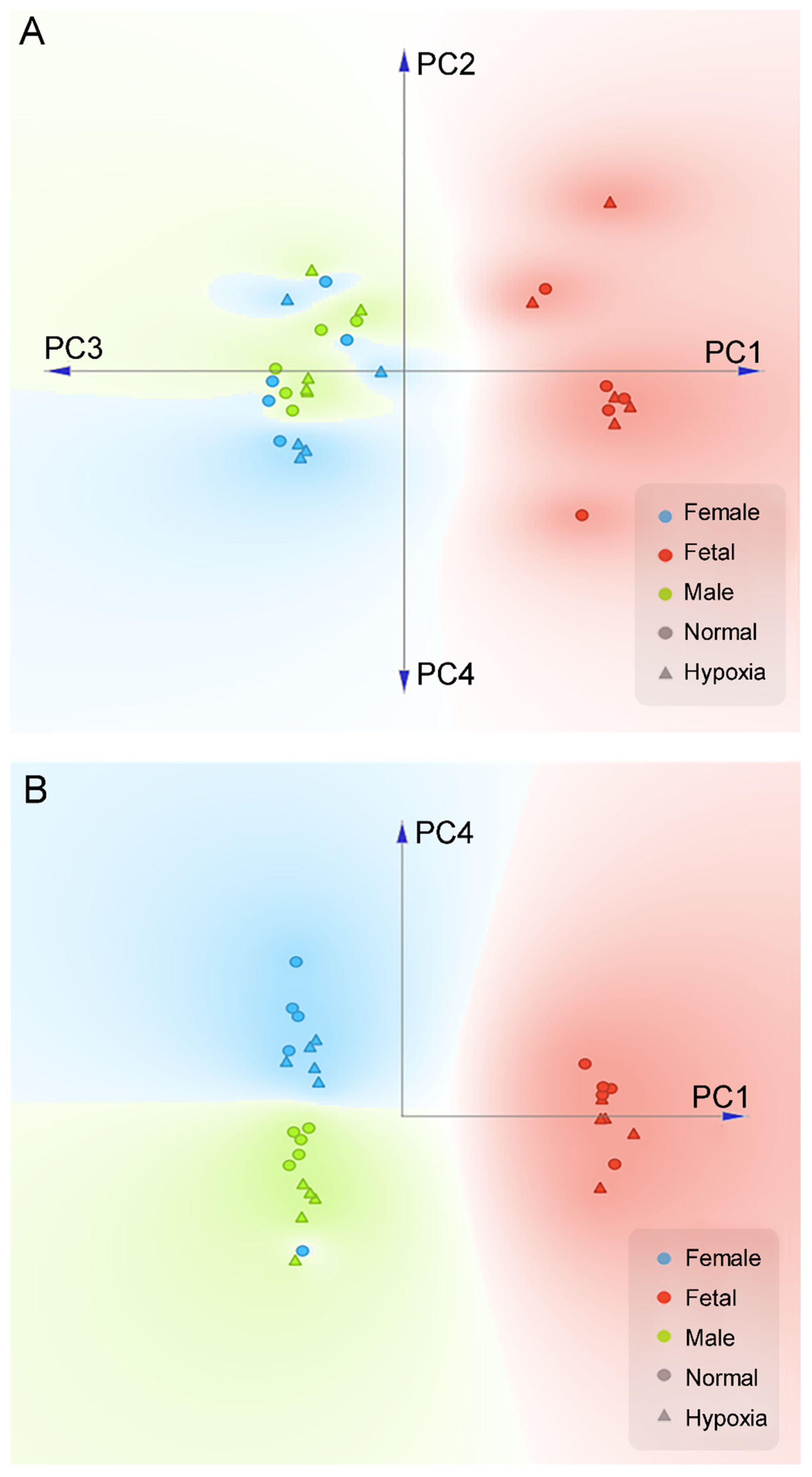
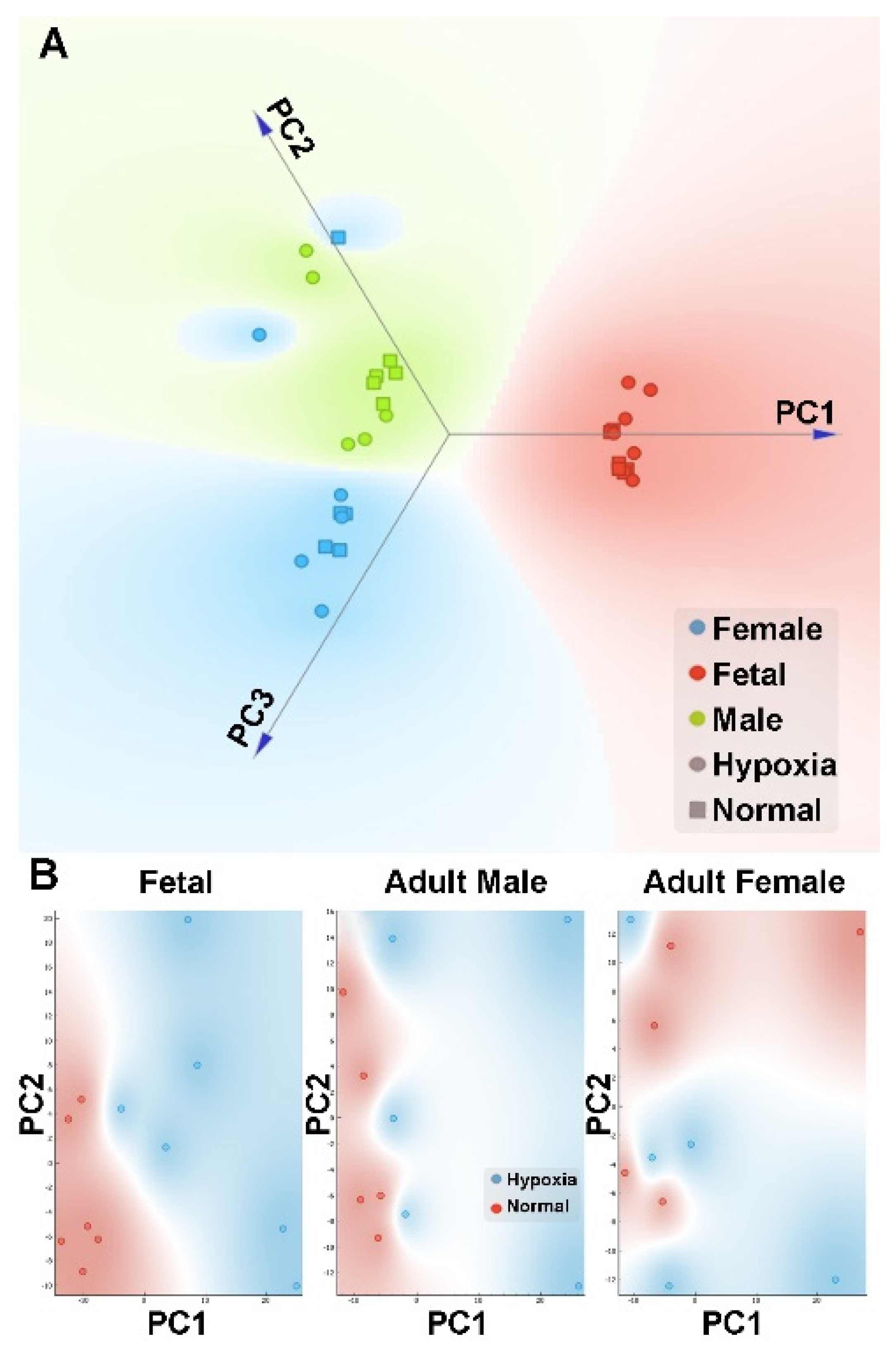
3.2. Effect of Antenatal Hypoxia on Proteome in the Heart
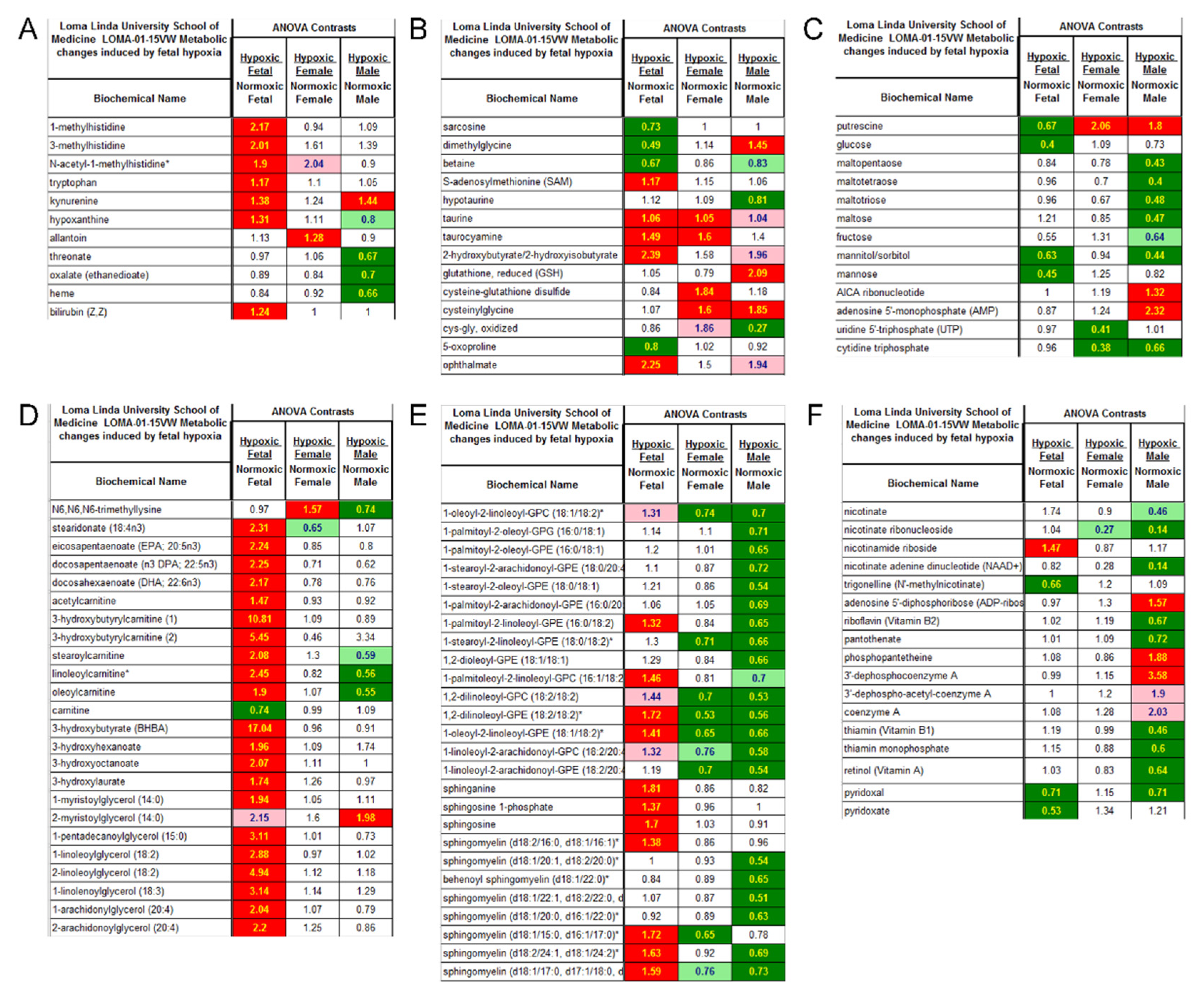
3.3. Effect of Antenatal Hypoxia on Metabolome in the Heart
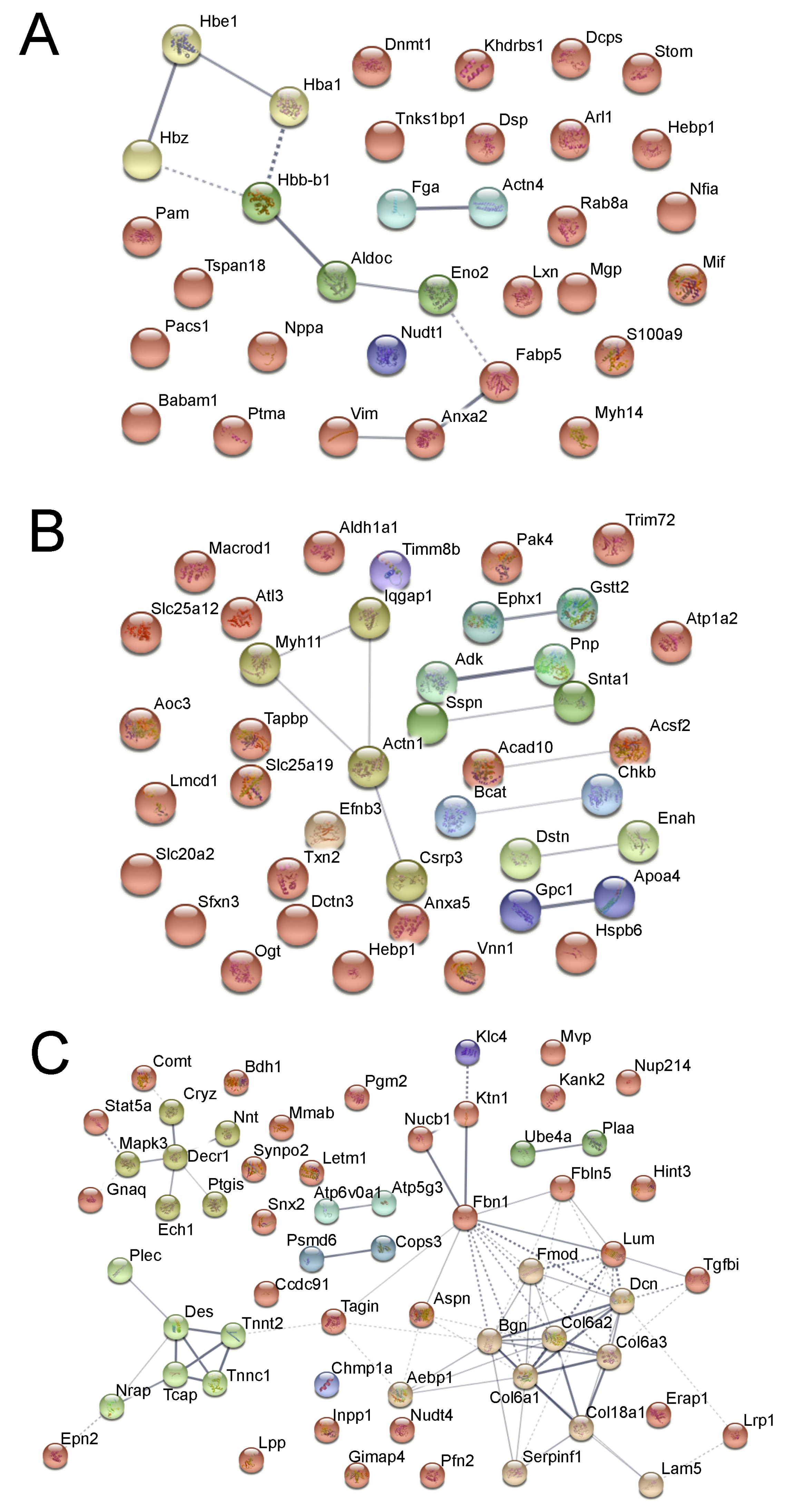
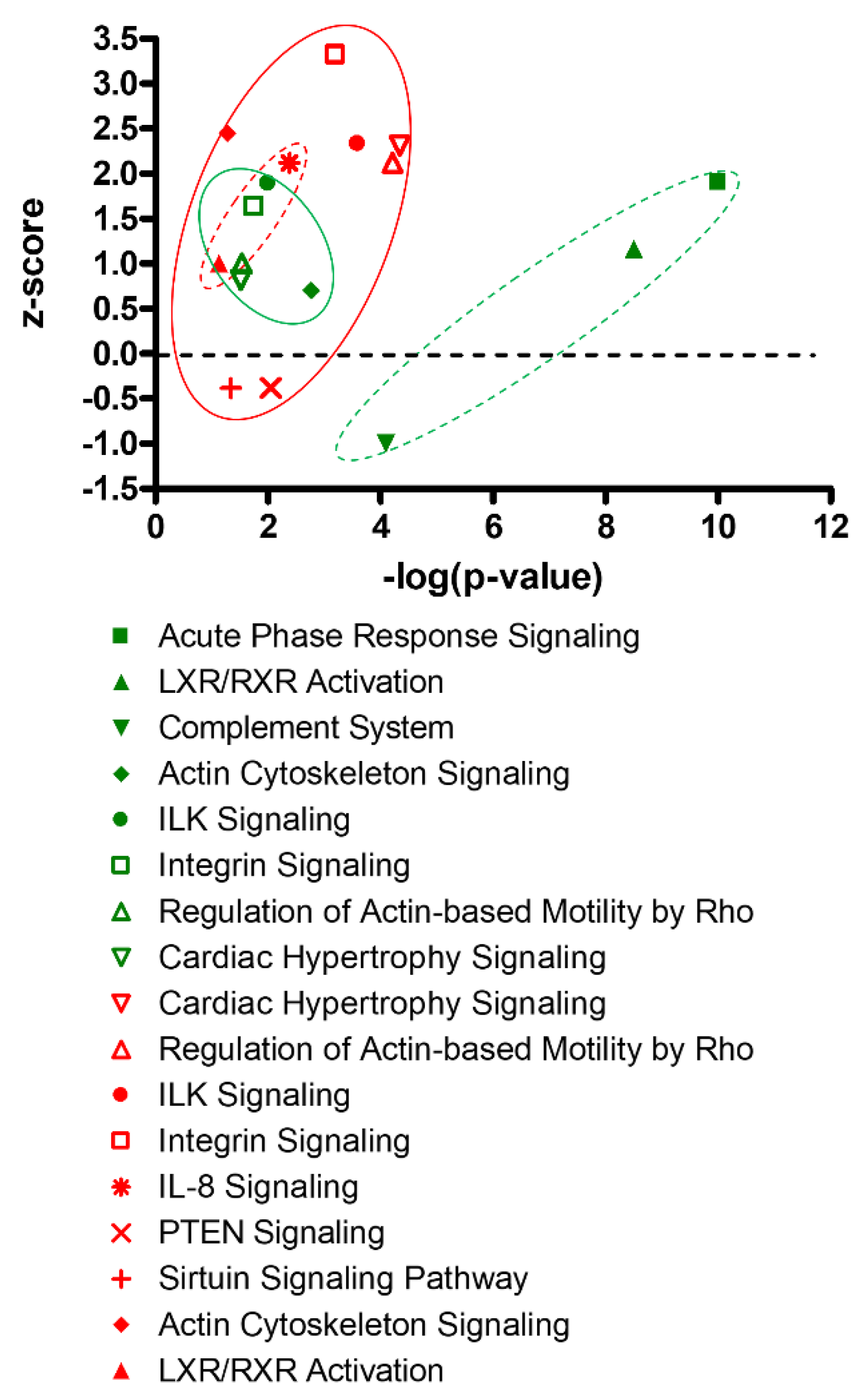
3.4. Integrated Multi-Omics Data Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barker, D.J.; Osmond, C. Infant mortality, childhood nutrition, and ischaemic heart disease in England and Wales. Lancet 1986, 1, 1077–1081. [Google Scholar] [CrossRef]
- Bateson, P.; Barker, D.; Clutton-Brock, T.; Deb, D.; D’Udine, B.; Foley, R.A.; Gluckman, P.; Godfrey, K.; Kirkwood, T.; Lahr, M.M.; et al. Developmental plasticity and human health. Nature 2004, 430, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in utero and early-life conditions on adult health and disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillen, I.C.; Robinson, J.S. Developmental origins of the metabolic syndrome: Prediction, plasticity, and programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef]
- Ducsay, C.A.; Goyal, R.; Pearce, W.J.; Wilson, S.; Hu, X.Q.; Zhang, L. Gestational Hypoxia and Developmental Plasticity. Physiol. Rev. 2018, 98, 1241–1334. [Google Scholar] [CrossRef]
- Heydeck, D.; Roigas, J.; Roigas, C.; Papies, B.; Lun, A. The catecholamine sensitivity of adult rats is enhanced after prenatal hypoxia. Biol. Neonate 1994, 66, 106–111. [Google Scholar] [CrossRef]
- Roigas, J.; Roigas, C.; Heydeck, D.; Papies, B. Prenatal hypoxia alters the postnatal development of beta-adrenoceptors in the rat myocardium. Biol. Neonate 1996, 69, 383–388. [Google Scholar] [CrossRef]
- Butler, T.G.; Schwartz, J.; McMillen, I.C. Differential effects of the early and late intrauterine environment on corticotrophic cell development. J. Clin. Investig. 2002, 110, 783–791. [Google Scholar] [CrossRef]
- Peyronnet, J.; Dalmaz, Y.; Ehrstrom, M.; Mamet, J.; Roux, J.C.; Pequignot, J.M.; Thoren, H.P.; Lagercrantz, H. Long-lasting adverse effects of prenatal hypoxia on developing autonomic nervous system and cardiovascular parameters in rats. Pflug. Arch. 2002, 443, 858–865. [Google Scholar] [CrossRef]
- Davis, L.; Roullet, J.B.; Thornburg, K.L.; Shokry, M.; Hohimer, A.R.; Giraud, G.D. Augmentation of coronary conductance in adult sheep made anaemic during fetal life. J. Physiol. 2003, 547, 53–59. [Google Scholar] [CrossRef]
- Jones, R.D.; Morice, A.H.; Emery, C.J. Effects of perinatal exposure to hypoxia upon the pulmonary circulation of the adult rat. Physiol. Res. 2004, 53, 11–17. [Google Scholar] [PubMed]
- Mone, S.M.; Gillman, M.W.; Miller, T.L.; Herman, E.H.; Lipshultz, S.E. Effects of environmental exposures on the cardiovascular system: Prenatal period through adolescence. Pediatrics 2004, 113, 1058–1069. [Google Scholar] [PubMed]
- Williams, S.J.; Hemmings, D.G.; Mitchell, J.M.; McMillen, I.C.; Davidge, S.T. Effects of maternal hypoxia or nutrient restriction during pregnancy on endothelial function in adult male rat offspring. J. Physiol. 2005, 565, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Ream, M.; Ray, A.M.; Chandra, R.; Chikaraishi, D.M. Early fetal hypoxia leads to growth restriction and myocardial thinning. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R583–R595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camm, E.J.; Hansell, J.A.; Kane, A.D.; Herrera, E.A.; Lewis, C.; Wong, S.; Morrell, N.W.; Giussani, D.A. Partial contributions of developmental hypoxia and undernutrition to prenatal alterations in somatic growth and cardiovascular structure and function. Am. J. Obs. Gynecol. 2010, 203, 495-e24. [Google Scholar] [CrossRef]
- Giussani, D.A.; Camm, E.J.; Niu, Y.; Richter, H.G.; Blanco, C.E.; Gottschalk, R.; Blake, E.Z.; Horder, K.A.; Thakor, A.S.; Hansell, J.A.; et al. Developmental programming of cardiovascular dysfunction by prenatal hypoxia and oxidative stress. PLoS ONE 2012, 7, e31017. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Xiao, Y.; Estrella, J.L.; Ducsay, C.A.; Gilbert, R.D.; Zhang, L. Effect of fetal hypoxia on heart susceptibility to ischemia and reperfusion injury in the adult rat. J. Soc. Gynecol. Investig. 2003, 10, 265–274. [Google Scholar] [CrossRef]
- Li, G.; Bae, S.; Zhang, L. Effect of prenatal hypoxia on heat stress-mediated cardioprotection in adult rat heart. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H1712–H1719. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Williams, S.J.; O’Brien, D.; Davidge, S.T. Hypoxia or nutrient restriction during pregnancy in rats leads to progressive cardiac remodeling and impairs postischemic recovery in adult male offspring. Faseb J. 2006, 20, 1251–1253. [Google Scholar] [CrossRef]
- Xue, Q.; Zhang, L. Prenatal hypoxia causes a sex-dependent increase in heart susceptibility to ischemia and reperfusion injury in adult male offspring: Role of protein kinase C epsilon. J. Pharm. Exp. 2009, 330, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Patterson, A.J.; Chen, M.; Xue, Q.; Xiao, D.; Zhang, L. Chronic prenatal hypoxia induces epigenetic programming of PKC {epsilon} gene repression in rat hearts. Circ. Res. 2010, 107, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Xue, Q.; Dasgupta, C.; Chen, M.; Zhang, L. Foetal hypoxia increases cardiac AT(2)R expression and subsequent vulnerability to adult ischaemic injury. Cardiovasc. Res. 2011, 89, 300–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, A.J.; Xiao, D.; Xiong, F.; Dixon, B.; Zhang, L. Hypoxia-derived oxidative stress mediates epigenetic repression of PKCepsilon gene in foetal rat hearts. Cardiovasc. Res. 2012, 93, 302–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, F.; Lin, T.; Song, M.; Ma, Q.; Martinez, S.R.; Lv, J.; MataGreenwood, E.; Xiao, D.; Xu, Z.; Zhang, L. Antenatal hypoxia induces epigenetic repression of glucocorticoid receptor and promotes ischemic-sensitive phenotype in the developing heart. J. Mol. Cell Cardiol. 2016, 91, 160–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, J.; Ma, Q.; Dasgupta, C.; Xu, Z.; Zhang, L. Antenatal Hypoxia and Programming of Glucocorticoid Receptor Expression in the Adult Rat Heart. Front. Physiol. 2019, 10, 323. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, X.; Dasgupta, C.; Chen, W.; Song, R.; Wang, C.; Zhang, L. Foetal hypoxia impacts methylome and transcriptome in developmental programming of heart disease. Cardiovasc. Res. 2019, 115, 1306–1319. [Google Scholar] [CrossRef]
- Xiao, D.; Dasgupta, C.; Chen, M.; Zhang, K.; Buchholz, J.; Xu, Z.; Zhang, L. Inhibition of DNA methylation reverses norepinephrine-induced cardiac hypertrophy in rats. Cardiovasc. Res. 2014, 101, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Drakakaki, G.; van de Ven, W.; Pan, S.; Miao, Y.; Wang, J.; Keinath, N.F.; Weatherly, B.; Jiang, L.; Schumacher, K.; Hicks, G.; et al. Isolation and proteomic analysis of the SYP61 compartment reveal its role in exocytic trafficking in Arabidopsis. Cell Res. 2012, 22, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Park, S.K.; Venable, J.D.; Wohlschlegel, J.A.; Diedrich, J.K.; Cociorva, D.; Lu, B.; Liao, L.; Hewel, J.; Han, X.; et al. ProLuCID: An improved SEQUEST-like algorithm with enhanced sensitivity and specificity. J. Proteom. 2015, 129, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Park, S.K.; Aslanian, A.; McClatchy, D.B.; Han, X.; Shah, H.; Singh, M.; Rauniyar, N.; Moresco, J.J.; Pinto, A.F.; Diedrich, J.K.; et al. Census 2: Isobaric labeling data analysis. Bioinformatics 2014, 30, 2208–2209. [Google Scholar] [CrossRef] [Green Version]
- Noble, W.S. Mass spectrometrists should search only for peptides they care about. Nat. Methods 2015, 12, 605–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subhash, S.; Kanduri, C. GeneSCF: A real-time based functional enrichment tool with support for multiple organisms. Bmc Bioinform. 2016, 17, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, M.P.; Wang, D.; Lau, E.; Liem, D.A.; Kim, A.K.; Ng, D.C.; Liang, X.; Bleakley, B.J.; Liu, C.; Tabaraki, J.D.; et al. Protein kinetic signatures of the remodeling heart following isoproterenol stimulation. J. Clin. Investig. 2014, 124, 1734–1744. [Google Scholar] [CrossRef] [Green Version]
- Griffin, M.; Lee, H.W.; Zhao, L.; Eghbali-Webb, M. Gender-related differences in proliferative response of cardiac fibroblasts to hypoxia: Effects of estrogen. Mol. Cell Biochem. 2000, 215, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.J.; Berg, H.M.; Easton, C.J.; Bakker, A.J. The effect of taurine depletion on the contractile properties and fatigue in fast-twitch skeletal muscle of the mouse. Amino Acids 2006, 31, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Lake, N. Loss of cardiac myofibrils: Mechanism of contractile deficits induced by taurine deficiency. Am. J. Physiol. 1993, 264, H1323–H1326. [Google Scholar] [CrossRef]
- Gall, W.E.; Beebe, K.; Lawton, K.A.; Adam, K.P.; Mitchell, M.W.; Nakhle, P.J.; Ryals, J.A.; Milburn, M.V.; Nannipieri, M.; Camastra, S.; et al. alpha-hydroxybutyrate is an early biomarker of insulin resistance and glucose intolerance in a nondiabetic population. PLoS ONE 2010, 5, e10883. [Google Scholar] [CrossRef] [Green Version]
- Dello, S.A.; Neis, E.P.; de Jong, M.C.; van Eijk, H.M.; Kicken, C.H.; Olde Damink, S.W.; Dejong, C.H. Systematic review of ophthalmate as a novel biomarker of hepatic glutathione depletion. Clin. Nutr. 2013, 32, 325–330. [Google Scholar] [CrossRef]
- Hunter, R.W.; Treebak, J.T.; Wojtaszewski, J.F.; Sakamoto, K. Molecular mechanism by which AMP-activated protein kinase activation promotes glycogen accumulation in muscle. Diabetes 2011, 60, 766–774. [Google Scholar] [CrossRef] [Green Version]
- Arad, M.; Seidman, C.E.; Seidman, J.G. AMP-activated protein kinase in the heart: Role during health and disease. Circ. Res. 2007, 100, 474–488. [Google Scholar] [CrossRef] [Green Version]
- Novak, F.; Kolar, F.; Hamplova, B.; Mrnka, L.; Pelouch, V.; Ostadal, B.; Novakova, O. Myocardial phospholipid remodeling under different types of load imposed during early postnatal development. Physiol. Res. 2009, 58 (Suppl. 2), S13–S32. [Google Scholar] [PubMed]
- Karliner, J.S. Sphingosine kinase and sphingosine 1-phosphate in the heart: A decade of progress. Biochim Biophys. Acta 2013, 1831, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, M. Cardioprotective role of sphingosine-1-phosphate. J. Physiol. Pharm. 2011, 62, 601–607. [Google Scholar]
- Hakimi, A.A.; Reznik, E.; Lee, C.H.; Creighton, C.J.; Brannon, A.R.; Luna, A.; Aksoy, B.A.; Liu, E.M.; Shen, R.; Lee, W.; et al. An Integrated Metabolic Atlas of Clear Cell Renal Cell Carcinoma. Cancer Cell 2016, 29, 104–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, R.L.; Weber, R.J.; Liu, H.; Sharma-Oates, A.; Viant, M.R. Galaxy-M: A Galaxy workflow for processing and analyzing direct infusion and liquid chromatography mass spectrometry-based metabolomics data. Gigascience 2016, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Villiers, F.; Ducruix, C.; Hugouvieux, V.; Jarno, N.; Ezan, E.; Garin, J.; Junot, C.; Bourguignon, J. Investigating the plant response to cadmium exposure by proteomic and metabolomic approaches. Proteomics 2011, 11, 1650–1663. [Google Scholar] [CrossRef]
- Zhang, W.; Li, F.; Nie, L. Integrating multiple ‘omics’ analysis for microbial biology: Application and methodologies. Microbiology 2010, 156, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Xiao, Y.; Li, G.; Casiano, C.A.; Zhang, L. Effect of maternal chronic hypoxic exposure during gestation on apoptosis in fetal rat heart. Am. J. Physiol Heart Circ. Physiol. 2003, 285, H983–H990. [Google Scholar] [CrossRef] [Green Version]
- Tong, W.; Xue, Q.; Li, Y.; Zhang, L. Maternal hypoxia alters matrix metalloproteinase expression patterns and causes cardiac remodeling in fetal and neonatal rats. Am. J. Physiol Heart Circ. Physiol. 2011, 301, H2113–H2121. [Google Scholar] [CrossRef] [Green Version]
- Tong, W.; Xiong, F.; Li, Y.; Zhang, L. Hypoxia inhibits cardiomyocyte proliferation in fetal rat hearts via upregulating TIMP-4. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R613–R620. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L. Prenatal hypoxia and cardiac programming. J. Soc. Gynecol. Investig. 2005, 12, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Rueda-Clausen, C.F.; Morton, J.S.; Lopaschuk, G.D.; Davidge, S.T. Long-term effects of intrauterine growth restriction on cardiac metabolism and susceptibility to ischaemia/reperfusion. Cardiovasc Res. 2011, 90, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, D.; Ducsay, C.A.; Zhang, L. Chronic hypoxia and developmental regulation of cytochrome c expression in rats. J. Soc. Gynecol. Investig. 2000, 7, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Al-Hasan, Y.M.; Pinkas, G.A.; Thompson, L.P. Prenatal Hypoxia Reduces Mitochondrial Protein Levels and Cytochrome c Oxidase Activity in Offspring Guinea Pig Hearts. Reprod. Sci. 2014, 21, 883–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hasan, Y.M.; Evans, L.C.; Pinkas, G.A.; Dabkowski, E.R.; Stanley, W.C.; Thompson, L.P. Chronic hypoxia impairs cytochrome oxidase activity via oxidative stress in selected fetal Guinea pig organs. Reprod Sci. 2013, 20, 299–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.; Reyes, L.M.; Morton, J.S.; Fung, D.; Schneider, J.; Davidge, S.T. Effect of resveratrol on metabolic and cardiovascular function in male and female adult offspring exposed to prenatal hypoxia and a high-fat diet. J. Physiol. 2016, 594, 1465–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Count in Gene Set | FDR |
|---|---|---|
| Biological Process (GO) | ||
| small molecule metabolic process (GO:0044281) | 102/933 | 1.94 × 10−83 |
| oxidation-reduction process (GO:0055114) | 82/564 | 1.06 × 10−73 |
| generation of precursor metabolites and energy (GO:0006091) | 50/187 | 4.30 ×10−54 |
| Molecular Function (GO) | ||
| catalytic activity (GO:0003824) | 118/2668 | 3.23 × 10−58 |
| oxidoreductase activity (GO:0016491) | 61/458 | 3.20 × 10−51 |
| cofactor binding (GO:0048037) | 46/350 | 1.55 × 10−37 |
| Cellular Component (GO) | ||
| mitochondrion (GO:0005739) | 123/817 | 3.99 × 10−122 |
| mitochondrial part (GO:0044429) | 94/514 | 1.14 × 10−95 |
| mitochondrial matrix (GO:0005759) | 52/179 | 7.22 × 10−59 |
| KEGG Pathway | ||
| metabolic pathways (rno01100) | 109/1240 | 3.34 × 10−82 |
| oxidative phosphorylation (rno00190) | 40/130 | 1.52 × 10−45 |
| carbon metabolism (rno01200) | 37/112 | 4.08 × 10−43 |
| Reactome Pathway | ||
| TCA cycle and respiratory electron transport (RNO-1428517) | 46/123 | 8.62 × 10−56 |
| metabolism (RNO-1430728) | 82/1330 | 2.63 × 10−47 |
| pyruvate metabolism and TCA cycle (RNO-71406) | 22/34 | 2.13 × 10−30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Dasgupta, C.; Huang, L.; Song, R.; Zhang, Z.; Zhang, L. Multi-Omics Integration Reveals Short and Long-Term Effects of Gestational Hypoxia on the Heart Development. Cells 2019, 8, 1608. https://doi.org/10.3390/cells8121608
Gao Y, Dasgupta C, Huang L, Song R, Zhang Z, Zhang L. Multi-Omics Integration Reveals Short and Long-Term Effects of Gestational Hypoxia on the Heart Development. Cells. 2019; 8(12):1608. https://doi.org/10.3390/cells8121608
Chicago/Turabian StyleGao, Yu, Chiranjib Dasgupta, Lei Huang, Rui Song, Ziwei Zhang, and Lubo Zhang. 2019. "Multi-Omics Integration Reveals Short and Long-Term Effects of Gestational Hypoxia on the Heart Development" Cells 8, no. 12: 1608. https://doi.org/10.3390/cells8121608
APA StyleGao, Y., Dasgupta, C., Huang, L., Song, R., Zhang, Z., & Zhang, L. (2019). Multi-Omics Integration Reveals Short and Long-Term Effects of Gestational Hypoxia on the Heart Development. Cells, 8(12), 1608. https://doi.org/10.3390/cells8121608