Implications and Current Limitations of Oogenesis from Female Germline or Oogonial Stem Cells in Adult Mammalian Ovaries


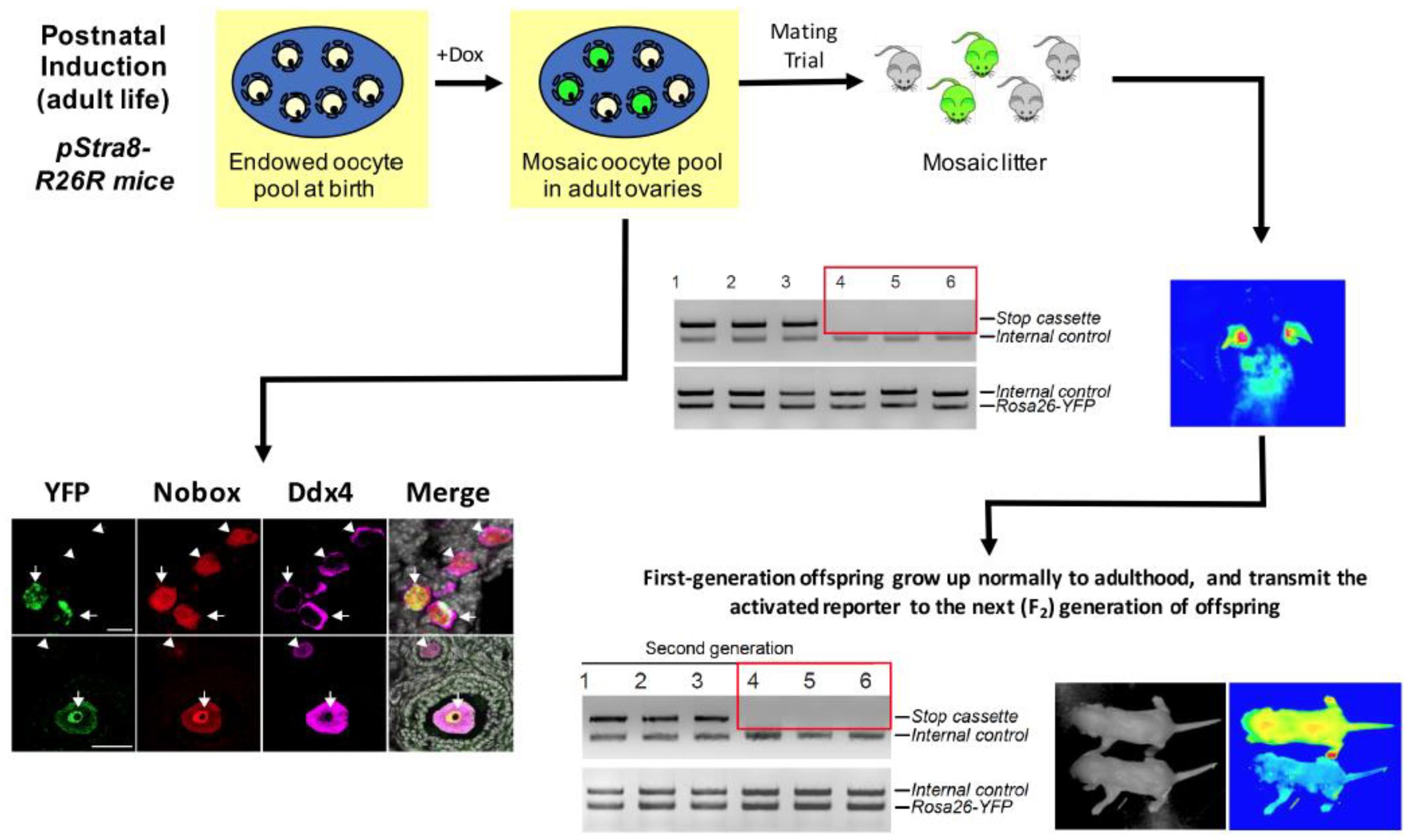{kind=link}
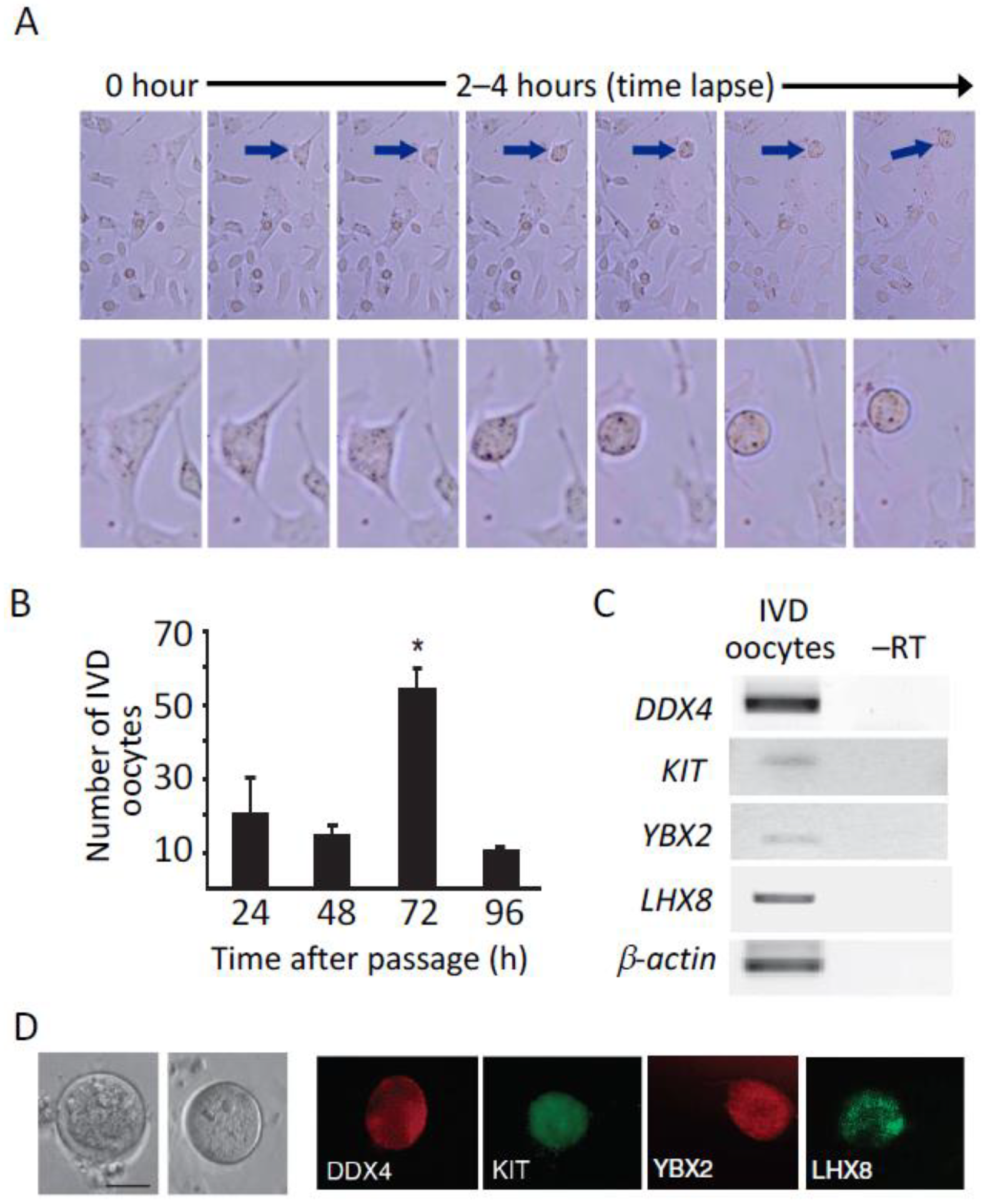{kind=link}
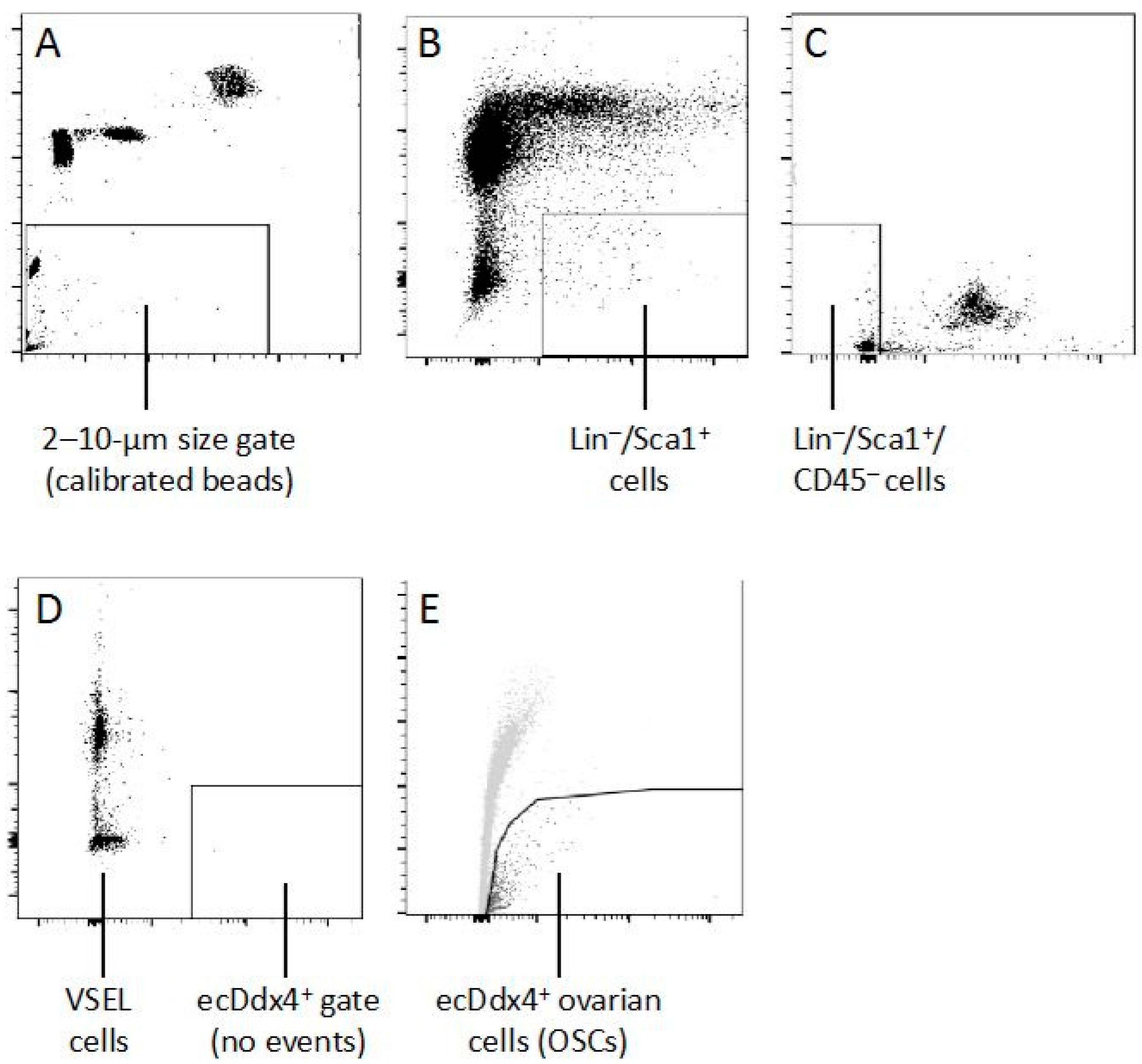{kind=link}
Abstract
:1. Introduction: A Brief History of Postnatal Oogenesis in Mammals
2. Germline Stem Cells in Adult Female Flies—So, Why Not Female Mammals?
3. Discovery and Characterization of Human OSCs
4. ‘Artificial’ Eggs in a Dish from Pluripotent Stem Cells
5. Will Stem Cell-Derived Oocytes be of Future Value to Human Assisted Reproduction?
Funding
Acknowledgments
Conflicts of Interest
References
- Everett, N.B. The present status of the germ-cell problem in vertebrates. Biol. Rev. Camb. Philos. Soc. 1945, 20, 45–55. [Google Scholar] [CrossRef]
- Waldeyer-Hartz, W. Eierstock und Ei; Engelmann: Leipzig, Germany, 1870. [Google Scholar]
- Morris, M.A. Pregnancy following removal of both ovaries and tubes. Boston Med. Surg. J. 1901, 144, 86–87. [Google Scholar] [CrossRef]
- Arai, H. On the postnatal development of the ovary (albino rat), with especial reference to the number of ova. Am. J. Anat. 1920, 27, 405–462. [Google Scholar] [CrossRef]
- Allen, E. Ovogenesis during sexual maturity. Am. J. Anat. 1923, 31, 439–482. [Google Scholar] [CrossRef]
- Davenport, C.B. Regeneration of ovaries in mice. J. Exp. Zool. 1925, 42, 1–12. [Google Scholar] [CrossRef]
- Butcher, E.O. The origin of the definitive ova in the white rat (Mus norvegicus albinus). Anat. Rec. 1927, 37, 13–29. [Google Scholar] [CrossRef]
- Parkes, A.S.; Fielding, U.; Brambell, W.R. Ovarian regeneration in the mouse after complete double ovariectomy. Proc. R. Soc. Lond. Ser. B 1927, 101, 328–354. [Google Scholar] [CrossRef]
- Pallot, O. Apropos de la régénération ovarienne et des modifications périodiques de l’épithelium vaginal chez le rat blanc. Comp. Rend. Soc. Biol. 1928, 99, 1333–1334. [Google Scholar]
- Schwarz, O.H.; Young, C.C., Jr.; Crouse, J.C. Ovogenesis in the adult human ovary. Am. J. Obstet. Gynecol. 1949, 58, 54–64. [Google Scholar] [CrossRef]
- Zuckerman, S. The number of oocytes in the mature ovary. Rec. Prog. Horm. Res. 1951, 6, 63–108. [Google Scholar]
- Franchi, L.L.; Mandl, A.M.; Zuckerman, S. The development of the ovary and the process of oogenesis. In The Ovary; Zuckerman, S., Ed.; Academic Press: New York, NY, USA, 1962; pp. 1–88. [Google Scholar]
- Zuckerman, S. Beyond the Ivory Tower. The Frontiers of Public and Private Science; Taplinger: New York, NY, USA, 1971; pp. 22–34. [Google Scholar]
- Pearl, R.; Schoppe, W.F. Studies on the physiology of reproduction in the domestic fowl. J. Exp. Zool. 1921, 34, 101–118. [Google Scholar] [CrossRef]
- Pansky, B.; Mossman, H.W. The regenerative capacity of the rabbit ovary. Anat. Rec. 1953, 116, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Vermande-Van Eck, G.J. Neo-ovogenesis in the adult monkey. Consequences of atresia of ovocytes. Anat. Rec. 1956, 125, 207–224. [Google Scholar] [CrossRef] [PubMed]
- Artem’eva, N.S. Regenerative capacity of the rat ovary after compensatory hypertrophy. Bull. Exp. Biol. Med. 1961, 51, 76–81. [Google Scholar] [CrossRef]
- King, R.C.; Rubinson, A.C.; Smith, R.F. Oogenesis in adult Drosophila melanogaster. Growth 1956, 20, 121–157. [Google Scholar] [PubMed]
- Lin, H.; Spradling, A.C. Germline stem cell division and egg chamber development in transplanted Drosophila germaria. Dev. Biol. 1993, 159, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.M.; Schedl, P. Establishment of stem cell identity in the Drosophila germline. Dev. Dyn. 1997, 210, 371–382. [Google Scholar] [CrossRef]
- Deng, W.; Lin, H. Asymmetric germ cell division and oocyte determination during Drosophila oogenesis. Int. Rev. Cytol. 2001, 203, 93–138. [Google Scholar] [PubMed]
- Schulze, C. Morphological characteristics of the spermatogonial stem cells in man. Cell Tissue Res. 1979, 198, 191–199. [Google Scholar] [CrossRef]
- Lin, H. The stem-cell niche theory: Lessons from flies. Nat. Genet. 2002, 3, 931–940. [Google Scholar] [CrossRef]
- Brinster, R.L. Male germline stem cells: From mice to men. Science 2007, 316, 404–405. [Google Scholar] [CrossRef] [PubMed]
- Matunis, E.L.; Stine, R.R.; de Cuevas, M. Recent advances in Drosophila male germline stem cell biology. Spermatogenesis 2012, 2, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, L.J.; de Cuevas, M.; Matunis, E. Genetics of gonadal stem cell renewal. Annu. Rev. Cell Dev. Biol. 2015, 31, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Kirilly, D.; Xie, T. The Drosophila ovary: An active stem cell community. Cell Res. 2007, 17, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Xuan, Y.; Li, X.; Xi, R. Age-related changes of germline stem cell activity, niche signaling activity and egg production in Drosophila. Aging Cell 2008, 7, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Tworzydlo, W.; Kloc, M.; Bilinski, S.M. Female germline stem cell niches of earwigs are structurally simple and different from those of Drosophila melanogaster. J. Morphol. 2010, 271, 634–640. [Google Scholar] [PubMed]
- Underwood, J.L.; Hestand, R.S., III; Thompson, B.Z. Gonad regeneration in grass carp following bilateral gonadectomy. Prog. Fish-Cult. 1986, 48, 54–56. [Google Scholar] [CrossRef]
- Kersten, C.A.; Krisfalusi, M.; Parsons, J.E.; Cloud, J.G. Gonadal regeneration in masculinized female or steroid-treated rainbow trout (Oncorhynchus mykiss). J. Exp. Zool. 2001, 290, 396–401. [Google Scholar] [CrossRef]
- Draper, B.W.; McCallum, C.M.; Moens, C.B. nanos1 is required to maintain oocyte production in adult zebrafish. Dev. Biol. 2007, 305, 589–598. [Google Scholar] [CrossRef]
- Nakamura, S.; Kobayashi, K.; Nishimura, T.; Higashijima, S.; Tanaka, M. Identification of germline stem cells in the ovary of the teleost medaka. Science 2010, 328, 1561–1563. [Google Scholar] [CrossRef]
- Nakamura, S.; Kobayashi, K.; Nishimura, T.; Tanaka, M. Ovarian germline stem cells in the teleost fish, medaka (Oryzias latipes). Int. J. Biol. Sci. 2011, 7, 403–409. [Google Scholar] [CrossRef] [PubMed]
- White, Y.A.R.; Woods, D.C.; Wood, A.W. A transgenic zebrafish model of targeted oocyte ablation and de novo oogenesis. Dev. Dyn. 2011, 240, 1929–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, D.C.; Tilly, J.L. An evolutionary perspective on adult female germline stem cell function from flies to humans. Semin. Reprod. Med. 2013, 31, 24–32. [Google Scholar] [PubMed]
- Johnson, J.; Canning, J.; Kaneko, T.; Pru, J.K.; Tilly, J.L. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 2004, 428, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F. Strong science challenges conventional wisdom: New perspectives on ovarian biology. Reprod. Biol. Endocrinol. 2004, 2, 28. [Google Scholar] [CrossRef] [PubMed]
- Byskov, A.G.; Faddy, M.J.; Lemmen, J.G.; Andersen, C.Y. Eggs forever? Differentiation 2005, 73, 438–446. [Google Scholar] [CrossRef]
- Skaznik-Wikiel, M.; Tilly, J.C.; Lee, H.-J.; Niikura, Y.; Kaneko-Tarui, T.; Johnson, J.; Tilly, J.L. Serious doubts over “Eggs Forever?”. Differentiation 2007, 75, 93–99. [Google Scholar] [CrossRef]
- Johnson, J.; Bagley, J.; Skaznik-Wikiel, M.; Lee, H.-J.; Adams, G.B.; Niikura, Y.; Tschudy, K.S.; Tilly, J.C.; Cortes, M.L.; Forkert, R.; et al. Oocyte generation in adult mammalian ovaries by putative germ cells derived from bone marrow and peripheral blood. Cell 2005, 122, 303–315. [Google Scholar] [CrossRef]
- Telfer, E.E.; Gosden, R.G.; Byskov, A.G.; Spears, N.; Albertini, D.; Andersen, C.Y.; Anderson, R.; Braw-Tal, R.; Clarke, H.; Gougeon, A.; et al. On regenerating the ovary and generating controversy. Cell 2005, 122, 821–822. [Google Scholar] [CrossRef]
- Johnson, J.; Skaznik-Wikiel, M.; Lee, H.-J.; Niikura, Y.; Tilly, J.C.; Tilly, J.L. Setting the record straight on data supporting postnatal oogenesis in female mammals. Cell Cycle 2005, 4, 1471–1477. [Google Scholar]
- Eggan, K.; Jurga, S.; Gosden, R.; Min, I.M.; Wagers, A. Ovulated oocytes in adult mice derive from non-circulating germ cells. Nature 2006, 441, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Selesniemi, K.; Niikura, Y.; Niikura, T.; Klein, R.; Dombkowski, D.M.; Tilly, J.L. Bone marrow transplantation generates immature oocytes and rescues long-term fertility in a preclinical mouse model of chemotherapy-induced premature ovarian failure. J. Clin. Oncol. 2007, 25, 3198–3204. [Google Scholar] [CrossRef]
- Selesniemi, K.; Lee, H.-J.; Niikura, T.; Tilly, J.L. Young adult donor bone marrow infusions into female mice postpone age-related reproductive failure and improve offspring survival. Aging 2008, 1, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Niikura, Y.; Niikura, T.; Wang, N.; Satirapod, C.; Tilly, J.L. Systemic signals in aged males exert potent rejuvenating effects on the ovarian follicle reserve in mammalian females. Aging 2010, 2, 999–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, J.B.; Duckett, R.; Myers, M.; Britt, K.L.; Mladenovska, T.; Findlay, J.K. Quantification of healthy follicles in the neonatal and adult mouse ovary: Evidence for maintenance of primordial follicle supply. Reproduction 2006, 132, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Niikura, Y.; Niikura, T.; Tilly, J.L. Aged mouse ovaries possess rare premeiotic germ cells that can generate oocytes following transplantation into a young host environment. Aging 2009, 1, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wu, C.; Lyu, Q.; Yang, D.; Albertini, D.F.; Keefe, D.L.; Liu, L. Germline stem cells and neo-oogenesis in the adult human ovary. Dev. Biol. 2007, 306, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Tilly, J.L.; Johnson, J. Recent arguments against germ cell renewal in the adult human ovary. Is an absence of marker gene expression really acceptable evidence of an absence of oogenesis? Cell Cycle 2007, 6, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Tilly, J.L.; Niikura, Y.; Rueda, B.R. The current status of evidence for and against postnatal oogenesis in mammals: A case of ovarian optimism versus pessimism? Biol. Reprod. 2009, 80, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Yuan, Z.; Yang, Z.; Luo, H.; Sun, K.; Zhou, L.; Xiang, J.; Shi, L.; Yu, Q.; Zhang, Y.; et al. Production of offspring from a germline stem cell line derived from neonatal ovaries. Nat. Cell Biol. 2009, 11, 631–636. [Google Scholar] [CrossRef]
- Tilly, J.L.; Telfer, E.E. Purification of germline stem cells from adult mammalian ovaries: A step closer towards control of the female biological clock? Mol. Hum. Reprod. 2009, 15, 393–398. [Google Scholar] [CrossRef] [PubMed]
- De Felici, M. Germ stem cells in the mammalian adult ovary: Considerations by a fan of primordial germ cells. Mol. Hum. Reprod. 2010, 16, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; Telfer, E.E.; Tilly, J.L. Oocyte family trees: Old branches or new stems? PLoS Genet. 2012, 8, e1002848. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, A.; Notarianni, E. There is no neo-oogenesis in the adult mammalian ovary. J. Turk. Ger. Gynecol. Assoc. 2011, 12, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Notarianni, E. Reinterpretation of evidence advanced for neo-oogenesis in mammals, in terms of a finite oocyte reserve. J. Ovarian Res. 2011, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oatley, J.M.; Hunt, P.A. Of mice and (wo)men: Purified oogonial stem cells from mouse and human ovaries. Biol. Reprod. 2012, 86, 196. [Google Scholar] [CrossRef]
- Woods, D.C.; White, Y.A.R.; Tilly, J.L. Purification of oogonial stem cells from adult mouse and human ovaries: An assessment of the literature and a view towards the future. Reprod. Sci. 2013, 20, 7–15. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, W.; Shen, Y.; Adhikari, D.; Ueno, H.; Liu, K. Experimental evidence showing that no mitotically active female germline progenitors exist in postnatal mouse ovaries. Proc. Natl. Acad. Sci. USA 2012, 109, 12580–12585. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Zhang, D.; Wang, L.; Liu, M.; Mao, J.; Yin, Y.; Ye, X.; Liu, N.; Han, J.; Gao, Y.; et al. No evidence for neo-oogenesis may link to ovarian senescence in adult monkey. Stem Cells 2013, 31, 2536–2550. [Google Scholar] [CrossRef]
- Lei, L.; Spradling, A.C. Female mice lack adult germ-line stem cells but sustain oogenesis using stable primordial follicles. Proc. Natl. Acad. Sci. USA 2013, 110, 8585–8590. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, S.F.; Vahidi, N.A.; Park, S.; Weitzel, R.P.; Tisdale, J.; Rueda, B.R.; Wolff, E.F. Characterization of extracellular DDX4- or Ddx4-positive ovarian cells. Nat. Med. 2015, 21, 1114–1116. [Google Scholar] [CrossRef]
- Zhang, H.; Panula, S.; Petropoulos, S.; Edsgärd, D.; Busayavalasa, K.; Liu, L.; Li, X.; Risal, S.; Shen, Y.; Shao, J.; et al. Adult human and mouse ovaries lack DDX4-expressing functional oogonial stem cells. Nat. Med. 2015, 21, 1116–1118. [Google Scholar] [CrossRef] [Green Version]
- Park, E.S.; Tilly, J.L. Use of DEAD-box polypeptide 4 (Ddx4) gene promoter-driven fluorescent reporter mice to identify mitotically active germ cells in postnatal mouse ovaries. Mol. Hum. Reprod. 2015, 21, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wu, J. Production of offspring from a germline stem cell line derived from prepubertal ovaries of germline reporter mice. Mol. Hum. Reprod. 2016, 22, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, D.C.; Tilly, J.L. Isolation, characterization and propagation of mitotically active germ cells from adult mouse and human ovaries. Nat. Protoc. 2013, 8, 966–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, D.C.; Tilly, J.L. Reply to adult human and mouse ovaries lack DDX4-expressing functional oogonial stem cells. Nat. Med. 2015, 21, 1118–1121. [Google Scholar] [CrossRef]
- Pacchiarotti, J.; Maki, C.; Ramos, T.; Marh, J.; Howerton, K.; Wong, J.; Pham, J.; Anorve, S.; Chow, Y.C.; Izadyar, F. Differentiation potential of germ line stem cells derived from the postnatal mouse ovary. Differentiation 2010, 79, 159–170. [Google Scholar] [CrossRef]
- Wang, N.; Tilly, J.L. Epigenetic status determines germ cell meiotic commitment in embryonic and postnatal mammalian gonads. Cell Cycle 2010, 9, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, Z.; Yang, Y.; Wang, S.; Shi, L.; Xie, W.; Sun, K.; Zou, K.; Wang, L.; Xiong, J.; et al. Production of transgenic mice by random recombination of targeted genes in female germline stem cells. J. Mol. Cell Biol. 2011, 3, 132–141. [Google Scholar] [CrossRef]
- White, Y.A.R.; Woods, D.C.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J.L. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Imudia, A.N.; Wang, N.; Tanaka, Y.; White, Y.A.; Woods, D.C.; Tilly, J.L. Comparative gene expression profiling of adult mouse ovary-derived oogonial stem cells supports a distinct cellular identity. Fertil. Steril. 2013, 100, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Park, E.S.; Woods, D.C.; Tilly, J.L. Bone morphogenetic protein 4 (BMP4) promotes mammalian oogonial stem cell differentiation via SMAD1/5/8 signaling. Fertil. Steril. 2013, 100, 1468–1475. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, M.; Bi, H.; Chen, X.; He, L.; Li, X.; Wu, J. Conversion of female germline stem cells from neonatal and prepubertal mice into pluripotent stem cells. J. Mol. Cell Biol. 2014, 6, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Wang, H.; Wu, J. Similar morphological and molecular signatures shared by female and male germline stem cells. Sci. Rep. 2014, 4, 5580. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, L.; Kang, J.X.; Xie, W.; Li, X.; Wu, C.; Xu, B.; Wu, J. Production of fat-1 transgenic rats using a post-natal female germline stem cell line. Mol. Hum. Reprod. 2014, 20, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Khosravi-Farsani, S.; Amidi, F.; Roudkenar, M.H.; Sobhani, A. Isolation and enrichment of mouse female germ line stem cells. Cell J. 2015, 16, 406–415. [Google Scholar] [PubMed]
- Xiong, J.; Lu, Z.; Wu, M.; Zhang, J.; Cheng, J.; Luo, A.; Shen, W.; Fang, L.; Zhou, S.; Wang, S. Intraovarian transplantation of female germline stem cells rescues ovarian function in chemotherapy injured ovaries. PLoS ONE 2015, 10, e0139824. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Li, C.-H.; Wang, X.-Y.; He, D.-J.; Zheng, P. Germ stem cells are active in postnatal mouse ovary under physiological conditions. Mol. Hum. Reprod. 2016, 22, 316–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Wu, M.; Zhang, J.; Xiong, J.; Cheng, J.; Shen, W.; Luo, A.; Fang, L.; Wang, S. Improvement in isolation and identification of mouse oogonial stem cells. Stem Cells Int. 2016, 2016, 2749461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Wu, J.; Wang, J.; Shen, T.; Li, H.; Lu, J.; Gu, Y.; Kang, Y.; Wong, C.H.; Ngan, C.Y.; et al. Integrative epigenomic analysis reveals unique epigenetic signatures involved in unipotency of mouse female germline stem cells. Genome Biol. 2016, 17, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Satirapod, C.; Ohguchi, Y.; Park, E.-S.; Woods, D.C.; Tilly, J.L. Genetic studies in mice directly link oocytes produced during adulthood to ovarian function and natural fertility. Sci. Rep. 2017, 7, 10011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Xu, B.; Li, X.; Ma, W.; Zhang, P.; Wu, J. Tracing and characterizing the development of transplanted female germline stem cells in vivo. Mol. Ther. 2017, 25, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Zheng, T.; Hu, C.; Pan, Z.; Huang, J.; Li, J.; Li, W.; Zheng, Y. The Hippo signaling pathway regulates ovarian function via the proliferation of ovarian germline stem cells. Cell. Physiol. Biochem. 2017, 41, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Wu, J.; Yang, W.; Xia, C.; Shi, X.; Li, H.; Sun, J.; Shao, Z.; Wu, J.; Zhao, X. STAT3 is required for proliferation and exerts a cell type-specific binding preference in mouse female germline stem cells. Mol. Omics 2018, 14, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gong, X.; Tian, G.G.; Hou, C.; Zhu, X.; Pei, X.; Wang, Y.; Wu, J. Long noncoding RNA growth arrest-specific 5 promotes proliferation and survival of female germline stem cells in vitro. Gene 2018, 653, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Xiong, J.; Ma, L.; Lu, Z.; Qin, X.; Luo, A.; Zhang, J.; Xie, H.; Shen, W.; Wang, S. Enrichment of female germline stem cells from mouse ovaries using the differential adhesion method. Cell. Physiol. Biochem. 2018, 46, 2114–2126. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yao, X.; Tang, F.; Wei, Y.; Hua, J.; Peng, S. Characterization of female germline stem cells from adult mouse ovaries and the role of rapamycin on them. Cytotechnol. 2018, 70, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Yang, Y.; Xia, Q.; Song, H.F.; Wei, R.; Wang, J.J.; Zou, K. Cadherin 22 participates in the self-renewal of mouse female germline stem cells via interaction with JAK2 and ß-catenin. Cell. Mol. Life Sci. 2018, 75, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Tian, G.G.; Yu, B.; Yang, Y.; Wu, J. Effects of bisphenol A on ovarian follicular development and female germline stem cells. Arch. Toxicol. 2018, 92, 1581–1591. [Google Scholar] [CrossRef]
- Dunlop, C.E.; Bayne, R.A.; McLaughlin, M.; Telfer, E.E.; Anderson, R.A. Isolation, purification, and culture of oogonial stem cells from adult human and bovine ovarian cortex. Lancet 2014, 383, S45. [Google Scholar] [CrossRef]
- de Souza, G.B.; Costa, J.; da Cunha, E.V.; Passos, J.; Ribeiro, R.P.; Saraiva, M.; van den Hurk, R.; Silva, J. Bovine ovarian stem cells differentiate into germ cells and oocyte-like structures after culture in vitro. Reprod. Domest. Anim. 2017, 52, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.-S.; Johnson, J.; White, Y.; St. John, J.C. The molecular characterization of porcine egg precursor cells. Oncotarget 2017, 8, 63484–63505. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Wang, J.; Li, X.; Wang, H.; Liu, G.; Xu, B.; Mei, X.; Hua, X.; Wu, J. Characteristics of female germline stem cells from porcine ovaries at sexual maturity. Cell Transplant. 2018, 27, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Liu, G.; Xu, B.; Wu, C.; Hui, N.; Ni, X.; Wang, J.; Du, M.; Teng, X.; Wu, J. Human GV oocytes generated by mitotically active germ cells obtained from follicular aspirates. Sci. Rep. 2016, 6, 28218. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, Y.L.; McLaughlin, M.; Waterfall, M.; Dunlop, C.E.; Skehel, P.A.; Anderson, R.A.; Telfer, E.E. Initial characterisation of adult human ovarian cell populations isolated by DDX4 expression and aldehyde dehydrogenase activity. Sci. Rep. 2018, 8, 6953. [Google Scholar] [CrossRef] [PubMed]
- Silvestris, E.; Cafforio, P.; D’Oronzo, S.; Felici, C.; Silvestris, F.; Loverro, G. In vitro differentiation of human oocyte-like cells from oogonial stem cells: Single-cell isolation and molecular characterization. Hum. Reprod. 2018, 33, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Silvestris, E.; D’Oronzo, S.; Cafforio, P.; D’Amato, G.; Loverno, G. Perspective in infertility: The ovarian stem cells. J. Ovarian Res. 2015, 8, 55. [Google Scholar] [CrossRef]
- Bothun, A.; Gao, Y.; Takai, Y.; Ishihara, O.; Seki, H.; Karger, B.; Tilly, J.L.; Woods, D.C. Quantitative proteomic profiling of the human ovary from early to mid-gestation reveals protein expression dynamics of oogenesis and folliculogenesis. Stem Cells Dev. 2018, 27, 723–735. [Google Scholar] [CrossRef]
- Fakih, M.H.; El Shmoury, M.; Szeptycki, J.; dela Cruz, D.B.; Lux, C.; Verjee, S.; Burgess, C.M.; Cohn, G.M.; Casper, R.F. The AUGMENTSM treatment: Physician reported outcomes of the initial global patient experience. JFIV Reprod. Med. Genet. 2015, 3, 154. [Google Scholar] [CrossRef]
- Oktay, K.; Baltaci, V.; Sonmezer, M.; Turan, V.; Unsal, E.; Baltaci, A.; Aktuna, S.; Moy, F. Oogonial precursor cell derived autologous mitochondria injection (AMI) to improve outcomes in women with multiple IVF failures due to low oocyte quality: A clinical translation. Reprod. Sci. 2015, 22, 1612–1617. [Google Scholar] [CrossRef]
- Woods, D.C.; Tilly, J.L. Autologous germline mitochondrial energy transfer (AUGMENT) in human assisted reproduction. Semin. Reprod. Med. 2015, 33, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Grieve, K.M.; McLaughlin, M.; Dunlop, C.E.; Telfer, E.E.; Anderson, R.A. The controversial existence and functional potential of oogonial stem cells. Maturitas 2015, 83, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; Tilly, J.L. The next (re)generation of human ovarian biology and female fertility: Is current science tomorrows practice? Fertil. Steril. 2012, 98, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; Tilly, J.L. Germline stem cells in adult mammalian ovaries. In Ten Critical Topics in Reproductive Medicine; Sanders, S., Ed.; Science/AAAS: Washington, DC, USA, 2013; pp. 10–12. [Google Scholar]
- Truman, A.M.; Tilly, J.L.; Woods, D.C. Ovarian regeneration: The potential for stem cell contribution in the postnatal ovary to sustained endocrine function. Mol. Cell. Endocrinol. 2017, 445, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, Y.; Komiya, T.; Kawabata, H.; Sato, M.; Fujimoto, H.; Furusawa, M.; Noce, T. Isolation of a DEAD-family protein gene that encodes a murine homolog of Drosophila vasa and its specific expression in germ cell lineage. Proc. Natl. Acad. Sci. USA 1994, 91, 12258–12262. [Google Scholar] [CrossRef] [PubMed]
- Toyooka, Y.; Tsunekawa, N.; Takahashi, Y.; Matsui, Y.; Satoh, M.; Noce, T. Expression and intracellular localization of mouse Vasa-homologue protein during germ cell development. Mech. Dev. 2000, 93, 139–149. [Google Scholar] [CrossRef]
- Castrillon, D.H.; Quade, B.J.; Wang, T.Y.; Quigley, C.; Crum, C.P. The human VASA gene is specifically expressed in the germ lineage. Proc. Natl. Acad. Sci. USA 2000, 97, 9585–9590. [Google Scholar] [CrossRef] [PubMed]
- Navaroli, D.M.; Tilly, J.L.; Woods, D.C. Isolation of mammalian oogonial stem cells by antibody-based fluorescence-activated cell sorting. Meth. Mol. Biol. 2016, 1457, 253–268. [Google Scholar]
- Crisan, M.; Dzierak, E. The many faces of hematopoietic stem cell heterogeneity. Development 2016, 143, 4571–4581. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, J.A.; Takai, Y.; Ishihara, O.; Seki, H.; Woods, D.C.; Tilly, J.L. Extracellular matrix signaling activates differentiation of adult ovary-derived oogonial stem cells in a species-specific manner. Fertil. Steril. 2019. in press. [Google Scholar] [CrossRef]
- Gosden, R.G.; Clarke, H.; Miller, D. Female gametogenesis. In Reproductive Medicine. Molecular, Cellular and Genetic Fundamentals; Fauser, B.C.J.M., Ed.; Parthenon Publishing: New York, NY, USA, 2003; pp. 365–380. [Google Scholar]
- West, F.D.; Mumaw, J.L.; Gallegos-Cardenas, A.; Young, A.; Stice, S.L. Human haploid cells differentiated from meiotic competent clonal germ cell lines that originated from embryonic stem cells. Stem Cells Dev. 2011, 20, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Ogushi, S.; Kurimoto, K.; Shimamoto, S.; Ohta, H.; Saitou, M. Offspring from oocytes derived from in vitro primordial germ cell-like cells in mice. Science 2012, 338, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Hikabe, O.; Hamazaki, N.; Nagamatsu, G.; Obata, Y.; Hirao, Y.; Hamada, N.; Shimamoto, S.; Imamura, T.; Nakashima, K.; Saitou, M.; et al. Reconstitution in vitro of the entire cycle of the mouse female germ line. Nature 2016, 539, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Panula, S.; Medrano, J.V.; Kee, K.; Bergström, R.; Nguyen, H.N.; Byers, B.; Wilson, K.D.; Wu, J.C.; Simon, C.; Hovatta, O.; et al. Human germ cell differentiation from fetal- and adult-derived induced pluripotent stem cells. Hum. Mol. Genet. 2011, 20, 752–762. [Google Scholar] [PubMed]
- Yamashiro, C.; Sasaki, K.; Yabuta, Y.; Kojima, Y.; Nakamura, T.; Okamoto, I.; Yokobayashi, S.; Murase, Y.; Ishikura, Y.; Shirane, K.; et al. Generation of human oogonia from induced pluripotent stem cells. Science 2018, 362, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.; Xiong, J.; Ye, M.; Qin, X.; Li, L.; Cheng, S.; Luo, M.; Peng, J.; Dong, J.; Tang, F.; et al. In vitro differentiation of human embryonic stem cells into ovarian follicle-like cells. Nat. Commun. 2017, 8, 15680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin, M.; Albertini, D.F.; Wallace, W.H.B.; Anderson, R.A.; Telfer, E.E. Metaphase II oocytes from human unilaminar follicles grown in a multi-step culture system. Mol. Hum. Reprod. 2018, 24, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Smitz, J.E.; Gilchrist, R.B. Are human oocytes from stem cells next? Nat. Biotechnol. 2016, 34, 1247–1248. [Google Scholar] [CrossRef]
- Cohen, I.G.; Daley, G.Q.; Adashi, E.Y. Disruptive reproductive technologies. Sci. Transl. Med. 2017, 9, eaag2959. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.B.; Larsson, N.-G. Keeping mtDNA in shape between generations. PLoS Genet. 2014, 10, e1004670. [Google Scholar] [CrossRef]
- Stewart, J.B.; Chinnery, P.F. The dynamics of mitochondrial DNA heteroplasmy: Implications for human health and disease. Nat. Rev. Genet. 2015, 16, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; Khrapko, K.; Tilly, J.L. Influence of maternal aging on mitochondrial heterogeneity, inheritance, and function in oocytes and preimplantation embryos. Genes 2018, 9, 265. [Google Scholar] [CrossRef] [PubMed]
- Bhartiya, D.; Patel, H.; Parte, S. Improved understanding of very small embryonic-like stem cells in adult mammalian ovary. Hum. Reprod. 2018, 33, 978–979. [Google Scholar] [CrossRef] [PubMed]
- Kucia, M.; Reca, R.; Campbell, F.R.; Zuba-Surma, E.; Majka, M.; Ratajczak, J.; Ratajczak, M.Z. A population of very small embryonic-like (VSEL) CXCR4+SSEA4+Oct-4+ stem cells identified in adult bone marrow. Leukemia 2006, 20, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Heider, A.; Danova-Alt, R.; Egger, D.; Cross, M.; Alt, R. Murine and human very small embryonic-like cells: A perspective. Cytometry A 2013, 83, 72–75. [Google Scholar] [CrossRef]
- Miyanishi, M.; Mori, Y.; Seita, J.; Chen, J.Y.; Karten, S.; Chan, C.K.F.; Nakauchi, H.; Weissman, I.L. Do pluripotent stem cells exist in mice as very small embryonic-like cells? Stem Cell Rep. 2013, 1, 198–208. [Google Scholar] [CrossRef]
- Havens, A.M.; Sun, H.; Shiozawa, Y.; Jung, Y.; Wang, J.; Mishra, A.; Jiang, Y.; O’Neill, D.W.; Krebsbach, P.H.; Rodgerson, D.O.; et al. Human and murine very small embryonic-like cells represent multipotent tissue progenitors, in vitro and in vivo. Stem Cells Dev. 2014, 23, 689–701. [Google Scholar] [CrossRef]
- Kim, Y.; Jeong, J.; Kang, H.; Lim, J.; Heo, J.; Ratajczak, J.; Ratajczak, M.Z.; Shin, D.M. The molecular nature of very small embryonic-like stem cells in adult tissues. Int. J. Stem Cells 2014, 7, 55–62. [Google Scholar] [CrossRef]
- Virant-Klun, I.; Zech, N.; Rozman, P.; Vogler, A.; Cvjeticanin, B.; Klemenc, P.; Malicev, E.; Meden-Vrtovec, H. Putative stem cells with an embryonic character isolated from the ovarian surface epithelium of women with no naturally present follicles or oocytes. Differentiation 2008, 76, 843–856. [Google Scholar] [CrossRef]
- Gong, S.P.; Lee, S.T.; Lee, E.J.; Kim, D.Y.; Lee, G.; Chi, S.G.; Ryu, B.K.; Lee, C.H.; Yum, K.E.; Lee, H.J.; et al. Embryonic stem cell-like cells established by culture of adult ovarian cells in mice. Fertil. Steril. 2010, 93, 2594–2601. [Google Scholar] [CrossRef]
- Parte, S.; Bhartiya, D.; Telang, J.; Daithankar, V.; Salvi, V.; Zaveri, K.; Hinduja, I. Detection, characterization, and spontaneous differentiation in vitro of very small embryonic-like putative stem cells in adult mammalian ovary. Stem Cells Dev. 2011, 20, 1451–1464. [Google Scholar] [CrossRef] [PubMed]
- Bhartiya, D.; Patel, H. Ovarian stem cells—Resolving controversies. J. Assist. Reprod. Genet. 2018, 35, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Bhartiya, D.; Parte, S.; Gunjal, P.; Yedurkar, S.; Bhatt, M. Follicle-stimulating hormone modulates ovarian stem cells through alternatively spliced receptor variant FSH-R3. J. Ovarian Res. 2013, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Bhartiya, D.; Unni, S.; Parte, S.; Anand, S. Very small embryonic-like cells: Implications in reproductive biology. Biomed. Res. Int. 2013, 2013, 682326. [Google Scholar] [CrossRef] [PubMed]
- Brinster, R.L.; Zimmermann, J.W. Spermatogenesis following male germ-cell transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11298–11302. [Google Scholar] [CrossRef] [PubMed]
- Brinster, R.L.; Avarbock, M.R. Germline transmission of donor haplotype following spermatogonial transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11303–11307. [Google Scholar] [CrossRef] [PubMed]
- Kanatsu-Shinohara, M.; Morimoto, H.; Shinohara, T. Fertility of male germline stem cells following spermatogonial transplantation in infertile mouse models. Biol. Reprod. 2016, 94, 112. [Google Scholar] [CrossRef] [PubMed]
- Tilly, J.L.; Woods, D.C. Compositions and Methods for Autologous Germline Mitochondrial Energy Transfer. U.S. Patent 8,642,329, 4 February 2014. [Google Scholar]
- Tilly, J.L.; Woods, D.C. Compositions and Methods for Autologous Germline Mitochondrial Energy Transfer. U.S. Patent 8,647,869, 11 February 2014. [Google Scholar]
- Park, S.; Yu, J.; Marquis, K.; DeCherney, A.H.; Wolff, E.F. Egg from ovarian-derived stem cells develops into embryo after intracytoplasmic sperm injection. Reprod. Sci. 2016, 23, 264A. [Google Scholar]
- Telfer, E.E.; McLaughlin, M.; Ding, C.; Thong, K.J. A two-step serum-free culture system supports development of human oocytes from primordial follicles in the presence of activin. Hum. Reprod. 2008, 23, 1151–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin, M.; Telfer, E.E. Oocyte development in bovine primordial follicles is promoted by activin and FSH within a two-step serum-free culture system. Reproduction 2010, 139, 971–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Fazleabas, A.; Shikanov, A.; Jackson, E.; Barrett, S.L.; Hirshfield-Cytron, J.; Kiesewetter, S.E.; Shea, L.D.; Woodruff, T.K. In vitro oocyte maturation and preantral follicle culture from the luteal-phase baboon ovary produce mature oocytes. Biol. Reprod. 2011, 84, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Telfer, E.E.; McLaughlin, M. Strategies to support human oocyte development in vitro. Int. J. Dev. Biol. 2012, 56, 901–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trounson, A.; Wood, C.; Kausche, A. In vitro maturation and the fertilization and developmental competence of oocytes recovered from untreated polycystic ovarian patients. Fertil. Steril. 1994, 62, 353–362. [Google Scholar] [CrossRef]
- Cha, K.Y.; Koo, J.J.; Ko, J.J.; Choi, D.H.; Han, S.Y.; Yoon, T.K. Pregnancy after in vitro fertilization of human follicular oocytes collected from nonstimulated cycles, their culture in vitro and their transfer in a donor oocyte program. Fertil. Steril. 1991, 55, 109–113. [Google Scholar] [CrossRef]
- Cha, K.Y.; Chian, R.C. Maturation in vitro of immature human oocytes for clinical use. Hum. Reprod. Update 1998, 4, 103–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.K.; Lee, S.C.; Kim, K.J.; Han, C.H.; Kim, J.H. In-vitro maturation, fertilization, and development of human germinal vesicle oocytes collected from stimulated cycles. Fertil. Steril. 2000, 74, 1153–1158. [Google Scholar] [CrossRef]
- Son, W.Y.; Chung, J.T.; Demirtas, E.; Holzer, H.; Sylvestre, C.; Buckett, W.; Chian, R.C.; Tan, S.L. Comparison of in-vitro maturation cycles with and without in-vivo matured oocytes retrieved. Reprod. Biomed. Online 2008, 17, 59–67. [Google Scholar] [CrossRef]
- Oktay, K.; Buyuk, E.; Rodriguez-Wallberg, K.A.; Sahin, G. In vitro maturation improves oocyte or embryo cryopreservation outcome in breast cancer patients undergoing ovarian stimulation for fertility preservation. Reprod. Biomed. Online 2010, 20, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Fadini, R.; Dal Canto, M.; Mignini Renzini, M.; Milani, R.; Fruscio, R.; Cantù, M.G.; Brambillasca, F.; Coticchio, G. Embryo transfer following in vitro maturation and cryopreservation of oocytes recovered from antral follicles during conservative surgery for ovarian cancer. J. Assist. Reprod. Genet. 2012, 29, 779–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, E.M.; Song, H.S.; Lee, D.R.; Lee, W.S.; Yoon, T.K. In vitro maturation of human oocytes: Its role in infertility treatment and new possibilities. Clin. Exp. Reprod. Med. 2014, 41, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; White, Y.A.R.; Niikura, Y.; Kiatpongsan, S.; Lee, H.J.; Tilly, J.L. Embryonic stem cell derived granulosa cells participate in follicle formation in vitro and in vivo. Reprod. Sci. 2013, 20, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Lipskind, S.; Lindsey, J.S.; Gerami-Naini, B. An embryonic and induced pluripotent stem cell model for ovarian granulosa cell development and steroidogenesis. Reprod. Sci. 2018, 25, 712–726. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, J.J.; Woods, D.C.; Tilly, J.L. Implications and Current Limitations of Oogenesis from Female Germline or Oogonial Stem Cells in Adult Mammalian Ovaries. Cells 2019, 8, 93. https://doi.org/10.3390/cells8020093
Martin JJ, Woods DC, Tilly JL. Implications and Current Limitations of Oogenesis from Female Germline or Oogonial Stem Cells in Adult Mammalian Ovaries. Cells. 2019; 8(2):93. https://doi.org/10.3390/cells8020093
Chicago/Turabian StyleMartin, Jessica J., Dori C. Woods, and Jonathan L. Tilly. 2019. "Implications and Current Limitations of Oogenesis from Female Germline or Oogonial Stem Cells in Adult Mammalian Ovaries" Cells 8, no. 2: 93. https://doi.org/10.3390/cells8020093
APA StyleMartin, J. J., Woods, D. C., & Tilly, J. L. (2019). Implications and Current Limitations of Oogenesis from Female Germline or Oogonial Stem Cells in Adult Mammalian Ovaries. Cells, 8(2), 93. https://doi.org/10.3390/cells8020093