Hyaluronan-Based Hydrogel Scaffolds for Limbal Stem Cell Transplantation: A Review

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
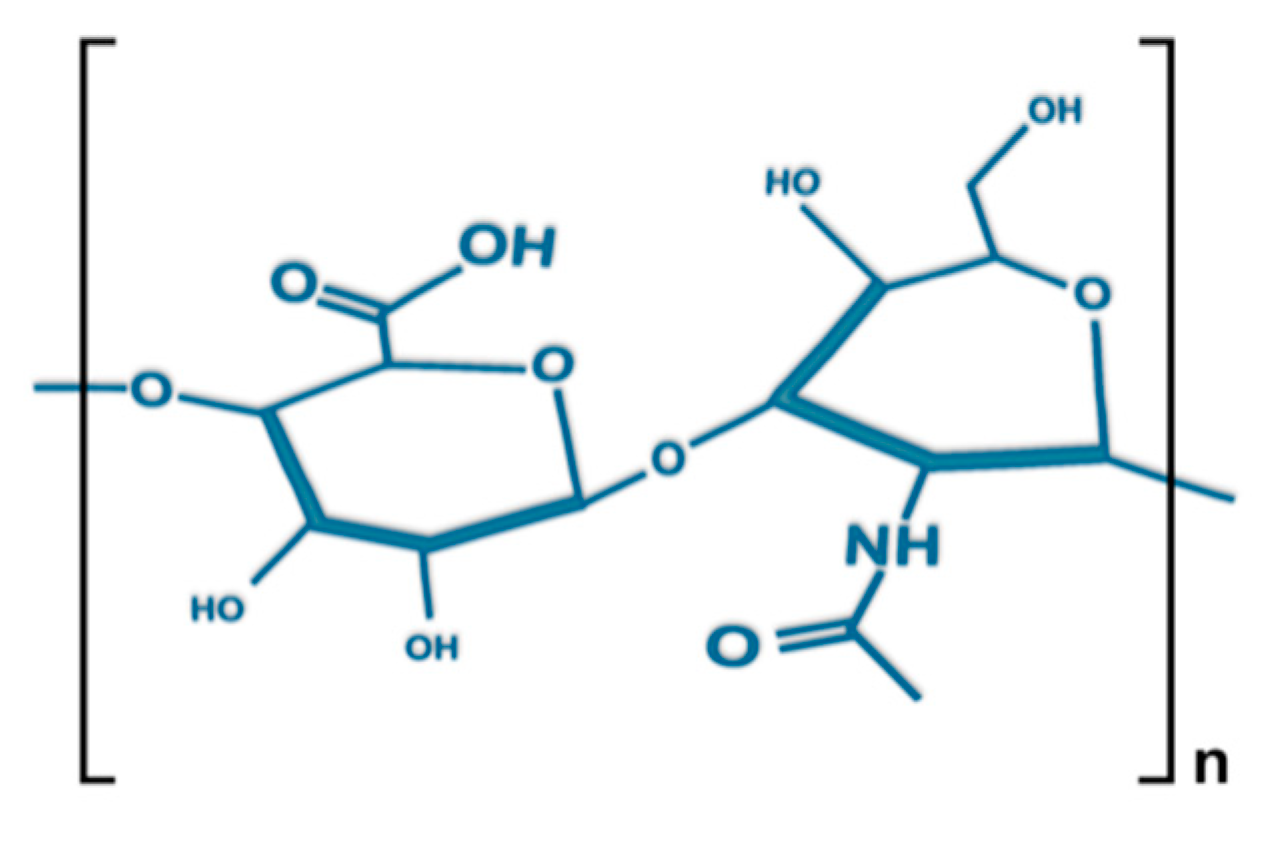
2. Structure, Size, Synthesis, and Degradation of Hyaluronan
3. Interactions of HA with ECM Components and Cell Receptor Proteins
4. Challenges in Designing HA-Hydrogel Scaffolds
5. Stem Cell Deficiency in the Cornea
6. Stem Cell-Based Therapy for Treatment of LSCD
7. HA-Based Hydrogel Scaffolds in the Treatment of LSCD
8. Conclusions and Future Work
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meyer, K.; Palmer, J.W. The polysaccharide of the vitreous humor. J. Biol. Chem. 1934, 107, 629–634. [Google Scholar]
- Balazs, E.A.; Laurent, T.C.; Jeanloz, R.W. Nomenclature of hyaluronic acid. Biochem. J. 1986, 235, 903. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.J.; Abatangelo, G. Functions of hyaluronan in wound repair. Wound Repair Regen. 1999, 7, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, A.S.; Pilling, D.; Gomer, R.H. High and low molecular weight hyaluronic acid differentially regulate human fibrocyte differentiation. PLoS ONE 2011, 6, e26078. [Google Scholar] [CrossRef]
- Kuo, J. Practical Aspects of Hyaluronan Based Medical Products; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Hascall, V.C.; Majors, A.K.; de la Motte, C.A.; Evanko, S.P.; Wang, A.; Drazba, J.A.; Strong, S.A.; Wight, T.N. Intracellular hyaluronan: A new frontier for inflammation? BBA-Gen. Subj. 2004, 1673, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Yamaguchi, M.; Sumitomo, S.; Takai, Y. Hyaluronan-based biomaterials in tissue engineering. Acta Histochem. Cytochem. 2004, 37, 1–5. [Google Scholar] [CrossRef]
- Necas, J.; Bartosikova, L.; Brauner, P.; Kolar, J. Hyaluronic acid (hyaluronan): A review. Vet. Med. 2008, 53, 397–411. [Google Scholar] [CrossRef]
- Evans, N.D.; Gentleman, E.; Polak, J.M. Scaffolds for stem cells. Mater. Today 2006, 9, 26–33. [Google Scholar] [CrossRef]
- Collins, M.N.; Birkinshaw, C. Hyaluronic acid based scaffolds for tissue engineering: A review. Carbohydr. Polym. 2013, 92, 1262–1279. [Google Scholar] [CrossRef]
- Stevens, M.M.; Marini, R.P.; Schaefer, D.; Aronson, J.; Langer, R.; Shastri, V.P. In vivo engineering of organs: The bone bioreactor. Proc. Natl. Acad. Sci. USA 2005, 102, 11450–11455. [Google Scholar] [CrossRef]
- Weinand, C.; Pomerantseva, I.; Neville, C.M.; Gupta, R.; Weinberg, E.; Madisch, I.; Shapiro, F.; Abukawa, H.; Troulis, M.J.; Vacanti, J.P. Hydrogel-β-TCP scaffolds and stem cells for tissue engineering bone. Bone 2006, 38, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; McAnulty, J.F.; Schurr, M.J.; Murphy, C.J.; Abbott, N.L. 8—Polymeric materials for chronic wound and burn dressings A2—Farrar, David. In Advanced Wound Repair Therapies; Woodhead Publishing: Sawston, UK, 2011; pp. 186–208. [Google Scholar]
- Burdick, J.A.; Prestwich, G.D. Hyaluronic acid hydrogels for biomedical applications. Adv. Mater. 2011, 23, H41–H56. [Google Scholar] [CrossRef] [PubMed]
- Dai Prè, E.; Conti, G.; Sbarbati, A. Hyaluronic acid (Ha) scaffolds and multipotent stromal cells (Mscs) in regenerative medicine. Stem Cell Rev. Rep. 2016, 12, 664–681. [Google Scholar] [CrossRef]
- O’Callaghan, A.R.; Daniels, J.T. Concise review: Limbal epithelial stem cell therapy: Controversies and challenges. Stem Cells 2011, 29, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
- Sejpal, K.; Bakhtiari, P.; Deng, S.X. Presentation, diagnosis and management of limbal stem cell deficiency. Middle East Afr. J. Ophthalmol. 2013, 20, 5. [Google Scholar]
- Ahmad, S. Concise review: Limbal stem cell deficiency, dysfunction, and distress. Stem Cell Transl. Med. 2012, 1, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Gesteira, T.F.; Sun, M.; Coulson-Thomas, Y.M.; Yamaguchi, Y.; Yeh, L.-K.; Hascall, V.; Coulson-Thomas, V.J. Hyaluronan rich microenvironment in the limbal stem cell niche regulates limbal stem cell differentiation. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4407–4421. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.E.; Heatley, F. Biological properties of hyaluronan in aqueous solution are controlled and sequestered by reversible tertiary structures, defined by NMR spectroscopy. Biomacromolecules 2002, 3, 547–553. [Google Scholar] [CrossRef]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528. [Google Scholar] [CrossRef]
- Day, A.J.; Sheehan, J.K. Hyaluronan: Polysaccharide chaos to protein organisation. Curr. Opin. Struct. Biol. 2001, 11, 617–622. [Google Scholar] [CrossRef]
- Cowman, M.K.; Matsuoka, S. The intrinsic viscosity of hyaluronan. In Hyaluronan; Elsevier: Amsterdam, The Netherlands, 2002; pp. 75–78. [Google Scholar]
- Cowman, M.K.; Spagnoli, C.; Kudasheva, D.; Li, M.; Dyal, A.; Kanai, S.; Balazs, E.A. Extended, relaxed, and condensed conformations of hyaluronan observed by atomic force microscopy. Biophys. J. 2005, 88, 590–602. [Google Scholar] [CrossRef] [PubMed]
- Weissmann, B.; Meyer, K. The structure of hyalobiuronic acid and of hyaluronic acid from umbilical Cord1, 2. J. Am. Chem. Soc. 1954, 76, 1753–1757. [Google Scholar] [CrossRef]
- McKee, C.M.; Penno, M.B.; Cowman, M.; Burdick, M.D.; Strieter, R.M.; Bao, C.; Noble, P.W. Hyaluronan (HA) fragments induce chemokine gene expression in alveolar macrophages. The role of HA size and CD44. J. Clin. Investig. 1996, 98, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Turley, E.A.; Noble, P.W.; Bourguignon, L.Y. Signaling properties of hyaluronan receptors. J. Biol. Chem. 2002, 277, 4589–4592. [Google Scholar] [CrossRef] [PubMed]
- Altman, R.; Bedi, A.; Manjoo, A.; Niazi, F.; Shaw, P.; Mease, P. Anti-inflammatory effects of intra-articular hyaluronic acid: A systematic review. Cartilage 2019, 10, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Skandalis, S.S.; Tzanakakis, G.N.; Karamanos, N.K. Proteoglycans in health and disease: Novel roles for proteoglycans in malignancy and their pharmacological targeting. FEBS J. 2010, 277, 3904–3923. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.; Stern, R. Chain gangs: New aspects of hyaluronan metabolism. Biochem. Res. Int. 2012, 2012, 893947. [Google Scholar] [CrossRef]
- Siiskonen, H.; Oikari, S.; Pasonen-Seppänen, S.; Rilla, K. Hyaluronan synthase 1: A mysterious enzyme with unexpected functions. Front. Mmunol. 2015, 6, 43. [Google Scholar] [CrossRef]
- Lee, J.Y.; Spicer, A.P. Hyaluronan: A multifunctional, megaDalton, stealth molecule. Curr. Opin. Cell Biol. 2000, 12, 581–586. [Google Scholar] [CrossRef]
- Jacobson, A.; Brinck, J.; Briskin, M.J.; Spicer, A.P.; Heldin, P. Expression of human hyaluronan synthases in response to external stimuli. Biochem. J. 2000, 348, 29–35. [Google Scholar] [CrossRef]
- Vigetti, D.; Viola, M.; Karousou, E.; Deleonibus, S.; Karamanou, K.; De Luca, G.; Passi, A. Epigenetics in extracellular matrix remodeling and hyaluronan metabolism. FEBS J. 2014, 281, 4980–4992. [Google Scholar] [CrossRef]
- Lokeshwar, V.B.; Gomez, P.; Kramer, M.; Knapp, J.; McCornack, M.A.; Lopez, L.E.; Fregien, N.; Dhir, N.; Scherer, S.; Klumpp, D.J. Epigenetic regulation of HYAL-1 hyaluronidase expression identification of HYAL-1 promoter. J. Biol. Chem. 2008, 283, 29215–29227. [Google Scholar] [CrossRef]
- Buhren, B.A.; Schrumpf, H.; Hoff, N.-P.; Bölke, E.; Hilton, S.; Gerber, P.A. Hyaluronidase: From clinical applications to molecular and cellular mechanisms. Eur. J. Med. Res. 2016, 21, 5. [Google Scholar] [CrossRef]
- Csoka, A.B.; Frost, G.I.; Stern, R. The six hyaluronidase-like genes in the human and mouse genomes. Matrix Biol. 2001, 20, 499–508. [Google Scholar] [CrossRef]
- Stern, R.; Jedrzejas, M.J. Hyaluronidases: Their genomics, structures, and mechanisms of action. Chem. Rev. 2006, 106, 818–839. [Google Scholar] [CrossRef]
- Laurent, T.C.; Fraser, J.R.E. The properties and turnover of hyaluronan. In Functions of the Proteoglycans; Evered, D., Whelan, J., Eds.; A Wiley—Interscience Publication: London, UK, 1986; Volume 124, pp. 9–29. [Google Scholar]
- Stern, R. Hyaluronan catabolism: A new metabolic pathway. Eur. J. Cell Biol. 2004, 83, 317–325. [Google Scholar] [CrossRef]
- Lennon, F.E.; Singleton, P.A. Role of hyaluronan and hyaluronan-binding proteins in lung pathobiology. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L137–L147. [Google Scholar] [CrossRef]
- Izawa, N.; Serata, M.; Sone, T.; Omasa, T.; Ohtake, H. Hyaluronic acid production by recombinant Streptococcus thermophilus. J. Biosci. Bioeng. 2011, 111, 665–670. [Google Scholar] [CrossRef]
- Kharkar, P.M.; Kiick, K.L.; Kloxin, A.M. Designing degradable hydrogels for orthogonal control of cell microenvironments. Chem. Soc. Rev. 2013, 42, 7335–7372. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef]
- Järveläinen, H.; Sainio, A.; Koulu, M.; Wight, T.N.; Penttinen, R. Extracellular matrix molecules: Potential targets in pharmacotherapy. Pharmacol. Rev. 2009, 61, 198–223. [Google Scholar] [CrossRef] [PubMed]
- Hubmacher, D.; Apte, S.S. The biology of the extracellular matrix: Novel insights. Curr. Opin. Rheumatol. 2013, 25, 65. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Rasch, M.G.; Weaver, V.M. Dynamic interplay between the collagen scaffold and tumor evolution. Curr. Opin. Cell Biol. 2010, 22, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O.; Naba, A. Overview of the matrisome—An inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef] [PubMed]
- Wight, T.N.; Mecham, R.P. Biology of Proteoglycans; Academic Press: Orlando, FL, USA, 2013. [Google Scholar]
- Yanagishita, M. Function of proteoglycans in the extracellular matrix. Pathol. Int. 1993, 43, 283–293. [Google Scholar] [CrossRef]
- Ricard-Blum, S. Protein–glycosaminoglycan interaction networks: Focus on heparan sulfate. Perspect. Sci. 2017, 11, 62–69. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; Prestwich, G.D. Hyaluronan-binding proteins: Tying up the giant. J. Biol. Chem. 2002, 277, 4585–4588. [Google Scholar] [CrossRef]
- Kohda, D.; Morton, C.J.; Parkar, A.A.; Hatanaka, H.; Inagaki, F.M.; Campbell, I.D.; Day, A.J. Solution structure of the link module: A hyaluronan-binding domain involved in extracellular matrix stability and cell migration. Cell 1996, 86, 767–775. [Google Scholar] [CrossRef]
- Day, A. The Structure and Regulation of Hyaluronan-Binding Proteins; Portland Press Limited: London, UK, 1999. [Google Scholar]
- Ziebell, M.R. Ligand Binding and Structural Characterization of a Recombinant Hyaluronic Acid Binding Domain; State University of New York: Stony Brook, NY, USA, 1999. [Google Scholar]
- Kuo, J.; Prestwich, G. Materials of biological origin—Materials analysis and implant uses. In Comprehensive Biomaterials; Ducheyne, P., Healy, K., Hutmacher, D., Kirkpatrick, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Roberts, J.J.; Elder, R.M.; Neumann, A.J.; Jayaraman, A.; Bryant, S.J. Interaction of hyaluronan binding peptides with glycosaminoglycans in poly (ethylene glycol) hydrogels. Biomacromolecules 2014, 15, 1132–1141. [Google Scholar] [CrossRef]
- Segura, T.; Anderson, B.C.; Chung, P.H.; Webber, R.E.; Shull, K.R.; Shea, L.D. Crosslinked hyaluronic acid hydrogels: A strategy to functionalize and pattern. Biomaterials 2005, 26, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Borzacchiello, A.; Russo, L.; Malle, B.M.; Schwach-Abdellaoui, K.; Ambrosio, L. Hyaluronic acid based hydrogels for regenerative medicine applications. BioMed Res. Int. 2015, 2015, 871218. [Google Scholar] [CrossRef] [PubMed]
- Notara, M.; Alatza, A.; Gilfillan, J.; Harris, A.R.; Levis, H.J.; Schrader, S.; Vernon, A.; Daniels, J.T. In sickness and in health: Corneal epithelial stem cell biology, pathology and therapy. Exp. Eye Res. 2010, 90, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Schermer, A.; Galvin, S.; Sun, T.T. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J. Cell Biol. 1986, 103, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Dua, H.S.; Azuara-Blanco, A. Limbal stem cells of the corneal epithelium. Surv. Ophthalmol. 2000, 44, 415–425. [Google Scholar] [CrossRef]
- Utheim, T.P. Limbal epithelial cell therapy: Past, present, and future. In Corneal Regenerative Medicine; Springer: New York, NY, USA, 2013; pp. 3–43. [Google Scholar]
- He, H.; Yiu, S.C. Stem cell-based therapy for treating limbal stem cells deficiency: A review of different strategies. Saudi J. Ophthalmol. 2014, 28, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Ramaesh, T.; Collinson, J.M.; Ramaesh, K.; Kaufman, M.H.; West, J.D.; Dhillon, B. Corneal abnormalities in Pax6+/− small eye mice mimic human aniridia-related keratopathy. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1871–1878. [Google Scholar] [CrossRef]
- Trosan, P.; Svobodova, E.; Chudickova, M.; Krulova, M.; Zajicova, A.; Holan, V. The key role of insulin-like growth factor I in limbal stem cell differentiation and the corneal wound-healing process. Stem Cells Dev. 2012, 21, 3341–3350. [Google Scholar] [CrossRef] [PubMed]
- Klenkler, B.; Sheardown, H. Growth factors in the anterior segment: Role in tissue maintenance, wound healing and ocular pathology. Exp. Eye Res. 2004, 79, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, M.; Gorivodsky, M.; Shtrom, S.; Grinberg, A.; Niehrs, C.; Morasso, M.I.; Westphal, H. Dkk2 plays an essential role in the corneal fate of the ocular surface epithelium. Development 2006, 133, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Takács, L.; Tóth, E.; Losonczy, G.; Szanto, A.; Bähr-Ivacevic, T.; Benes, V.; Berta, A.; Vereb, G. Differentially expressed genes associated with human limbal epithelial phenotypes: New molecules that potentially facilitate selection of stem cell-enriched populations. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1252–1260. [Google Scholar] [CrossRef]
- Atallah, M.R.; Palioura, S.; Perez, V.L.; Amescua, G. Limbal stem cell transplantation: Current perspectives. Clin. Ophthalmol. 2016, 10, 593. [Google Scholar]
- Sehic, A.; Utheim, Ø.; Ommundsen, K.; Utheim, T. Pre-clinical cell-based therapy for limbal stem cell deficiency. J. Funct Biomater. 2015, 6, 863–888. [Google Scholar] [CrossRef]
- Daya, S.M.; Watson, A.; Sharpe, J.R.; Giledi, O.; Rowe, A.; Martin, R.; James, S.E. Outcomes and DNA analysis of ex vivo expanded stem cell allograft for ocular surface reconstruction. Ophthalmology 2005, 112, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.A.; Brereton, H.M.; Aggarwal, R.; Sykes, P.J.; Turner, D.R.; Russ, G.R.; Coster, D.J. Use of DNA polymorphisms and the polymerase chain reaction to examine the survival of a human limbal stem cell allograft. Am. J. Ophthalmol. 1995, 120, 342–350. [Google Scholar] [CrossRef]
- Shimazaki, J.; Kaido, M.; Shinozaki, N.; Shimmura, S.; Munkhbat, B.; Hagihara, M.; Tsuji, K.; Tsubota, K. Evidence of long-term survival of donor-derived cells after limbal allograft transplantation. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1664–1668. [Google Scholar]
- Henderson, T.; McCall, S.; Taylor, G.; Noble, B. Do transplanted corneal limbal stem cells survive in vivo long term? Possible techniques to detect donor cell survival by polymerase chain reaction with the amelogenin gene and Y-specific probes. Eye 1997, 11, 779. [Google Scholar] [CrossRef]
- Djalilian, A.R.; Mahesh, S.P.; Koch, C.A.; Nussenblatt, R.B.; Shen, D.; Zhuang, Z.; Holland, E.J.; Chan, C.C. Survival of donor epithelial cells after limbal stem cell transplantation. Investig. Ophthalmol. Vis. Sci. 2005, 46, 803–807. [Google Scholar] [CrossRef]
- Utheim, T.P.; Utheim, Ø.A.; Khan, Q.-E.; Sehic, A. Culture of oral mucosal epithelial cells for the purpose of treating limbal stem cell deficiency. J. Funct Biomater. 2016, 7, 5. [Google Scholar] [CrossRef]
- Levis, H.; Daniels, J.T. New technologies in limbal epithelial stem cell transplantation. Curr. Opin. Biotechnol. 2009, 20, 593–597. [Google Scholar] [CrossRef]
- Nguyen, K.N.; Bobba, S.; Richardson, A.; Park, M.; Watson, S.L.; Wakefield, D.; Di Girolamo, N. Native and synthetic scaffolds for limbal epithelial stem cell transplantation. Acta Biomater. 2018, 65, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.-Y. Hyaluronic acid concentration-mediated changes in structure and function of porous carriers for corneal endothelial cell sheet delivery. Mater. Sci. Eng. C 2016, 59, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.; Amankwah, R.; Powell-Richards, A.; Dua, H. Sodium hyaluronate (hyaluronic acid) promotes migration of human corneal epithelial cells in vitro. Br. J. Ophthalmol. 2004, 88, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Fiorica, C.; Senior, R.A.; Pitarresi, G.; Palumbo, F.S.; Giammona, G.; Deshpande, P.; MacNeil, S. Biocompatible hydrogels based on hyaluronic acid cross-linked with a polyaspartamide derivative as delivery systems for epithelial limbal cells. Int. J. Pharm. 2011, 414, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Kiiskinen, J. Co-Culture of Corneal Epithelial Cells and Adipose Stem Cells-Towards the Use of Hydrogels in Ocular Surface Reconstruction; University of Tampere: Tampere, Finland, 2016; p. 58. [Google Scholar]
- Chen, D.; Qu, Y.; Hua, X.; Zhang, L.; Liu, Z.; Pflugfelder, S.; Li, D. A hyaluronan hydrogel scaffold-based xeno-free culture system for ex vivo expansion of human corneal epithelial stem cells. Eye 2017, 31, 962. [Google Scholar] [CrossRef] [PubMed]
- Vanderhooft, J.L.; Alcoutlabi, M.; Magda, J.J.; Prestwich, G.D. Rheological Properties of Cross-Linked Hyaluronan–Gelatin Hydrogels for Tissue Engineering. Macromol. Biosci. 2009, 9, 20–28. [Google Scholar] [CrossRef]
- Morra, M. Engineering of biomaterials surfaces by hyaluronan. Biomacromolecules 2005, 6, 1205–1223. [Google Scholar] [CrossRef]
- Berriaud, N.; Milas, M.; Rinaudo, M. Characterization and properties of hyaluronic acid (hyaluronan). In Polysaccharides in Medicine and Biotechnology; Severian, D., Ed.; Marcel Dekker: New York, NY, USA, 1998; pp. 313–334. [Google Scholar]
- Geckil, H.; Xu, F.; Zhang, X.; Moon, S.; Demirci, U. Engineering hydrogels as extracellular matrix mimics. Nanomedicine 2010, 5, 469–484. [Google Scholar] [CrossRef]
- McDonald, C.; Kaye, S.; Figueiredo, F.; Macintosh, G.; Lockett, C. A randomised, crossover, multicentre study to compare the performance of 0.1%(w/v) sodium hyaluronate with 1.4%(w/v) polyvinyl alcohol in the alleviation of symptoms associated with dry eye syndrome. Eye 2002, 16, 601. [Google Scholar] [CrossRef]
- Jeng, B.H.; Hoyt, C.S.; McLeod, S.D. Completion rate of continuous curvilinear capsulorhexis in pediatric cataract surgery using different viscoelastic materials. J. Cataract Refract. Surg. 2004, 30, 85–88. [Google Scholar] [CrossRef]
- Shimmura, S.; Shimazaki, J.; Omoto, M.; Teruya, A.; Ishioka, M.; Tsubota, K. Deep lamellar keratoplasty (DLKP) in keratoconus patients using viscoadaptive viscoelastics. Cornea 2005, 24, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.-Y.; Tu, I.-H. Adhesion, phenotypic expression, and biosynthetic capacity of corneal keratocytes on surfaces coated with hyaluronic acid of different molecular weights. Acta Biomater. 2012, 8, 1068–1079. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yazdani, M.; Shahdadfar, A.; Jackson, C.J.; Utheim, T.P. Hyaluronan-Based Hydrogel Scaffolds for Limbal Stem Cell Transplantation: A Review. Cells 2019, 8, 245. https://doi.org/10.3390/cells8030245
Yazdani M, Shahdadfar A, Jackson CJ, Utheim TP. Hyaluronan-Based Hydrogel Scaffolds for Limbal Stem Cell Transplantation: A Review. Cells. 2019; 8(3):245. https://doi.org/10.3390/cells8030245
Chicago/Turabian StyleYazdani, Mazyar, Aboulghassem Shahdadfar, Catherine Joan Jackson, and Tor Paaske Utheim. 2019. "Hyaluronan-Based Hydrogel Scaffolds for Limbal Stem Cell Transplantation: A Review" Cells 8, no. 3: 245. https://doi.org/10.3390/cells8030245
APA StyleYazdani, M., Shahdadfar, A., Jackson, C. J., & Utheim, T. P. (2019). Hyaluronan-Based Hydrogel Scaffolds for Limbal Stem Cell Transplantation: A Review. Cells, 8(3), 245. https://doi.org/10.3390/cells8030245