Novel Flavivirus Antiviral That Targets the Host Nuclear Transport Importin α/β1 Heterodimer

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Inhibitors
2.2. Cell Culture and Virus Propagation
2.3. Protein Expression
2.4. AlphaScreen
2.5. Thermostability Assay
2.6. Immunofluorescence
2.7. Circular Dichroism Spectroscopy
2.8. Analytical Ultracentrifugation
2.9. Cell Cytotoxicity Assay
2.10. Plaque Assay
2.11. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.12. Quantification and Statistical Analysis
3. Results
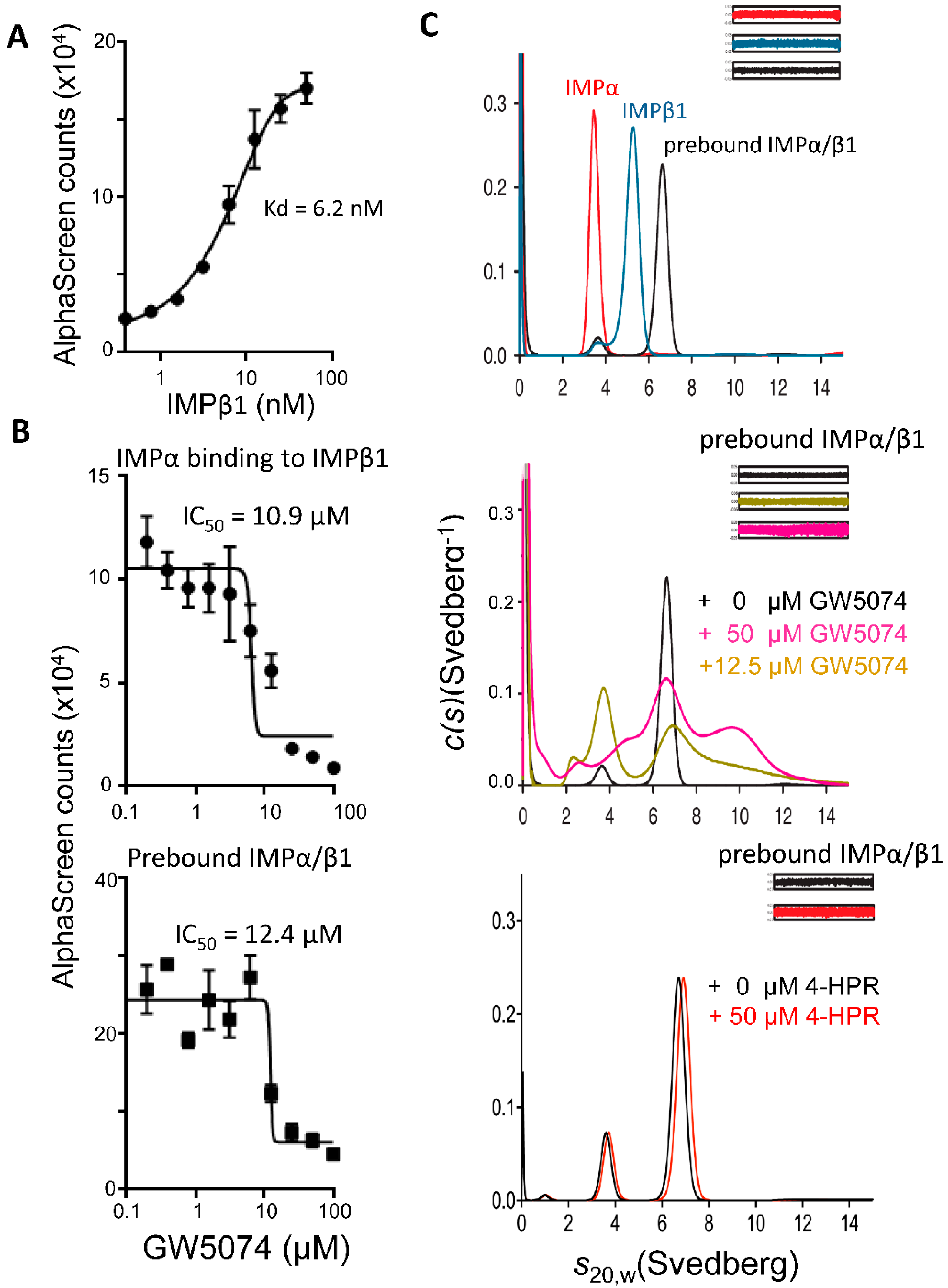
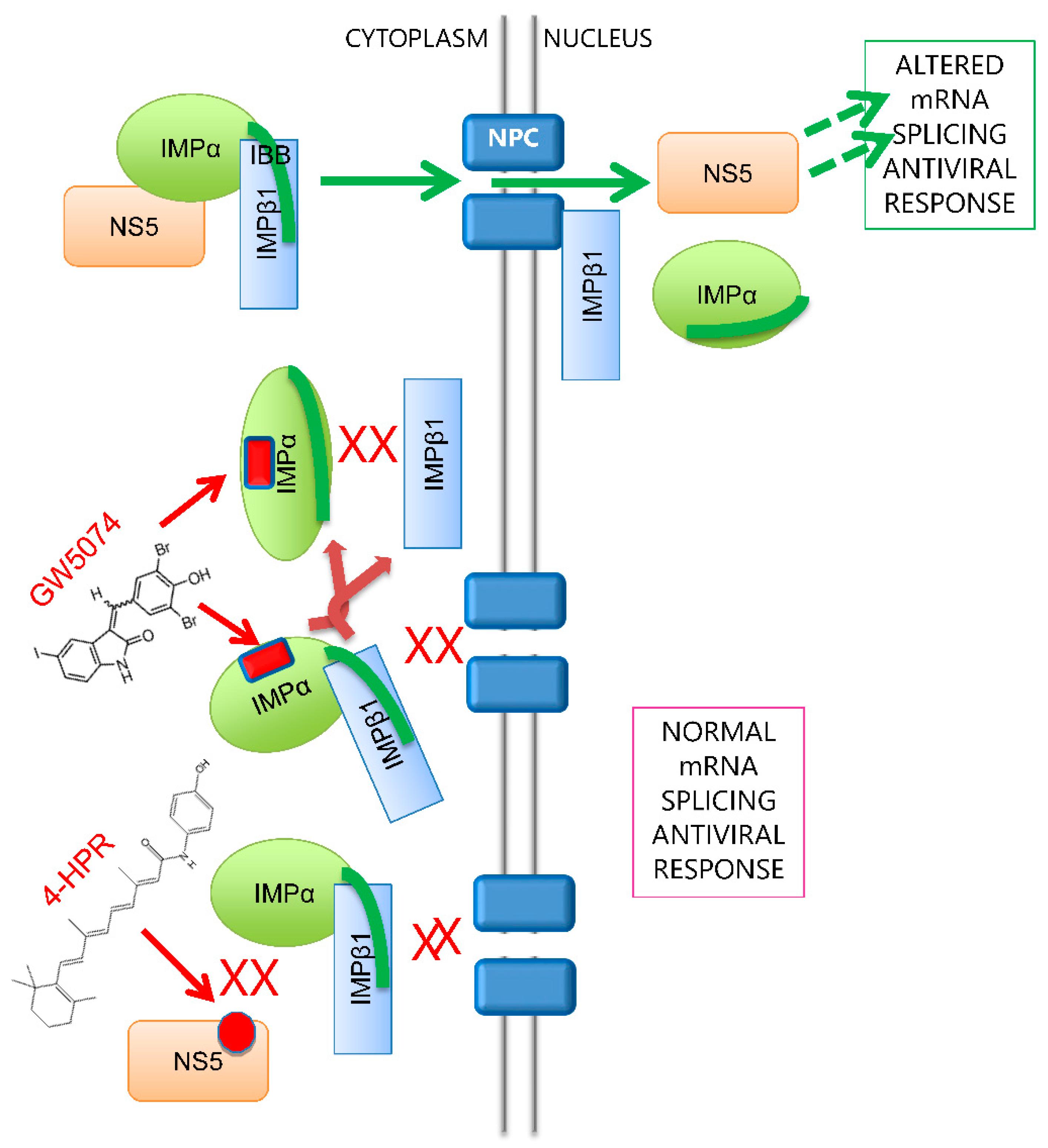
3.1. GW5074 Inhibits DENV NS5–IMP Binding and NS5 Nuclear Localisation in Infected Cells
3.2. GW5074 Can Inhibit Binding of IMPα to IMPβ1 and Dissociate the IMPα/β1 Heterodimer
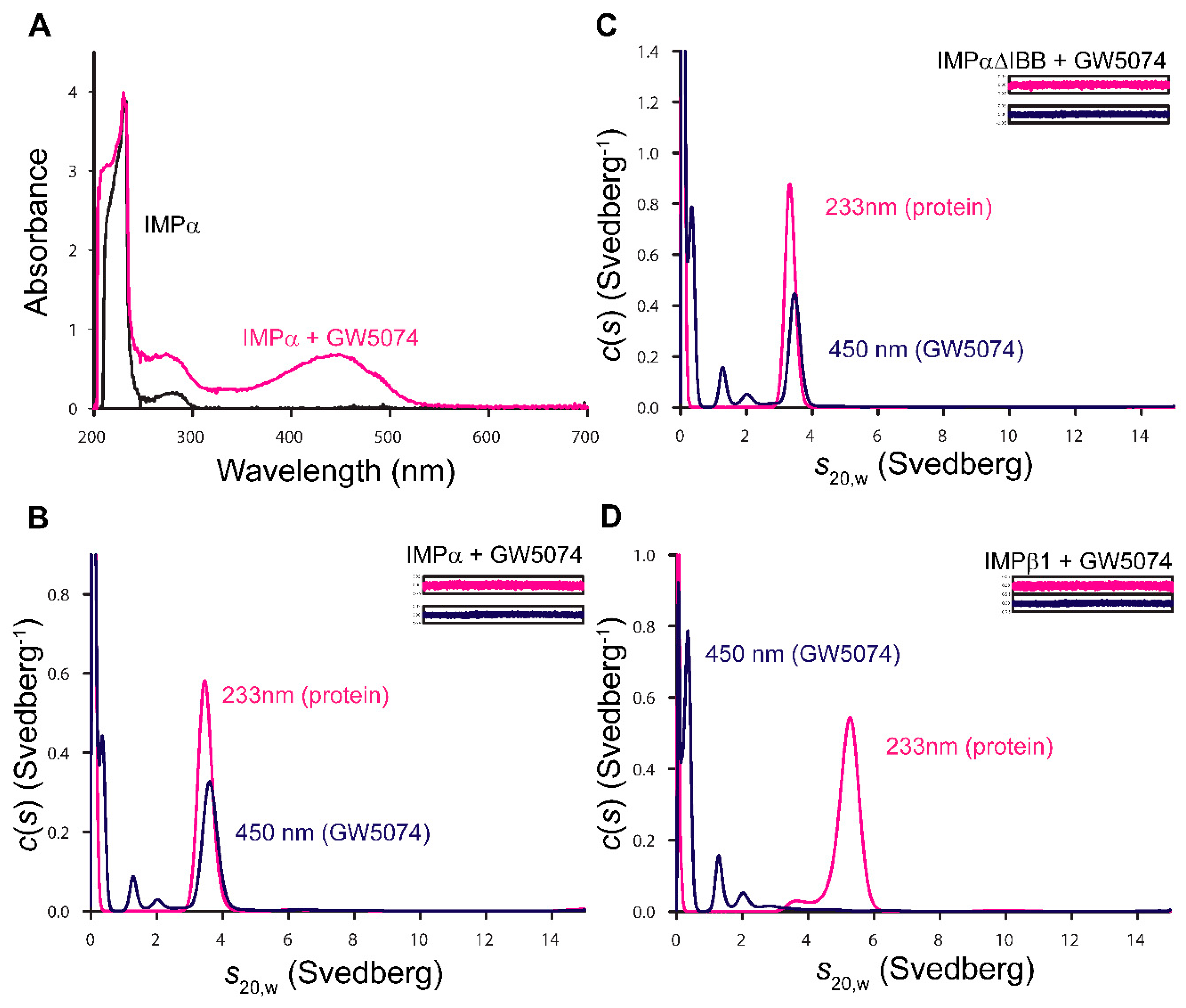
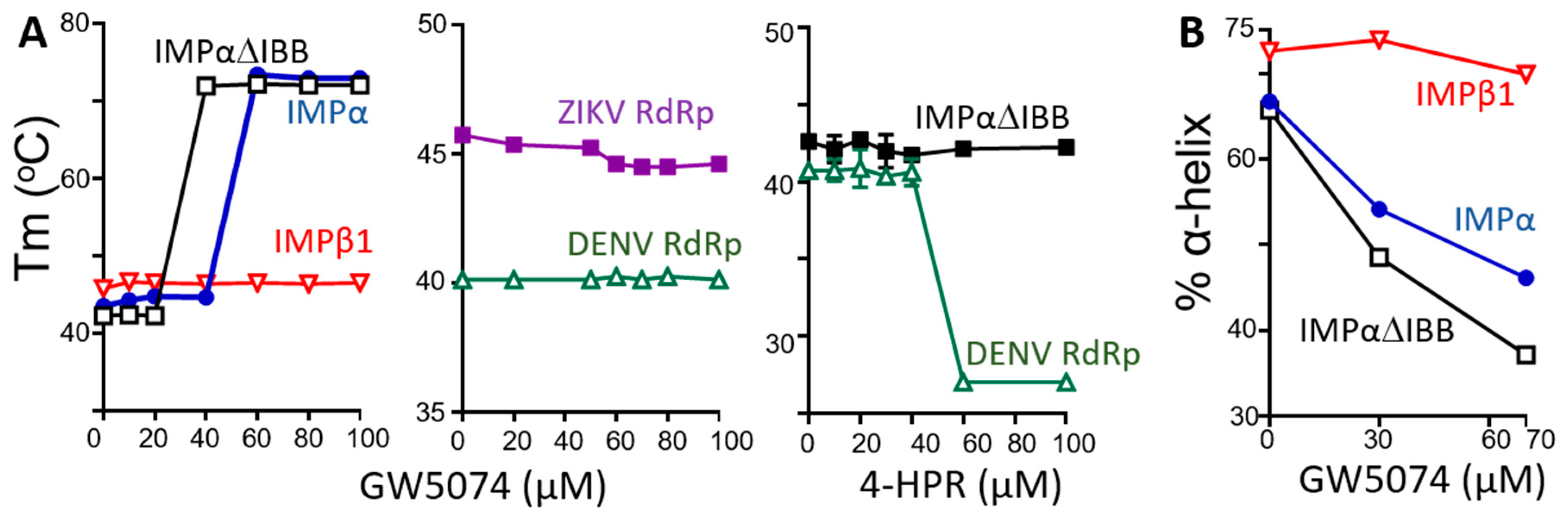
3.3. GW5074 Binds Directly to the IMPα ARM Repeat Domain with Effects on Thermostability and Conformation
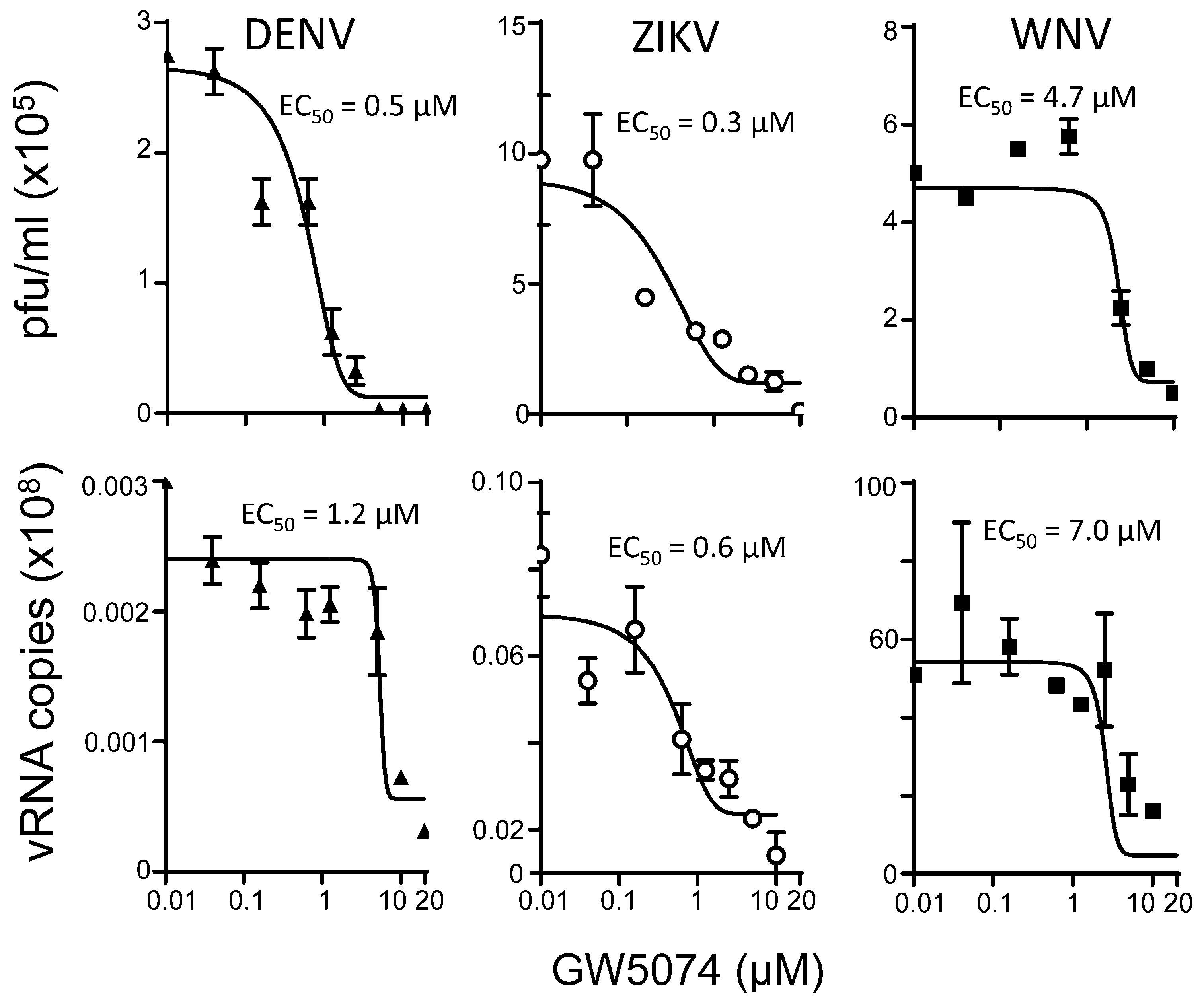
3.4. GW5074 Is A Potent Anti-Flavivirus Agent
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gubler, D.J. The global emergence/resurgence of arboviral diseases as public health problems. Arch. Med. Res. 2002, 33, 330–342. [Google Scholar] [CrossRef]
- Villar, L.; Dayan, G.H.; Arredondo-Garcia, J.L.; Rivera, D.M.; Cunha, R.; Deseda, C.; Reynales, H.; Costa, M.S.; Morales-Ramirez, J.O.; Carrasquilla, G.; et al. Efficacy of a tetravalent dengue vaccine in children in Latin America. N. Engl. J. Med. 2015, 372, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Coudeville, L.; Baurin, N.; L’Azou, M.; Guy, B. Potential impact of dengue vaccination: Insights from two large-scale phase III trials with a tetravalent dengue vaccine. Vaccine 2016, 34, 6426–6435. [Google Scholar] [CrossRef] [Green Version]
- Moureau, G.; Cook, S.; Lemey, P.; Nougairede, A.; Forrester, N.L.; Khasnatinov, M.; Charrel, R.N.; Firth, A.E.; Gould, E.A.; de Lamballerie, X. New Insights into Flavivirus Evolution, Taxonomy and Biogeographic History, Extended by Analysis of Canonical and Alternative Coding Sequences. PLoS ONE 2015, 10, e0117849. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Jans, D.A. Nucleocytoplasmic Trafficking of Dengue Non-structural Protein 5 as a Target for Antivirals. Adv. Exp. Med. Biol. 2018, 1062, 199–213. [Google Scholar]
- El Sahili, A.; Lescar, J. Dengue Virus Non-Structural Protein 5. Viruses 2017, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- Pryor, M.J.; Rawlinson, S.M.; Butcher, R.E.; Barton, C.L.; Waterhouse, T.A.; Vasudevan, S.G.; Bardin, P.G.; Wright, P.J.; Jans, D.A.; Davidson, A.D. Nuclear localization of dengue virus nonstructural protein 5 through its importin α/β-recognized nuclear localization sequences is integral to viral infection. Traffic 2007, 8, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Jans, D.A.; Martin, A.; Wagstaff, K.M. Inhibitors of Nuclear Transport. Curr. Opin. Cell Biol. 2019, 58, 50–60. [Google Scholar] [CrossRef]
- Fulcher, A.; Jans, D.A. Regulation of nucleocytoplasmic trafficking of viral proteins; an integral role in pathogenesis? Biochem. Biophys. Acta Mol. Cell Res. 2011, 1813, 2176–2190. [Google Scholar] [CrossRef]
- Audsley, M.D.; Jans, D.A.; Moseley, G.W. Roles of nuclear trafficking in infection by cytoplasmic negative strand RNA viruses: Paramyxoviruses and beyond. J. Gen. Virol. 2016, 97, 2463–2481. [Google Scholar] [CrossRef]
- Forwood, J.K.; Brooks, A.; Briggs, L.J.; Xiao, C.Y.; Jans, D.A.; Vasudevan, S.G. The 37-amino-acid interdomain of dengue virus NS5 protein contains a functional NLS and inhibitory CK2 site. Biochem. Biophys. Res. Commun. 1999, 257, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Tay, M.Y.; Fraser, J.E.; Chan, W.K.; Moreland, N.J.; Rathore, A.P.; Wang, C.; Vasudevan, S.G.; Jans, D.A. Nuclear localization of dengue virus (DENV) 1-4 non-structural protein 5; protection against all 4 DENV serotypes by the inhibitor Ivermectin. Antivir. Res. 2013, 99, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, K.M.; Sivakumaran, H.; Heaton, S.M.; Harrich, D.; Jans, D.A. Ivermectin is a specific inhibitor of importin α/β-mediated nuclear import able to inhibit replication of HIV-1 and dengue virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef] [Green Version]
- Fraser, J.E.; Watanabe, S.; Wang, C.; Chan, W.K.; Maher, B.; Lopez-Denman, A.; Hick, C.; Wagstaff, K.M.; Mackenzie, J.M.; Sexton, P.M.; et al. A nuclear transport inhibitor that modulates the unfolded protein response and provides in vivo protection against lethal dengue virus infection. J. Infect. Dis. 2014, 210, 1780–1791. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Denman, A.J.; Russo, A.; Wagstaff, K.M.; White, P.A.; Jans, D.A.; Mackenzie, J.M. Nucleocytoplasmic shuttling of the West Nile virus RNA-dependent RNA polymerase NS5 is critical to infection. Cell. Microbiol. 2018, 20, e12848. [Google Scholar] [CrossRef]
- Wang, C.; Yang, S.N.Y.; Smith, K.; Forwood, J.K.; Jans, D.A. Nuclear import inhibitor N-(4-hydroxyphenyl) retinamide targets Zika virus (ZIKV) nonstructural protein 5 to inhibit ZIKV infection. Biochem. Biophys. Res. Commun. 2017, 493, 1555–1559. [Google Scholar] [CrossRef] [PubMed]
- Rawlinson, S.M.; Pryor, M.J.; Wright, P.J.; Jans, D.A. CRM1-mediated nuclear export of dengue virus RNA polymerase NS5 modulates interleukin-8 induction and virus production. J. Biol. Chem. 2009, 284, 15589–15597. [Google Scholar] [CrossRef] [PubMed]
- Medin, C.L.; Fitzgerald, K.A.; Rothman, A.L. Dengue virus nonstructural protein NS5 induces interleukin-8 transcription and secretion. J. Virol. 2005, 79, 11053–11061. [Google Scholar] [CrossRef] [PubMed]
- De Maio, F.A.; Risso, G.; Iglesias, N.G.; Shah, P.; Pozzi, B.; Gebhard, L.G.; Mammi, P.; Mancini, E.; Yanovsky, M.J.; Andino, R.; et al. The Dengue Virus NS5 Protein Intrudes in the Cellular Spliceosome and Modulates Splicing. PLoS Pathog. 2016, 12, e1005841. [Google Scholar] [CrossRef]
- Wagstaff, K.M.; Rawlinson, S.M.; Hearps, A.C.; Jans, D.A. An AScreen(R)-based assay for high-throughput screening for specific inhibitors of nuclear import. J. Biomol. Screen. 2011, 16, 192–200. [Google Scholar] [CrossRef]
- Lundberg, L.; Pinkham, C.; Baer, A.; Amaya, M.; Narayanan, A.; Wagstaff, K.M.; Jans, D.A.; Kehn-Hall, K. Nuclear import and export inhibitors alter capsid protein distribution in mammalian cells and reduce Venezuelan Equine Encephalitis Virus replication. Antivir. Res. 2013, 100, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Gras, S.; Wilmann, P.G.; Chen, Z.; Halim, H.; Liu, Y.C.; Kjer-Nielsen, L.; Purcell, A.W.; Burrows, S.R.; McCluskey, J.; Rossjohn, J. A structural basis for varied αβ TCR usage against an immunodominant EBV antigen restricted to a HLA-B8 molecule. J. Immunol. 2012, 188, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Munoz, V.; Blanco, F.J.; Serrano, L. The distribution of α-helix propensity along the polypeptide chain is not conserved in proteins from the same family. Protein Sci. 1995, 4, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Laue, T.M.; Shah, B.D.; Ridgeway, T.M.; Pelletier, S.L. Computer-aided interpretation of analytical sedimentation data for proteins. In Analytical Ultracentfiugation in Biochemistry and Polymer Science; The Royal Society of Chemistry: Cambridge, UK, 1992; pp. 90–125. [Google Scholar]
- Atkinson, S.C.; Audsley, M.D.; Lieu, K.G.; Marsh, G.A.; Thomas, D.R.; Heaton, S.M.; Paxman, J.J.; Wagstaff, K.M.; Buckle, A.M.; Moseley, G.W.; et al. Recognition by host nuclear transport proteins drives disorder-to-order transition in Hendra virus V. Sci. Rep. 2018, 8, 358. [Google Scholar] [CrossRef] [PubMed]
- Schuck, P. Size-distribution analysis of macromolecules by sedimentation velocity ultracentrifugation and lamm equation modeling. Biophys. J. 2000, 78, 1606–1619. [Google Scholar] [CrossRef]
- Lo, M.C.; Aulabaugh, A.; Jin, G.; Cowling, R.; Bard, J.; Malamas, M.; Ellestad, G. Evaluation of fluorescence-based thermal shift assays for hit identification in drug discovery. Anal. Biochem. 2004, 332, 153–159. [Google Scholar] [CrossRef]
- Matulis, D.; Kranz, J.K.; Salemme, F.R.; Todd, M.J. Thermodynamic stability of carbonic anhydrase: Measurements of binding affinity and stoichiometry using ThermoFluor. Biochemistry 2005, 44, 5258–5266. [Google Scholar] [CrossRef]
- Kobe, B. Autoinhibition by an internal nuclear localization signal revealed by the crystal structure of mammalian importin α. Nat. Struct. Biol. 1999, 6, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Malet, H.; Egloff, M.P.; Selisko, B.; Butcher, R.E.; Wright, P.J.; Roberts, M.; Gruez, A.; Sulzenbacher, G.; Vonrhein, C.; Bricogne, G.; et al. Crystal structure of the RNA polymerase domain of the West Nile virus non-structural protein 5. J. Biol. Chem. 2007, 282, 10678–10689. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, K.M.; Headey, S.; Telwatte, S.; Tyssen, D.; Hearps, A.C.; Thomas, D.R.; Tachedjian, G.; Jans, D.A. Molecular dissection of an inhibitor targeting the HIV integrase dependent preintegration complex nuclear import. Cell. Microbiol. 2018, e12953. [Google Scholar] [CrossRef] [PubMed]
- Ford Siltz, L.A.; Viktorova, E.G.; Zhang, B.; Kouiavskaia, D.; Dragunsky, E.; Chumakov, K.; Isaacs, L.; Belov, G.A. New small-molecule inhibitors effectively blocking picornavirus replication. J. Virol. 2014, 88, 11091–11107. [Google Scholar] [CrossRef]
- Medin, C.L.; Rothman, A.L. Cell type-specific mechanisms of interleukin-8 induction by dengue virus and differential response to drug treatment. J. Infect. Dis. 2006, 193, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Netsawan, J.; Noisakra, S.; Puttikhunt, C.; Kasinrerk, W.; Wongwiwat, W.; Malasit, P.; Yenchitsomanus, P.; Limjindaporn, T. Nuclear localization of dengue virus capsid protein is required for DAXX interaction and apoptosis. Virus Res. 2010, 147, 275–283. [Google Scholar] [CrossRef]
- Bhuvanakantham, R.; Chong, M.K.; Ng, M.L. Specific interaction of capsid protein and importin-α/β influences West Nile virus production. Biochem. Biophys. Res. Commun. 2009, 389, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Johnston, T.; Hendricks, G.L.; Shen, S.; Chen, R.F.; Kwon, B.; Kelso, M.J.; Kim, W.; Burgwyn Fuchs, B.; Mylonakis, E. Raf-kinase inhibitor GW5074 shows antibacterial activity against methicillin-resistant Staphylococcus aureus and potentiates the activity of gentamicin. Future Med. Chem. 2016, 8, 1941–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Garcia, D.; Barril, X. Molecular simulations with solvent competition quantify water displaceability and provide accurate interaction maps of protein binding sites. J. Med. Chem. 2014, 57, 8530–8539. [Google Scholar] [CrossRef] [PubMed]
- Zachariae, U.; Grubmuller, H. Importin-β: Structural and dynamic determinants of a molecular spring. Structure 2008, 16, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Caly, L.; Wagstaff, K.M.; Jans, D.A. Subcellular trafficking of pathogens: Targeting for therapeutics. Antivir. Res. 2012, 95, 202–206. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Parameters | ||
|---|---|---|
| Proteins | Kd * (nM) | IC50 for GW5074 * (µM) |
| IMPα + IMPβ1 a | 5.4 ± 1.1 (2) b | 10.3 ± 0.8 (2) b |
| IMPα/β1 c | N/A d | 12.5 ± 0.1 (2) b |
| IMPα/β1: DENV NS5 e | 0.8 ± 0.2 (2) b | 6.3 ± 0.9 (2) b |
| EC50 (µM) * | |||
|---|---|---|---|
| Virus | DENV | ZIKV | WNV |
| Plaque assay | 0.8 ± 0.4 (2) a | 0.3 (1) a | 5.2 ± 0.7 (2) a |
| RT-qPCR | 1.4 ± 0.2 (2) a | 0.5 ± 0.1 (2) a | 4.8 ± 3.8 (2) a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.N.Y.; Atkinson, S.C.; Fraser, J.E.; Wang, C.; Maher, B.; Roman, N.; Forwood, J.K.; Wagstaff, K.M.; Borg, N.A.; Jans, D.A. Novel Flavivirus Antiviral That Targets the Host Nuclear Transport Importin α/β1 Heterodimer. Cells 2019, 8, 281. https://doi.org/10.3390/cells8030281
Yang SNY, Atkinson SC, Fraser JE, Wang C, Maher B, Roman N, Forwood JK, Wagstaff KM, Borg NA, Jans DA. Novel Flavivirus Antiviral That Targets the Host Nuclear Transport Importin α/β1 Heterodimer. Cells. 2019; 8(3):281. https://doi.org/10.3390/cells8030281
Chicago/Turabian StyleYang, Sundy N. Y., Sarah C. Atkinson, Johanna E. Fraser, Chunxiao Wang, Belinda Maher, Noelia Roman, Jade K. Forwood, Kylie M. Wagstaff, Natalie A. Borg, and David A. Jans. 2019. "Novel Flavivirus Antiviral That Targets the Host Nuclear Transport Importin α/β1 Heterodimer" Cells 8, no. 3: 281. https://doi.org/10.3390/cells8030281
APA StyleYang, S. N. Y., Atkinson, S. C., Fraser, J. E., Wang, C., Maher, B., Roman, N., Forwood, J. K., Wagstaff, K. M., Borg, N. A., & Jans, D. A. (2019). Novel Flavivirus Antiviral That Targets the Host Nuclear Transport Importin α/β1 Heterodimer. Cells, 8(3), 281. https://doi.org/10.3390/cells8030281