MiR-34b-5p Mediates the Proliferation and Differentiation of Myoblasts by Targeting IGFBP2




Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Cells
2.2. Cell Culture and Transfection
2.3. Plasmid Construction
2.4. Dual-Luciferase Reporter Assay
2.5. RNA Isolation, Complementary DNA (cDNA) Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
2.6. EdU (5-Ethynyl-2′-Deoxyuridine) Assay
2.7. Flow Cytometric Analysis
2.8. Western Blotting
2.9. Immunofluorescence
2.10. Statistical Analysis
3. Results
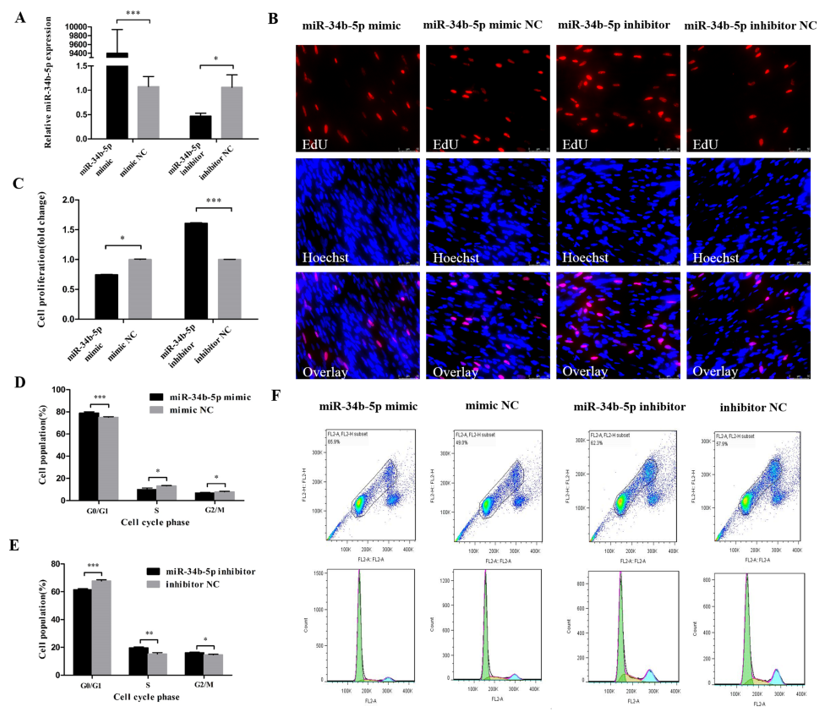
3.1. miR-34b-5p Represses the Cell Cycle Progression of Myoblasts
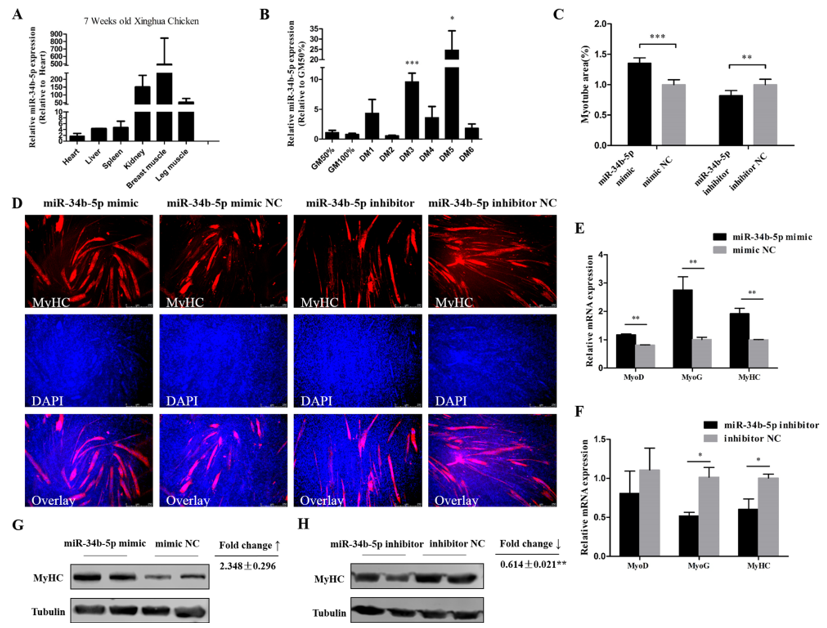
3.2. miR-34b-5p Promotes the Formation of Myotubes
3.3. IGFBP2 is a Direct Target of miR-34b-5p
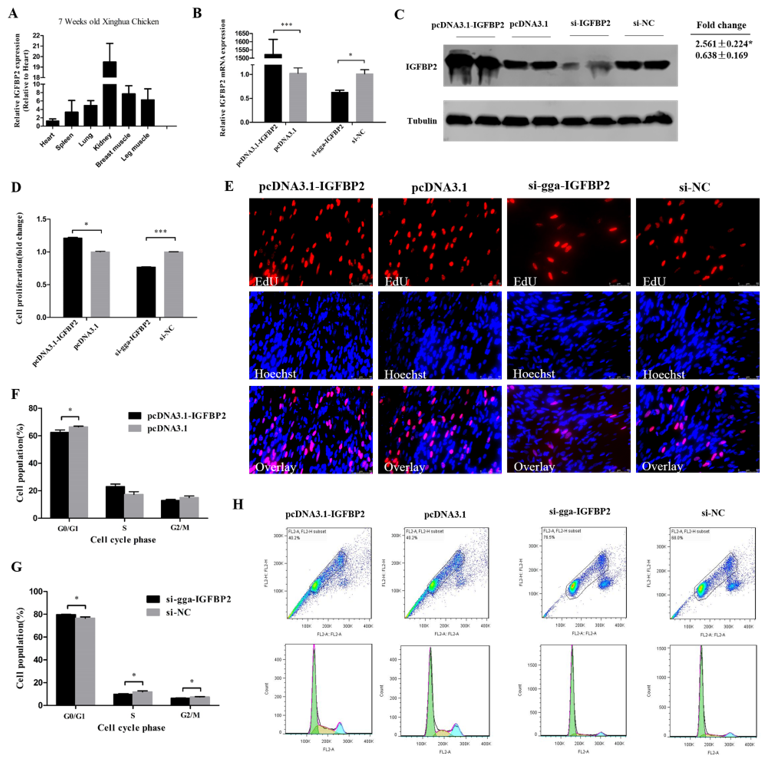
3.4. IGFBP2 Facilitates the Cell Cycle Progression of Myoblasts
3.5. IGFBP2 is an Inhibitor of Myotube Formation
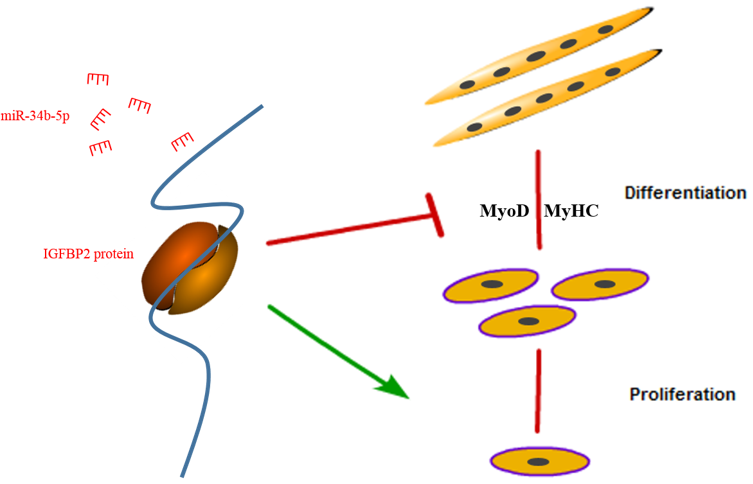
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Saunders, M.A.; Liang, H.; Li, W. Human polymorphism at microRNAs and microRNA target sites. Proc. Natl. Acad. Sci. USA 2007, 104, 3300–3305. [Google Scholar] [CrossRef] [PubMed]
- Cullen, B.R. MicroRNAs as mediators of viral evasion of the immune system. Nat. Immunol. 2013, 14, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Ahmed, R.P.H. MicroRNAs Inducing Proliferation of Quiescent Adult Cardiomyocytes. Cardiovasc. Regener. Med. 2015, 2, e519. [Google Scholar]
- Taft, R.J.; Pang, K.C.; Mercer, T.R.; Dinger, M.; Mattick, J.S. Non-coding RNAs: Regulators of disease. J. Pathol. 2010, 220, 126–139. [Google Scholar] [CrossRef]
- Yu, J.; Peng, J.; Luan, Z.; Zheng, F.; Su, W. MicroRNAs as a Novel Tool in the Diagnosis of Liver Lipid Dysregulation and Fatty Liver Disease. Molecules 2019, 24, 230. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; He, X.; Li, G.; Xu, H.; Jia, X.; Nie, Q.; Zhang, X. Deep Sequencing Analysis of miRNA Expression in Breast Muscle of Fast-Growing and Slow-Growing Broilers. Int. J. Mol. Sci. 2015, 16, 16242–16262. [Google Scholar] [CrossRef] [PubMed]
- Jebessa, E.; Ouyang, H.; Abdalla, B.A.; Li, Z.; Abdullahi, A.Y.; Liu, Q.; Nie, Q.; Zhang, X. Characterization of miRNA and their target gene during chicken embryo skeletal muscle development. Oncotarget 2018, 9, 17309–17324. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, X.; Lim, L.P.; De Stanchina, E.; Xuan, Z.; Liang, Y.; Xue, W.; Zender, L.; Magnus, J.; Ridzon, D.; et al. A microRNA component of the p53 tumour suppressor network. Nature 2007, 447, 1116–1130. [Google Scholar] [CrossRef] [PubMed]
- Bommer, G.T.; Gerin, I.; Feng, Y.; Kaczorowski, A.J.; Kuick, R.; Love, R.E.; Zhai, Y.; Giordano, T.J.; Qin, Z.S.; Moore, B.B.; et al. p53-mediated activation of miRNA34 candidate tumor-suppressor genes. Curr. Biol. 2007, 17, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010, 17, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Luo, Q.; Xu, H.; Zheng, M.; Abdalla, B.A.; Feng, M.; Cai, B.; Zhang, X.; Nie, Q.; Zhang, X. MiR-34b-5p Suppresses Melanoma Differentiation-Associated Gene 5 (MDA5) Signaling Pathway to Promote Avian Leukosis Virus Subgroup J (ALV-J)-Infected Cells Proliferaction and ALV-J Replication. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef]
- Huang, C.W.; Li, Y.H.; Hu, S.Y.; Chi, J.R.; Lin, G.H.; Lin, C.C.; Gong, H.Y.; Chen, J.Y.; Chen, R.H.; Chang, S.J.; et al. Differential expression patterns of growth-related microRNAs in the skeletal muscle of Nile tilapia (Oreochromis niloticus). J. Anim. Sci. 2012, 90, 4266–4279. [Google Scholar] [CrossRef]
- Bae, Y.; Yang, T.; Zeng, H.; Campeau, P.M.; Chen, Y.; Bertin, T.; Dawson, B.C.; Munivez, E.; Tao, J.; Lee, B.H. miRNA-34c regulates Notch signaling during bone development. Hum. Mol. Genet. 2012, 21, 2991–3000. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Shi, Y.; Zheng, L.; Zhou, B.; Inose, H.; Wang, J.; Guo, X.E.; Grosschedl, R.; Karsenty, G. miR-34s inhibit osteoblast proliferation and differentiation in the mouse by targeting SATB2. J. Cell Biol. 2012, 197, 509–521. [Google Scholar] [CrossRef]
- Wang, P.; Xu, J.; Hou, Z.; Wang, F.; Song, Y.; Wang, J.; Zhu, H.; Jin, H. miRNA-34a promotes proliferation of human pulmonary artery smooth muscle cells by targeting PDGFRA. Cell Prolif. 2016, 49, 484–493. [Google Scholar] [CrossRef]
- Chen, Q.; Yang, F.; Guo, M.; Wen, G.; Zhang, C.; Le Anh, L.; Zhu, J.; Xiao, Q.; Zhang, L. miRNA-34a reduces neointima formation through inhibiting smooth muscle cell proliferation and migration. J. Mol. Cell. Cardiol. 2015, 89, 75–86. [Google Scholar] [CrossRef]
- Choe, N.; Kwon, J.; Kim, Y.S.; Eom, G.H.; Ahn, Y.K.; Baik, Y.H.; Park, H.; Kook, H. The microRNA miR-34c inhibits vascular smooth muscle cell proliferation and neointimal hyperplasia by targeting stem cell factor. Cell Signal. 2015, 27, 1056–1065. [Google Scholar] [CrossRef]
- Hou, L.; Xu, J.; Li, H.; Ou, J.; Jiao, Y.; Hu, C.; Wang, C. MiR-34c represses muscle development by forming a regulatory loop with Notch1. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.C. Insulin-like growth factor (IGF)-binding proteins: Interactions with IGFs and intrinsic bioactivities. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E967–E976. [Google Scholar] [CrossRef] [PubMed]
- Pickard, A.; McCance, D.J. IGF-binding protein 2-oncogene or tumor suppressor? Front. Endocrinol. 2015, 6. [Google Scholar] [CrossRef]
- Schoen, T.J.; Mazuruk, K.; Waldbillig, R.J.; Potts, J.; Beebe, D.C.; Chader, G.J.; Rodriguez, I.R. Cloning and characterization of a chick embryo cDNA and gene for IGF-binding protein-2. J. Mol. Endocrinol. 1995, 15, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Gerrard, D.E.; Okamura, C.S.; Grant, A.L. Expression and location of IGF binding proteins-2, -4, and -5 in developing fetal tissues. J. Anim. Sci. 1999, 77, 1431–1441. [Google Scholar] [CrossRef]
- Sironen, A.I.; Uimari, P.; Serenius, T.; Mote, B.; Rothschild, M.; Vilkki, J. Effect of polymorphisms in candidate genes on reproduction traits in Finnish pig populations. J. Anim. Sci. 2010, 88, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Prasongsook, S.; Choi, I.; Bates, R.O.; Raney, N.E.; Ernst, C.W.; Tumwasorn, S. Association of Insulin-like growth factor binding protein 2 genotypes with growth, carcass and meat quality traits in pigs. J. Anim. Sci. Technol. 2015, 57, 31. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ouyang, H.; Chen, X.; Yu, J.; Abdallat, B.A.; Chen BNie, Q. Gga-miR-205a Affecting Myoblast Proliferation and Differentiation by Targeting CDH11. Front. Genet. 2018, 9. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bass, J.J.; Wilkinson, D.J.; Rankin, D.; Phillips, B.E.; Szewczyk, N.J.; Smith KAtherton, P.J. An overview of technical considerations for Western blotting applications to physiological research. Scand. J. Med. Sci. Sports 2017, 27, 4–25. [Google Scholar] [CrossRef]
- Chen, E.H.; Grote, E.; Mohler, W.; Vignery, A. Cell-cell fusion. FEBS Lett. 2007, 581, 2181–2193. [Google Scholar] [CrossRef]
- Luo, W.; Nie, Q.; Zhang, X. MicroRNAs Involved in Skeletal Muscle Differentiation. J. Genet. Genom. 2013, 40, 107–116. [Google Scholar] [CrossRef]
- Rokavec, M.; Li, H.; Jiang, L.; Hermeking, H. The p53/miR-34 axis in development and disease. J. Mol. Cell Biol. 2014, 6, 214–230. [Google Scholar] [CrossRef]
- Tang, Z.; Qiu, H.; Luo, L.; Liu, N.; Zhong, J.; Kang, K.; Gou, D. miR-34b Modulates Skeletal Muscle Cell Proliferation and Differentiation. J. Cell. Biochem. 2017, 118, 4285–4295. [Google Scholar] [CrossRef]
- Venuti, J.M.; Morris, J.H.; Vivian, J.L.; Olson, E.N.; Klein, W.H. Myogenin is required for late but not early aspects of myogenesis during mouse development. J. Cell Biol. 1995, 128, 563–576. [Google Scholar] [CrossRef]
- Rudnicki, M.A.; Schnegelsberg, P.N.; Stead, R.H.; Braun, T.; Arnold, H.H.; Jaenisch, R. MyoD or Myf-5 is required for the formation of skeletal muscle. Cell 1993, 75, 1351–1359. [Google Scholar] [CrossRef]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Pette, D.; Staron, R.S. Myosin isoforms, muscle fiber types, and transitions. Microsc. Res. Tech. 2000, 50, 500–509. [Google Scholar] [CrossRef]
- Azar, W.J.; Zivkovic, S.; Werther, G.A.; Russo, V.C. IGFBP-2 nuclear translocation is mediated by a functional NLS sequence and is essential for its pro-tumorigenic actions in cancer cells. Oncogene 2014, 33, 578–588. [Google Scholar] [CrossRef]
- Lei, M.M.; Nie, Q.H.; Peng, X.; Zhang, D.X.; Zhang, X.Q. Single nucleotide polymorphisms of the chicken insulin-like factor binding protein 2 gene associated with chicken growth and carcass traits. Poultry Sci. 2005, 84, 1191–1198. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Li, H.; Zhang, H.; Wang, S.Z.; Wang, Q.G.; Wang, Y.X. Identification of a single nucleotide polymorphism of the insulin-like growth factor binding protein 2 gene and its association with growth and body composition traits in the chicken. J. Anim. Sci. 2006, 84, 2902–2906. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Luo, W.; Abdalla, B.A.; Ouyang, H.; Yu, J.; Hu, F.; Nie, Q.; Zhang, X. miRNA-223 upregulated by MYOD inhibits myoblast proliferation by repressing IGF2 and facilitates myoblast differentiation by inhibiting ZEB1. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Chen, J.; Li, L.; Ren, X.; Cheng, T.; Lu, S.; Lawal, R.A.; Nie, Q.; Zhang, X.; Hanotte, O. c-Myc inhibits myoblast differentiation and promotes myoblast proliferation and muscle fibre hypertrophy by regulating the expression of its target genes, miRNAs and lincRNAs. Cell Death Differ. 2018. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fragment Name | Sequence (5′ to 3′) |
|---|---|
| miR-34b-5p mimic | CAGGCAGUGUAGUUAGCUGAUUG |
| miR-34b-5p inhibitor | CUUUCUGCUUUCUUCUCUGCCUG |
| si-gga-IGFBP2 | GGGAGTGTCTCTCTTTCTT |
| Primer Name | Primer Sequences (5′ to 3′) | Size (bp) |
|---|---|---|
| IGFBP2-WT | F:ACCATTTCCCTCTTCCTCC | 295 |
| R:ACCAAGCATTCAGCTCCAC | ||
| IGFBP2-MT | F:CGAGACCAAAGACTGTAAATTGTGAGTCTTGTGTCCTGCC | 2987 |
| R:GGCAGGACACAAGACTCACAATTTACAGTCTTTGGTCTCG | ||
| IGFBP2 | F:AGCGGCAGATGGGCAAAGT | 184 |
| R:GGGGATGTGGAGGGAGTAGAGG | ||
| β-actin | F:TCATTGTGCTAGGTGCCA | 160 |
| R:TCATTGTGCTAGGTGCCA | ||
| MyoD | F: GCTACTACACGGAATCACCAAAT | 200 |
| R: CTGGGCTCCACTGTCACTCA | ||
| MyoG | F: CGGAGGCTGAAGAAGGTGAA | 320 |
| R: CGGTCCTCTGCCTGGTCAT | ||
| MyHC | F: CTCCTCACGCTTTGGTAA | 213 |
| R: TGATAGTCGTATGGGTTGGT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhang, X.; Li, Z.; Abdalla, B.A.; Chen, Y.; Nie, Q. MiR-34b-5p Mediates the Proliferation and Differentiation of Myoblasts by Targeting IGFBP2. Cells 2019, 8, 360. https://doi.org/10.3390/cells8040360
Wang Z, Zhang X, Li Z, Abdalla BA, Chen Y, Nie Q. MiR-34b-5p Mediates the Proliferation and Differentiation of Myoblasts by Targeting IGFBP2. Cells. 2019; 8(4):360. https://doi.org/10.3390/cells8040360
Chicago/Turabian StyleWang, Zhijun, Xiaocui Zhang, Zhenhui Li, Bahareldin Ali Abdalla, Yangfeng Chen, and Qinghua Nie. 2019. "MiR-34b-5p Mediates the Proliferation and Differentiation of Myoblasts by Targeting IGFBP2" Cells 8, no. 4: 360. https://doi.org/10.3390/cells8040360
APA StyleWang, Z., Zhang, X., Li, Z., Abdalla, B. A., Chen, Y., & Nie, Q. (2019). MiR-34b-5p Mediates the Proliferation and Differentiation of Myoblasts by Targeting IGFBP2. Cells, 8(4), 360. https://doi.org/10.3390/cells8040360