Administration of Tonsil-Derived Mesenchymal Stem Cells Improves Glucose Tolerance in High Fat Diet-Induced Diabetic Mice via Insulin-Like Growth Factor-Binding Protein 5-Mediated Endoplasmic Reticulum Stress Modulation

 , ,
, ,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Glucose and Insulin Tolerance Testing
2.3. Glucose-Stimulated Insulin Secretion
2.4. Histology and Immunofluorescence
2.5. Cell Culture
2.6. CM Preparation
2.7. MTT
2.8. Real-Time Polymerase Chain Reaction (PCR)
2.9. Western Blot Analyses
2.10. ELISA
2.11. Cholesterol and Triglyceride Measurements
2.12. Fluorescence-Activated Cell Sorting
2.13. Microarray Analysis
2.14. Proteomics
2.15. Statistical Analysis
3. Results
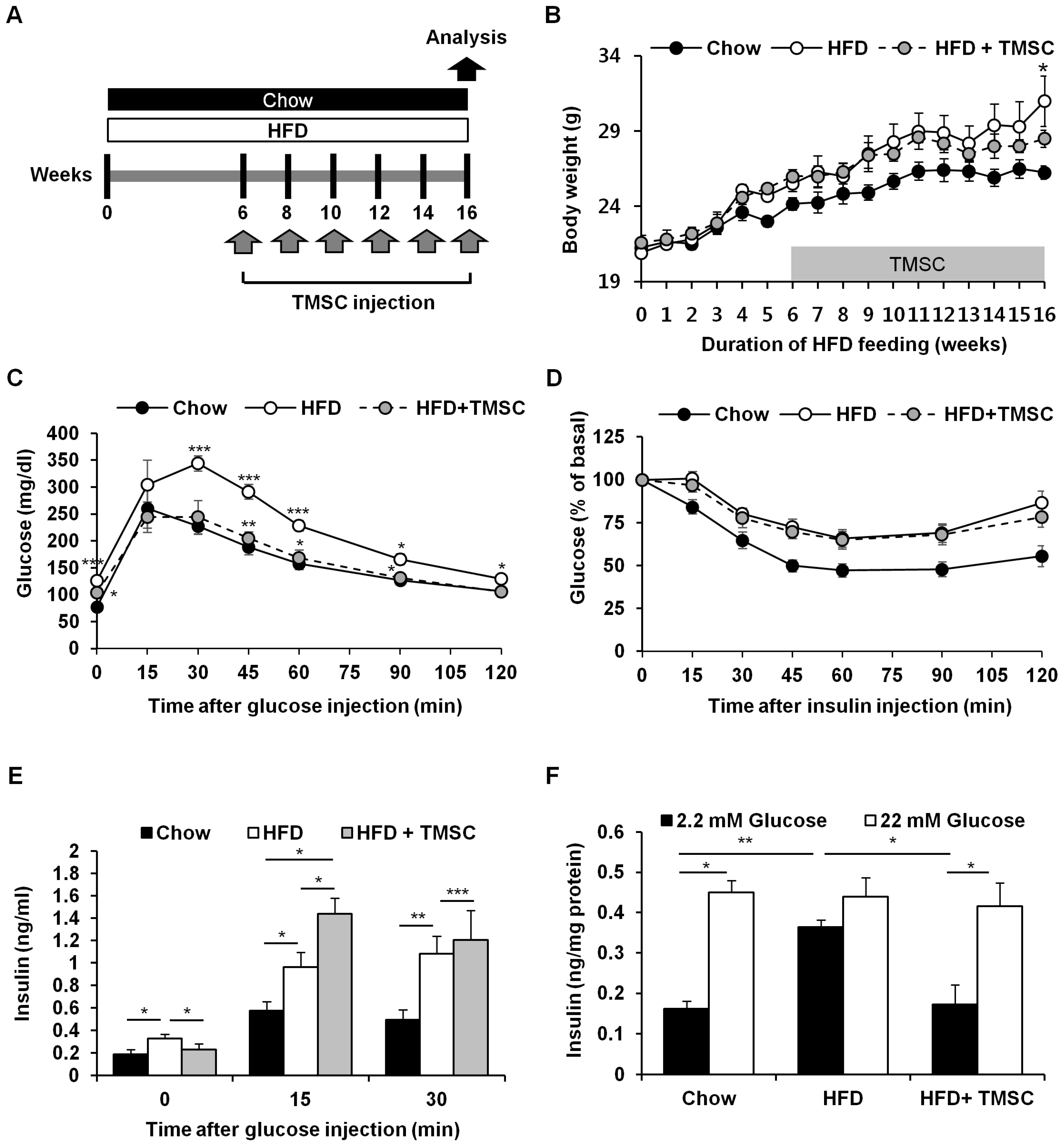
3.1. Intraperitoneal Administration of TMSCs Alleviated HFD-Induced Glucose Intolerance
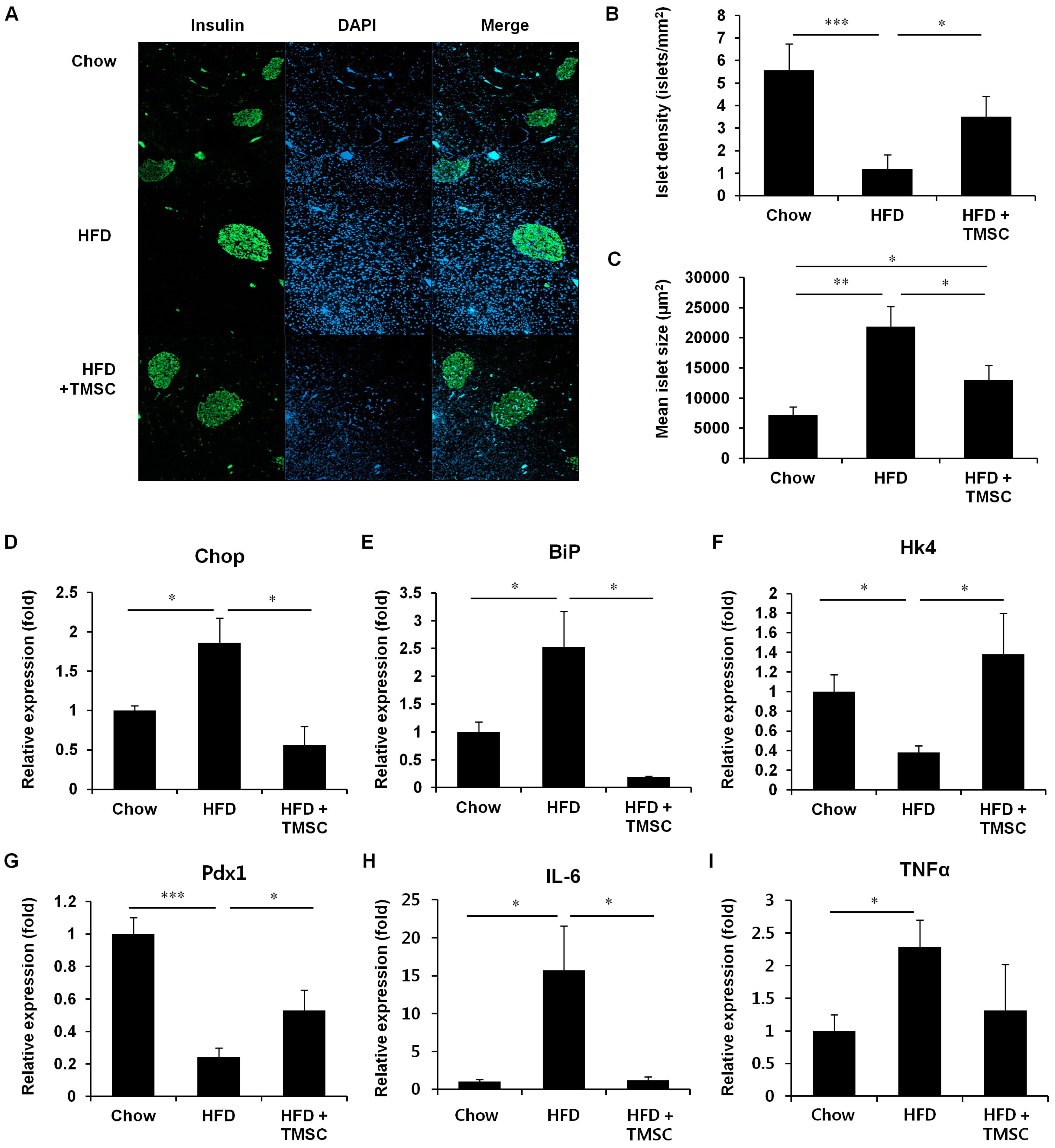
3.2. TMSC Administration Preserved Pancreas Integrity Despite HFD
3.3. CM from TMSCs Also Improved HFD-Induced Glucose Tolerance
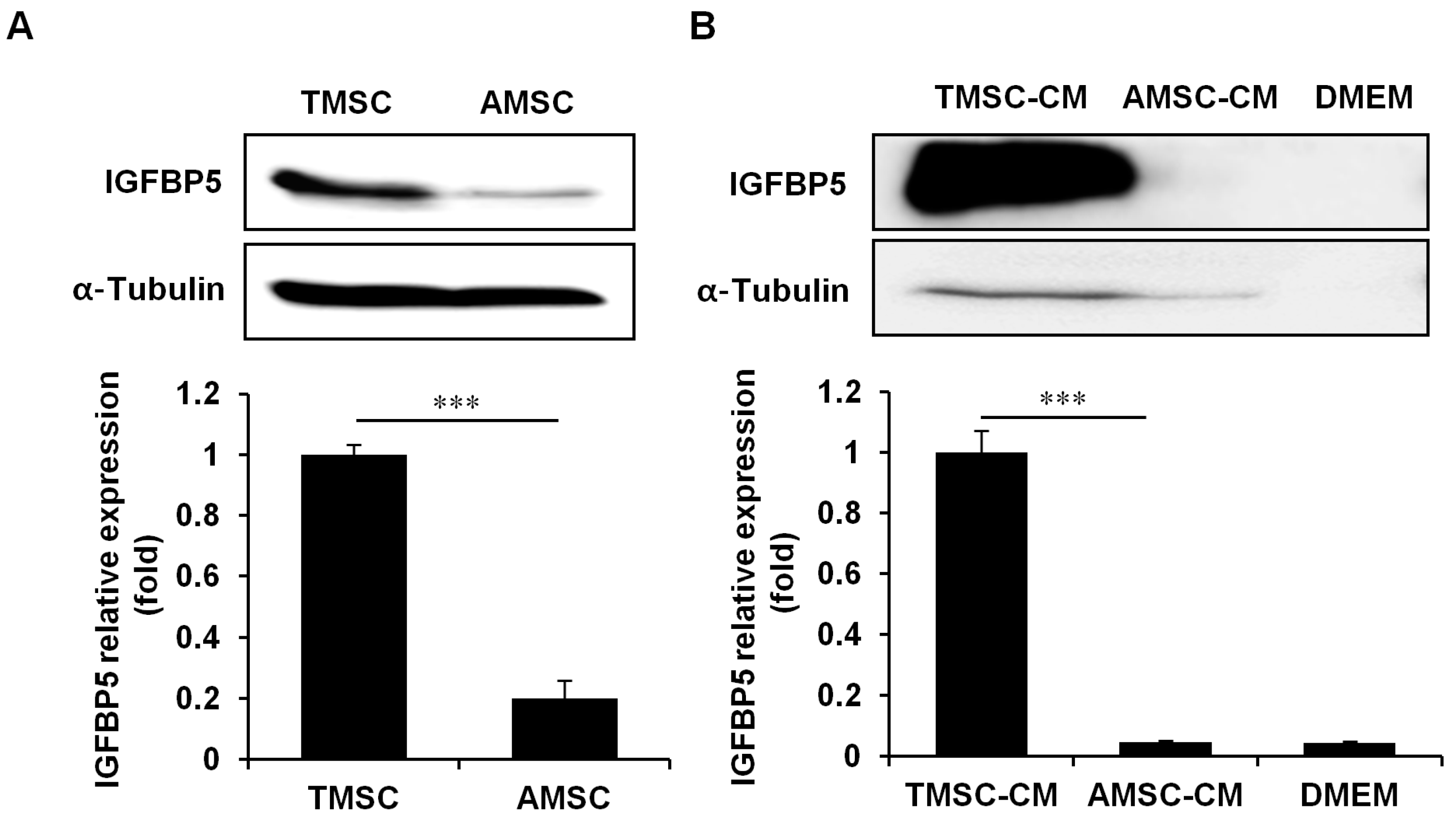
3.4. IGFBP5 was Critical for TMSC Protective Effects Against HFD-Induced Glucose Intolerance
3.5. IGFBP5 Knockdown in TMSCs Diminished Cell Proliferation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMSC | adipose tissue-derived MSC |
| AMSC-CM | conditioned medium derived from AMSCs |
| BiP | binding immunoglobulin protein |
| BM-MSC | bone marrow-derived MSC |
| cDNA | complementary DNA |
| CHOP | C/EBP homologous protein |
| CM | conditioned medium |
| ER | endoplasmic reticulum |
| Glut4 | glucose transporter type 4 |
| HFD | high fat diet |
| Hk4 | hexokinase 4 |
| HKRB | HEPES-added Krebs-Ringer bicarbonate buffer |
| IGF | insulin-like growth factor |
| IGFBP | insulin-like growth factor-binding protein |
| InsR | insulin receptor |
| IL-6 | interleukin-6 |
| MS | mass spectrometry |
| MSC | mesenchymal stem cell |
| PCR | polymerase chain reaction |
| Pdx1 | pancreatic and duodenal homeobox 1 |
| PLA2 | phospholipase A2 |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| STZ | streptozotocin |
| T2DM | type 2 diabetes mellitus |
| TMSC | tonsil-derived mesenchymal stem cell |
| TMSC-CM | conditioned medium derived from TMSCs |
| TNFα | tumor necrosis factor α |
References
- Back, S.H.; Kaufman, R.J. Endoplasmic reticulum stress and type 2 diabetes. Annu. Rev. Biochem. 2012, 81, 767–793. [Google Scholar] [CrossRef]
- Butler, A.E.; Janson, J.; Bonner-Weir, S.; Ritzel, R.; Rizza, R.A.; Butler, P.C. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes 2003, 52, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Prentki, M.; Nolan, C.J. Islet beta cell failure in type 2 diabetes. J. Clin. Invest. 2006, 116, 1802–1812. [Google Scholar] [CrossRef] [PubMed]
- Cnop, M.; Ladriere, L.; Igoillo-Esteve, M.; Moura, R.F.; Cunha, D.A. Causes and cures for endoplasmic reticulum stress in lipotoxic beta-cell dysfunction. Diabetes Obes. Metab. 2010, 12 (Suppl. 2), 76–82. [Google Scholar] [CrossRef]
- Rivera, J.F.; Costes, S.; Gurlo, T.; Glabe, C.G.; Butler, P.C. Autophagy defends pancreatic beta cells from human islet amyloid polypeptide-induced toxicity. J. Clin. Invest. 2014, 124, 3489–3500. [Google Scholar] [CrossRef]
- Zang, L.; Hao, H.; Liu, J.; Li, Y.; Han, W.; Mu, Y. Mesenchymal stem cell therapy in type 2 diabetes mellitus. Diabetol. Metab. Syndr. 2017, 9, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells - current trends and future prospective. Biosci. Rep. 2015, 35. [Google Scholar] [CrossRef] [PubMed]
- Ryu, K.H.; Kim, S.Y.; Kim, Y.R.; Woo, S.Y.; Sung, S.H.; Kim, H.S.; Jung, S.C.; Jo, I.; Park, J.W. Tonsil-derived mesenchymal stem cells alleviate concanavalin A-induced acute liver injury. Exp. Cell Res. 2014, 326, 143–154. [Google Scholar] [CrossRef]
- Park, M.; Kim, Y.H.; Woo, S.Y.; Lee, H.J.; Yu, Y.; Kim, H.S.; Park, Y.S.; Jo, I.; Park, J.W.; Jung, S.C.; et al. Tonsil-derived mesenchymal stem cells ameliorate CCl4-induced liver fibrosis in mice via autophagy activation. Sci. Rep. 2015, 5, 8616. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Park, M.; Cho, K.A.; Kim, B.K.; Ryu, J.H.; Woo, S.Y.; Ryu, K.H. Tonsil-Derived Mesenchymal Stem Cells Promote Bone Mineralization and Reduce Marrow and Visceral Adiposity in a Mouse Model of Senile Osteoporosis. Stem Cells Dev. 2016, 25, 1161–1171. [Google Scholar] [CrossRef]
- Kim, G.; Jin, Y.M.; Yu, Y.; Kim, H.Y.; Jo, S.A.; Park, Y.J.; Park, Y.S.; Jo, I. Double intratibial injection of human tonsil-derived mesenchymal stromal cells recovers postmenopausal osteoporotic bone mass. Cytotherapy 2018, 20, 1013–1027. [Google Scholar] [CrossRef]
- Yu, Y.; Song, E.M.; Lee, K.E.; Joo, Y.H.; Kim, S.E.; Moon, C.M.; Kim, H.Y.; Jung, S.A.; Jo, I. Therapeutic potential of tonsil-derived mesenchymal stem cells in dextran sulfate sodium-induced experimental murine colitis. PLoS ONE 2017, 12, e0183141. [Google Scholar] [CrossRef]
- Baugh, R.F.; Archer, S.M.; Mitchell, R.B.; Rosenfeld, R.M.; Amin, R.; Burns, J.J.; Darrow, D.H.; Giordano, T.; Litman, R.S.; Li, K.K.; et al. Clinical practice guideline: Tonsillectomy in children. Otolaryngol. Head Neck Surg. 2011, 144, S1–S30. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, Y.R.; Park, W.J.; Kim, H.S.; Jung, S.C.; Woo, S.Y.; Jo, I.; Ryu, K.H.; Park, J.W. Characterisation of insulin-producing cells differentiated from tonsil derived mesenchymal stem cells. Differentiation 2015, 90, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.A.; Park, M.; Kim, Y.H.; Woo, S.Y.; Ryu, K.H. RNA sequencing reveals a transcriptomic portrait of human mesenchymal stem cells from bone marrow, adipose tissue, and palatine tonsils. Sci. Rep. 2017, 7, 17114. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Park, Y.S.; Kim, H.S.; Kim, H.Y.; Jin, Y.M.; Jung, S.C.; Ryu, K.H.; Jo, I. Characterization of long-term in vitro culture-related alterations of human tonsil-derived mesenchymal stem cells: Role for CCN1 in replicative senescence-associated increase in osteogenic differentiation. J. Anat. 2014, 225, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Ryu, K.H.; Cho, K.A.; Park, H.S.; Kim, J.Y.; Woo, S.Y.; Jo, I.; Choi, Y.H.; Park, Y.M.; Jung, S.C.; Chung, S.M.; et al. Tonsil-derived mesenchymal stromal cells: Evaluation of biologic, immunologic and genetic factors for successful banking. Cytotherapy 2012, 14, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Kim, H.S.; Jin, Y.M.; Yu, Y.; Kim, H.Y.; Park, H.S.; Jung, S.C.; Han, K.H.; Park, Y.J.; Ryu, K.H.; et al. Differentiated tonsil-derived mesenchymal stem cells embedded in Matrigel restore parathyroid cell functions in rats with parathyroidectomy. Biomaterials 2015, 65, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Choi, Y.; Jung, N.; Yu, Y.; Ryu, K.H.; Kim, H.S.; Jo, I.; Choi, B.O.; Jung, S.C. Myogenic differentiation potential of human tonsil-derived mesenchymal stem cells and their potential for use to promote skeletal muscle regeneration. Int. J. Mol. Med. 2016, 37, 1209–1220. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Lee, S.Y.; Yang, E.J.; Kim, H.Y.; Jo, I.; Shin, S.J. Expression of tenocyte lineage-related factors from tonsil-derived mesenchymal stem cells. Tissue Eng. Regen. Med. 2016, 13, 162–170. [Google Scholar] [CrossRef]
- Patel, M.; Moon, H.J.; Jung, B.K.; Jeong, B. Microsphere-Incorporated Hybrid Thermogel for Neuronal Differentiation of Tonsil Derived Mesenchymal Stem Cells. Adv. Healthc. Mater. 2015, 4, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Zhao, Y.; Hao, H.; Liu, J.; Guo, Y.; Mu, Y.; Shen, J.; Cheng, Y.; Fu, X.; Han, W. Infusion of mesenchymal stem cells ameliorates hyperglycemia in type 2 diabetic rats: Identification of a novel role in improving insulin sensitivity. Diabetes 2012, 61, 1616–1625. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Pan, Q.; Dong, H.; Yuan, X.; Li, Y.; Sun, Z.; Dong, X.; Wang, H. Adipose-derived mesenchymal stem cells improve glucose homeostasis in high-fat diet-induced obese mice. Stem Cell Res. Ther. 2015, 6, 208. [Google Scholar] [CrossRef] [PubMed]
- Sensebe, L.; Fleury-Cappellesso, S. Biodistribution of mesenchymal stem/stromal cells in a preclinical setting. Stem Cells Int. 2013, 2013, 678063. [Google Scholar] [CrossRef]
- Heydemann, A. An Overview of Murine High Fat Diet as a Model for Type 2 Diabetes Mellitus. J. Diabetes Res. 2016, 2016, 2902351. [Google Scholar] [CrossRef]
- Park, J.W.; Park, W.J.; Kuperman, Y.; Boura-Halfon, S.; Pewzner-Jung, Y.; Futerman, A.H. Ablation of very long acyl chain sphingolipids causes hepatic insulin resistance in mice due to altered detergent-resistant membranes. Hepatology 2013, 57, 525–532. [Google Scholar] [CrossRef]
- Li, D.S.; Yuan, Y.H.; Tu, H.J.; Liang, Q.L.; Dai, L.J. A protocol for islet isolation from mouse pancreas. Nature Protocols 2009, 4, 1649–1652. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.A.; Park, M.; Kim, Y.H.; Woo, S.Y.; Ryu, K.H. Conditioned media from human palatine tonsil mesenchymal stem cells regulates the interaction between myotubes and fibroblasts by IL-1Ra activity. J. Cell Mol. Med. 2017, 21, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Cho, K.A.; Park, M.; Kim, Y.H.; Ryu, K.H.; Woo, S.Y. Mesenchymal stem cells inhibit RANK-RANKL interactions between osteoclasts and Th17 cells via osteoprotegerin activity. Oncotarget 2017, 8, 83419–83431. [Google Scholar] [CrossRef] [Green Version]
- Jehle, P.M.; Jehle, D.R.; Mohan, S.; Bohm, B.O. Serum levels of insulin-like growth factor system components and relationship to bone metabolism in Type 1 and Type 2 diabetes mellitus patients. J. Endocrinol. 1998, 159, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jia, Z.; Diao, S.; Lin, X.; Lian, X.; Wang, L.; Dong, R.; Liu, D.; Fan, Z. IGFBP5 enhances osteogenic differentiation potential of periodontal ligament stem cells and Wharton’s jelly umbilical cord stem cells, via the JNK and MEK/Erk signalling pathways. Cell Prolif. 2016, 49, 618–627. [Google Scholar] [CrossRef]
- Han, N.; Zhang, F.; Li, G.; Zhang, X.; Lin, X.; Yang, H.; Wang, L.; Cao, Y.; Du, J.; Fan, Z. Local application of IGFBP5 protein enhanced periodontal tissue regeneration via increasing the migration, cell proliferation and osteo/dentinogenic differentiation of mesenchymal stem cells in an inflammatory niche. Stem Cell Res. Ther. 2017, 8, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Song, L.; Shen, K.; Wang, H.; Qian, M.; Niu, W.; Qin, X. Bone marrow mesenchymal stem cells promote the repair of islets from diabetic mice through paracrine actions. Mol. Cell Endocrinol. 2014, 388, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Zickri, M.B.; Fadl, S.G.; Metwally, H.G. Comparative Study between Intravenous and Intraperitoneal Stem Cell Therapy in Amiodarone Induced Lung Injury in Rat. Int. J. Stem Cells 2014, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Liang, C.; Hu, H.; Zhou, L.; Xu, B.; Wang, X.; Han, Y.; Nie, Y.; Jia, S.; Liang, J.; et al. Intraperitoneal injection (IP), Intravenous injection (IV) or anal injection (AI)? Best way for mesenchymal stem cells transplantation for colitis. Sci. Rep. 2016, 6, 30696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Luca, C.; Olefsky, J.M. Inflammation and insulin resistance. FEBS Lett. 2008, 582, 97–105. [Google Scholar] [CrossRef]
- Park, M.; Kim, Y.H.; Ryu, J.H.; Woo, S.Y.; Ryu, K.H. Immune suppressive effects of tonsil-derived mesenchymal stem cells on mouse bone-marrow-derived dendritic cells. Stem Cells Int. 2015, 2015, 106540. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, M.; Kim, Y.H.; Ryu, K.H.; Lee, K.H.; Cho, K.A.; Woo, S.Y. Tonsil-derived mesenchymal stem cells (T-MSCs) prevent Th17-mediated autoimmune response via regulation of the programmed death-1/programmed death ligand-1 (PD-1/PD-L1) pathway. J. Tissue Eng. Regen. Med. 2018, 12, e1022–e1033. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, H.; Storgaard, H.; Rask-Madsen, C.; Steffen Hermann, T.; Ihlemann, N.; Baunbjerg Nielsen, D.; Spohr, C.; Kober, L.; Vaag, A.; Torp-Pedersen, C. Metabolic and vascular effects of tumor necrosis factor-alpha blockade with etanercept in obese patients with type 2 diabetes. J. Vasc Res. 2005, 42, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Gleason, C.E.; Ning, Y.; Cominski, T.P.; Gupta, R.; Kaestner, K.H.; Pintar, J.E.; Birnbaum, M.J. Role of insulin-like growth factor-binding protein 5 (IGFBP5) in organismal and pancreatic beta-cell growth. Mol. Endocrinol. 2010, 24, 178–192. [Google Scholar] [CrossRef]
- Firth, S.M.; Baxter, R.C. Cellular actions of the insulin-like growth factor binding proteins. Endocr. Rev. 2002, 23, 824–854. [Google Scholar] [CrossRef]
- Sureshbabu, A.; Okajima, H.; Yamanaka, D.; Tonner, E.; Shastri, S.; Maycock, J.; Szymanowska, M.; Shand, J.; Takahashi, S.; Beattie, J.; et al. IGFBP5 induces cell adhesion, increases cell survival and inhibits cell migration in MCF-7 human breast cancer cells. J. Cell Sci. 2012, 125, 1693–1705. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.K.; Haun, R.S. Insulin-like growth factor binding protein-5 influences pancreatic cancer cell growth. World J. Gastroenterol. 2009, 15, 3355–3366. [Google Scholar] [CrossRef] [Green Version]
- Grkovic, S.; O’Reilly, V.C.; Han, S.; Hong, M.; Baxter, R.C.; Firth, S.M. IGFBP-3 binds GRP78, stimulates autophagy and promotes the survival of breast cancer cells exposed to adverse microenvironments. Oncogene 2013, 32, 2412–2420. [Google Scholar] [CrossRef]
- Kim, H.J.; Li, Q.; Song, W.J.; Yang, H.M.; Kim, S.Y.; Park, S.C.; Ahn, J.O.; Youn, H.Y. Fibroblast growth factor-1 as a mediator of paracrine effects of canine adipose tissue-derived mesenchymal stem cells on in vitro-induced insulin resistance models. BMC Vet. Res. 2018, 14, 351. [Google Scholar] [CrossRef]
- Zhang, L.; Li, K.; Liu, X.; Li, D.; Luo, C.; Fu, B.; Cui, S.; Zhu, F.; Zhao, R.C.; Chen, X. Repeated systemic administration of human adipose-derived stem cells attenuates overt diabetic nephropathy in rats. Stem Cells Dev. 2013, 22, 3074–3086. [Google Scholar] [CrossRef]
- Bazhanov, N.; Ylostalo, J.H.; Bartosh, T.J.; Tiblow, A.; Mohammadipoor, A.; Foskett, A.; Prockop, D.J. Intraperitoneally infused human mesenchymal stem cells form aggregates with mouse immune cells and attach to peritoneal organs. Stem Cell Res. Ther. 2016, 7, 27. [Google Scholar] [CrossRef]
- Chen, L.; Tredget, E.E.; Wu, P.Y.; Wu, Y. Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS ONE 2008, 3, e1886. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name |
|---|
| 14-3-3 protein epsilon isoform |
| 71 Kd heat shock cognate protein |
| Beta-2 microglobulin |
| C1 esterase |
| Calcium binding protein Cab45 precursor |
| Cathepsin D |
| Decorin |
| Fibulin-1D |
| Insulin-like growth factor-binding protein 4 |
| Insulin-like growth factor-binding protein 5 |
| Insulin-like growth factor-binding protein 7 |
| Laminin A3 |
| Laminin B2 |
| Nucleobindin |
| Phospholipase A2 |
| Phospholipid transfer protein |
| Preprostromelysin |
| Pro-matrix metalloproteinase-1 |
| Quiescent cell proline dipeptidase |
| Quiescin |
| Ras GTPase-activating-like protein |
| Transketolase |
| Triosephosphate isomerase |
| Gene Number | Gene Symbol | Gene Description | TMSC/AMSC (fold) |
|---|---|---|---|
| NM_001145938 | MMP1 | Matrix metalloproteinase-1 | 70.7 |
| NM_002422 | MMP3 | Preprostromelysin | 6.27 |
| NM_000599 | IGFBP5 | Insulin-like growth factor-binding protein 5 | 2.18 |
| NM_000227 | LAMA3 | Laminin A3 | 2.09 |
| NM_001311193 | PLA2G4A | Phospholipase A2 | 2.07 |
| NM_001996 | FBLN1 | Fibulin 1 | 1.94 |
| NM_001552 | IGFBP4 | Insulin-like growth factor-binding protein 4 | 1.46 |
| NM_001004128 | QSOX1 | Quiescin sulfhydryl oxidase 1 | 1.29 |
| NM_005013 | NUCB2 | Nucleobindin 2 | 1.03 |
| NM_001242920 | PLTP | Phospholipid transfer protein | −1.03 |
| NM_002292 | LAMB2 | Laminin B2 | −1.40 |
| NM_001734 | C1S | C1 esterase | −1.66 |
| NM_001253835 | IGFBP7 | Insulin-like growth factor-binding protein 7 | −1.75 |
| NM_001920 | DCN | Decorin | −2.27 |
| NM_006761 | YWHAE | Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein epsilon | ND 1 |
| NM_006597 | HSPA8 | Heat shock protein family A | ND |
| NM_004048 | B2M | Beta-2 microglobulin | ND |
| NM_016176 | SDF4 | Stromal cell derived factor 4 | ND |
| NM_001909 | CTSD | Cathepsin D | ND |
| NM_013379 | DPP7 | Dipeptidyl peptidase 7 | ND |
| NM_003870 | IQGAP1 | IQ motif containing GTPase activating protein 1 | ND |
| NM_001064 | TKT | Transketolase | ND |
| NM_001258026 | TPI1 | Triosephosphate isomerase 1 | ND |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Shin, S.-H.; Cho, K.-A.; Kim, Y.-H.; Woo, S.-Y.; Kim, H.S.; Jung, S.-C.; Jo, I.; Jun, H.-S.; Park, W.-J.; et al. Administration of Tonsil-Derived Mesenchymal Stem Cells Improves Glucose Tolerance in High Fat Diet-Induced Diabetic Mice via Insulin-Like Growth Factor-Binding Protein 5-Mediated Endoplasmic Reticulum Stress Modulation. Cells 2019, 8, 368. https://doi.org/10.3390/cells8040368
Lee Y, Shin S-H, Cho K-A, Kim Y-H, Woo S-Y, Kim HS, Jung S-C, Jo I, Jun H-S, Park W-J, et al. Administration of Tonsil-Derived Mesenchymal Stem Cells Improves Glucose Tolerance in High Fat Diet-Induced Diabetic Mice via Insulin-Like Growth Factor-Binding Protein 5-Mediated Endoplasmic Reticulum Stress Modulation. Cells. 2019; 8(4):368. https://doi.org/10.3390/cells8040368
Chicago/Turabian StyleLee, Younghay, Sun-Hye Shin, Kyung-Ah Cho, Yu-Hee Kim, So-Youn Woo, Han Su Kim, Sung-Chul Jung, Inho Jo, Hee-Sook Jun, Woo-Jae Park, and et al. 2019. "Administration of Tonsil-Derived Mesenchymal Stem Cells Improves Glucose Tolerance in High Fat Diet-Induced Diabetic Mice via Insulin-Like Growth Factor-Binding Protein 5-Mediated Endoplasmic Reticulum Stress Modulation" Cells 8, no. 4: 368. https://doi.org/10.3390/cells8040368
APA StyleLee, Y., Shin, S. -H., Cho, K. -A., Kim, Y. -H., Woo, S. -Y., Kim, H. S., Jung, S. -C., Jo, I., Jun, H. -S., Park, W. -J., Park, J. -W., & Ryu, K. -H. (2019). Administration of Tonsil-Derived Mesenchymal Stem Cells Improves Glucose Tolerance in High Fat Diet-Induced Diabetic Mice via Insulin-Like Growth Factor-Binding Protein 5-Mediated Endoplasmic Reticulum Stress Modulation. Cells, 8(4), 368. https://doi.org/10.3390/cells8040368