Coordination between Rac1 and Rab Proteins: Functional Implications in Health and Disease


Abstract
:1. Introduction
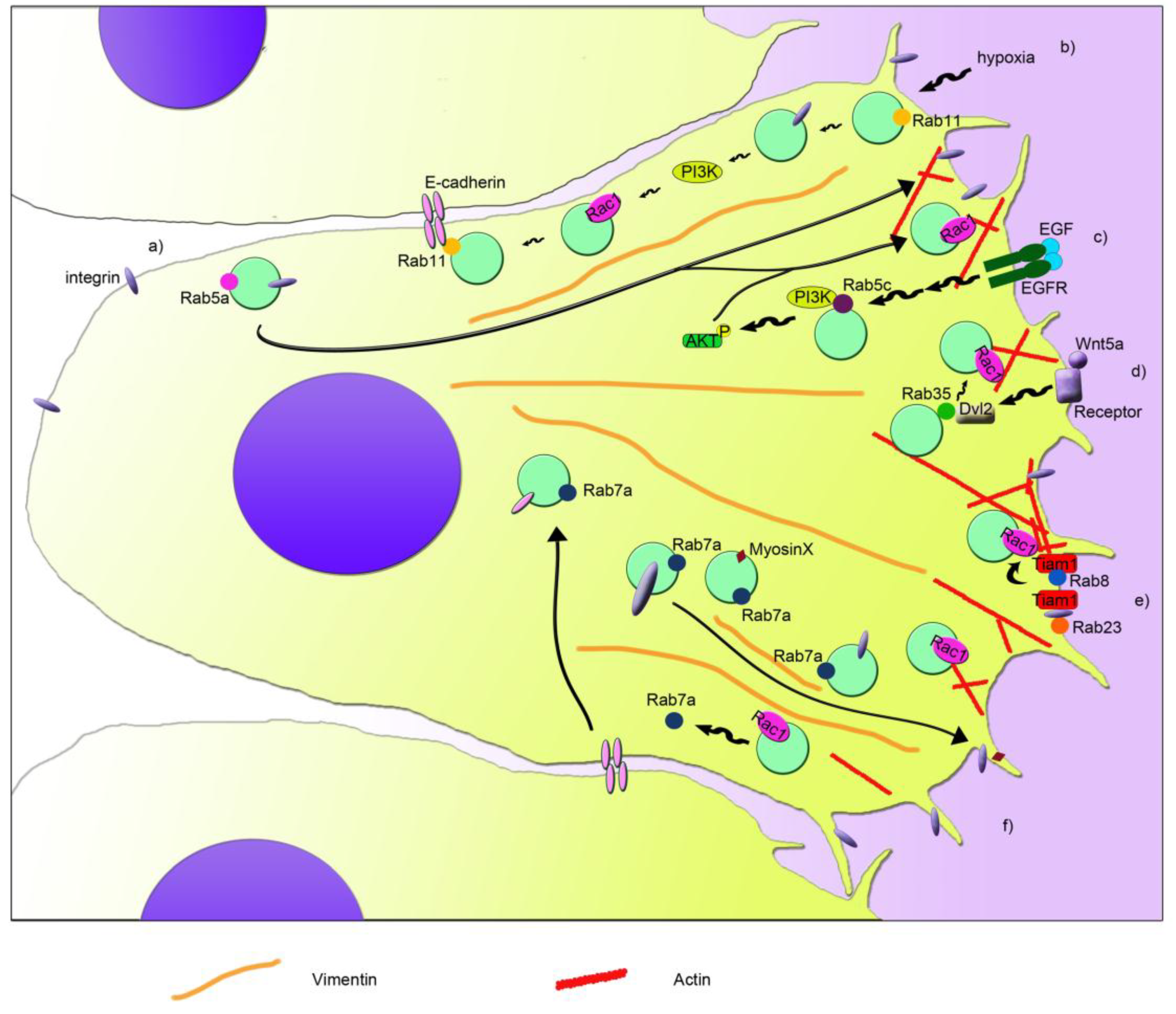
2. Rac1 and Rabs: Interactions and Regulations of Their Functions
3. Rac1 in Cancer: Role of Rac1 and Rabs in Cell Migration and Metastasis
4. Rac1 in Neurological Diseases: Role of Rab Proteins
5. Rac1 and Rab35 in Infections: Regulation of the Innate Immune Response
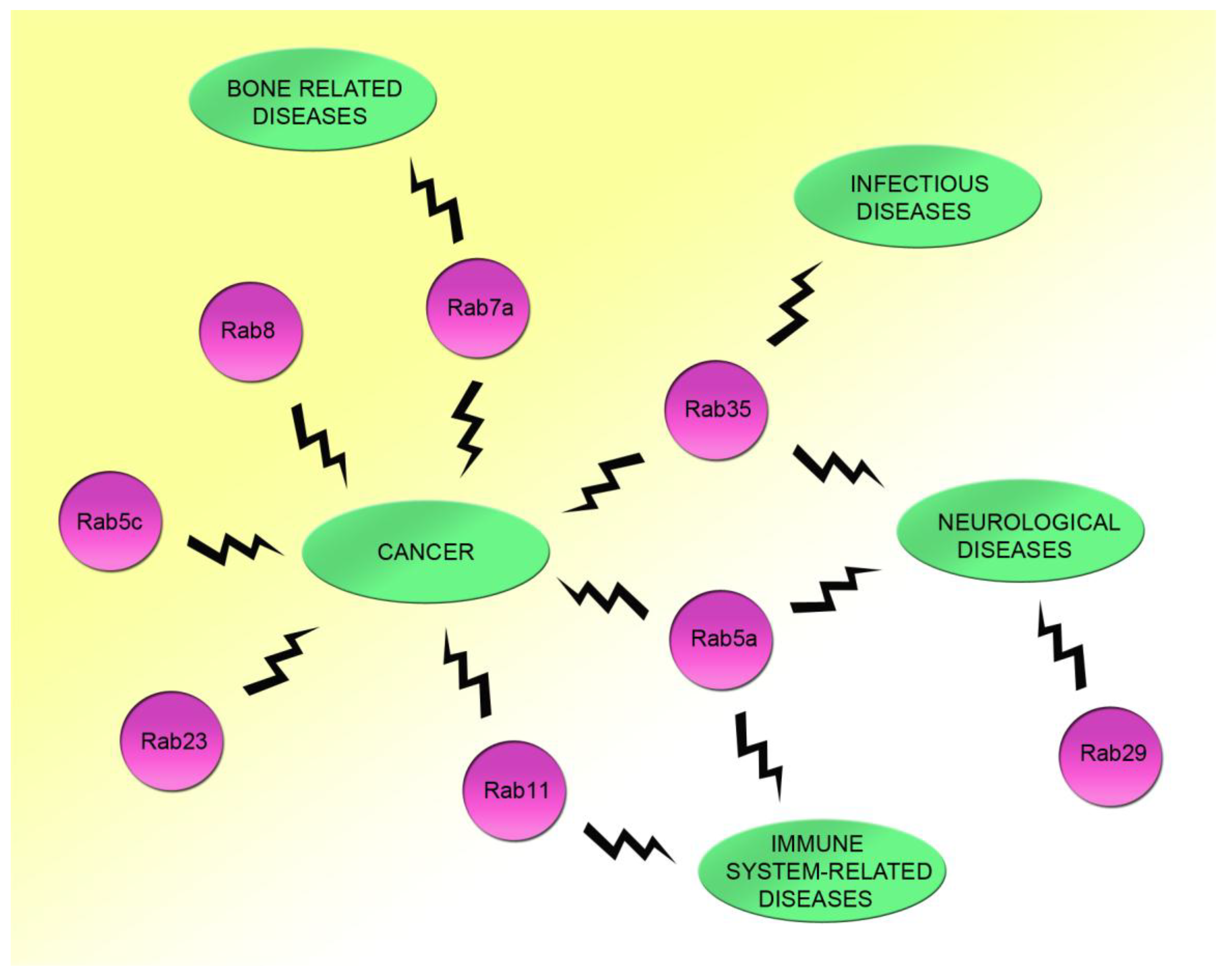
6. Possible Roles of Rac1 and Rab Proteins in Other Diseases
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Haga, R.B.; Ridley, A.J. Rho gtpases: Regulation and roles in cancer cell biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Lérida, I.; Sánchez-Perales, S.; Calvo, M.; Rentero, C.; Zheng, Y.; Enrich, C.; Del Pozo, M.A. A palmitoylation switch mechanism regulates rac1 function and membrane organization. EMBO J. 2012, 31, 534–551. [Google Scholar] [CrossRef] [PubMed]
- Lynch, E.A.; Stall, J.; Schmidt, G.; Chavrier, P.; D'Souza-Schorey, C. Proteasome-mediated degradation of rac1-gtp during epithelial cell scattering. Mol. Biol. Cell 2006, 17, 2236–2242. [Google Scholar] [CrossRef] [PubMed]
- Visvikis, O.; Lorès, P.; Boyer, L.; Chardin, P.; Lemichez, E.; Gacon, G. Activated rac1, but not the tumorigenic variant rac1b, is ubiquitinated on lys 147 through a jnk-regulated process. FEBS J. 2008, 275, 386–396. [Google Scholar] [CrossRef]
- Torrino, S.; Visvikis, O.; Doye, A.; Boyer, L.; Stefani, C.; Munro, P.; Bertoglio, J.; Gacon, G.; Mettouchi, A.; Lemichez, E. The e3 ubiquitin-ligase hace1 catalyzes the ubiquitylation of active rac1. Dev. Cell 2011, 21, 959–965. [Google Scholar] [CrossRef]
- Castillo-Lluva, S.; Tatham, M.H.; Jones, R.C.; Jaffray, E.G.; Edmondson, R.D.; Hay, R.T.; Malliri, A. Sumoylation of the gtpase rac1 is required for optimal cell migration. Nat. Cell Biol. 2010, 12, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.; Kwon, D.Y.; Chun, J.; Kim, J.H.; Kang, S.S. Akt protein kinase inhibits rac1-gtp binding through phosphorylation at serine 71 of rac1. J. Biol. Chem. 2000, 275, 423–428. [Google Scholar] [CrossRef]
- Tong, J.; Li, L.; Ballermann, B.; Wang, Z. Phosphorylation of rac1 t108 by extracellular signal-regulated kinase in response to epidermal growth factor: A novel mechanism to regulate rac1 function. Mol. Cell Biol. 2013, 33, 4538–4551. [Google Scholar] [CrossRef]
- Castillo-Lluva, S.; Tan, C.T.; Daugaard, M.; Sorensen, P.H.; Malliri, A. The tumour suppressor hace1 controls cell migration by regulating rac1 degradation. Oncogene 2013, 32, 1735–1742. [Google Scholar] [CrossRef]
- Schwarz, J.; Proff, J.; Hävemeier, A.; Ladwein, M.; Rottner, K.; Barlag, B.; Pich, A.; Tatge, H.; Just, I.; Gerhard, R. Serine-71 phosphorylation of rac1 modulates downstream signaling. PLoS ONE 2012, 7, e44358. [Google Scholar] [CrossRef]
- Osborn-Heaford, H.L.; Ryan, A.J.; Murthy, S.; Racila, A.M.; He, C.; Sieren, J.C.; Spitz, D.R.; Carter, A.B. Mitochondrial rac1 gtpase import and electron transfer from cytochrome c are required for pulmonary fibrosis. J. Biol. Chem. 2012, 287, 3301–3312. [Google Scholar] [CrossRef] [PubMed]
- Murthy, S.; Ryan, A.; He, C.; Mallampalli, R.K.; Carter, A.B. Rac1-mediated mitochondrial h2o2 generation regulates mmp-9 gene expression in macrophages via inhibition of sp-1 and ap-1. J. Biol. Chem. 2010, 285, 25062–25073. [Google Scholar] [CrossRef] [PubMed]
- Marei, H.; Carpy, A.; Macek, B.; Malliri, A. Proteomic analysis of rac1 signaling regulation by guanine nucleotide exchange factors. Cell Cycle 2016, 15, 1961–1974. [Google Scholar] [CrossRef]
- Quinn, C.C.; Pfeil, D.S.; Wadsworth, W.G. Ced-10/rac1 mediates axon guidance by regulating the asymmetric distribution of mig-10/lamellipodin. Curr. Biol. 2008, 18, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Kärkkäinen, S.; van der Linden, M.; Renkema, G.H. Posh2 is a ring finger e3 ligase with rac1 binding activity through a partial crib domain. FEBS Lett. 2010, 584, 3867–3872. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Mimuro, H.; Miki, H.; Takenawa, T.; Sasaki, T.; Nakanishi, H.; Takai, Y.; Sasakawa, C. Rho family gtpase cdc42 is essential for the actin-based motility of shigella in mammalian cells. J. Exp. Med. 2000, 191, 1905–1920. [Google Scholar] [CrossRef]
- Frasa, M.A.; Maximiano, F.C.; Smolarczyk, K.; Francis, R.E.; Betson, M.E.; Lozano, E.; Goldenring, J.; Seabra, M.C.; Rak, A.; Ahmadian, M.R.; et al. Armus is a rac1 effector that inactivates rab7 and regulates e-cadherin degradation. Curr. Biol. 2010, 20, 198–208. [Google Scholar] [CrossRef]
- Jezyk, M.R.; Snyder, J.T.; Gershberg, S.; Worthylake, D.K.; Harden, T.K.; Sondek, J. Crystal structure of rac1 bound to its effector phospholipase c-beta2. Nat. Struct. Mol. Biol. 2006, 13, 1135–1140. [Google Scholar] [CrossRef]
- Ridley, A.J.; Paterson, H.F.; Johnston, C.L.; Diekmann, D.; Hall, A. The small gtp-binding protein rac regulates growth factor-induced membrane ruffling. Cell 1992, 70, 401–410. [Google Scholar] [CrossRef]
- Guo, F.; Debidda, M.; Yang, L.; Williams, D.A.; Zheng, Y. Genetic deletion of rac1 gtpase reveals its critical role in actin stress fiber formation and focal adhesion complex assembly. J. Biol. Chem. 2006, 281, 18652–18659. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.Y.; Lee, S.; Briscoe, C.; Ellsworth, C.; Firtel, R.A. Role of rac in controlling the actin cytoskeleton and chemotaxis in motile cells. Proc. Natl. Acad. Sci. USA 2000, 97, 5225–5230. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Lal, H.; Golden, H.B.; Gerilechaogetu, F.; Smith, M.; Guleria, R.S.; Foster, D.M.; Lu, G.; Dostal, D.E. Rac1 and rhoa differentially regulate angiotensinogen gene expression in stretched cardiac fibroblasts. Cardiovasc. Res. 2011, 90, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.S.; Wynne, J.; Treisman, R. The rho family gtpases rhoa, rac1, and cdc42hs regulate transcriptional activation by srf. Cell 1995, 81, 1159–1170. [Google Scholar] [CrossRef]
- Westwick, J.K.; Lambert, Q.T.; Clark, G.J.; Symons, M.; Van Aelst, L.; Pestell, R.G.; Der, C.J. Rac regulation of transformation, gene expression, and actin organization by multiple, pak-independent pathways. Mol. Cell Biol. 1997, 17, 1324–1335. [Google Scholar] [CrossRef] [PubMed]
- Giehl, K.; Keller, C.; Muehlich, S.; Goppelt-Struebe, M. Actin-mediated gene expression depends on rhoa and rac1 signaling in proximal tubular epithelial cells. PLoS ONE 2015, 10, e0121589. [Google Scholar] [CrossRef]
- Santibáñez, J.F.; Kocić, J.; Fabra, A.; Cano, A.; Quintanilla, M. Rac1 modulates tgf-beta1-mediated epithelial cell plasticity and mmp9 production in transformed keratinocytes. FEBS Lett. 2010, 584, 2305–2310. [Google Scholar]
- Salvatierra, E.; Alvarez, M.J.; Leishman, C.C.; Rivas Baquero, E.; Lutzky, V.P.; Chuluyan, H.E.; Podhajcer, O.L. Sparc controls melanoma cell plasticity through rac1. PLoS ONE 2015, 10, e0134714. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, L.; Irani, K.; Moldovan, N.I.; Finkel, T.; Goldschmidt-Clermont, P.J. The actin cytoskeleton reorganization induced by rac1 requires the production of superoxide. Antioxid. Redox Sign. 1999, 1, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Zeng, L.W.; Feng, L.; Chen, D.B. Rac1-dependent intracellular superoxide formation mediates vascular endothelial growth factor-induced placental angiogenesis in vitro. Endocrinology 2010, 151, 5315–5325. [Google Scholar] [CrossRef]
- Joneson, T.; Bar-Sagi, D. A rac1 effector site controlling mitogenesis through superoxide production. J. Biol. Chem. 1998, 273, 17991–17994. [Google Scholar] [CrossRef] [PubMed]
- Doanes, A.M.; Irani, K.; Goldschmidt-Clermont, P.J.; Finkel, T. A requirement for rac1 in the pdgf-stimulated migration of fibroblasts and vascular smooth cells. BioChem. Mol. Biol. Int. 1998, 45, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Keely, P.J.; Westwick, J.K.; Whitehead, I.P.; Der, C.J.; Parise, L.V. Cdc42 and rac1 induce integrin-mediated cell motility and invasiveness through pi(3)k. Nature 1997, 390, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, K.; Sasaki, T.; Kotani, H.; Nishioka, H.; Takai, Y. Regulation of cell-cell adhesion by rac and rho small g proteins in mdck cells. J. Cell Biol. 1997, 139, 1047–1059. [Google Scholar] [CrossRef] [PubMed]
- Dumontier, M.; Höcht, P.; Mintert, U.; Faix, J. Rac1 gtpases control filopodia formation, cell motility, endocytosis, cytokinesis and development in dictyostelium. J. Cell Sci. 2000, 113, 2253–2265. [Google Scholar] [PubMed]
- Moore, K.A.; Sethi, R.; Doanes, A.M.; Johnson, T.M.; Pracyk, J.B.; Kirby, M.; Irani, K.; Goldschmidt-Clermont, P.J.; Finkel, T. Rac1 is required for cell proliferation and g2/m progression. Biochem. J. 1997, 326, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.G.; Cho, H.J.; Chang, I.Y.; Yoon, S.P.; Jeon, Y.J.; Chung, M.H.; You, H.J. Rac1 prevents cisplatin-induced apoptosis through down-regulation of p38 activation in nih3t3 cells. FEBS Lett. 2002, 518, 129–134. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, Y.; Shacter, E. Rac1 inhibits apoptosis in human lymphoma cells by stimulating bad phosphorylation on ser-75. Mol. Cell Biol. 2004, 24, 6205–6214. [Google Scholar] [CrossRef] [PubMed]
- Muise, A.M.; Walters, T.; Xu, W.; Shen-Tu, G.; Guo, C.H.; Fattouh, R.; Lam, G.Y.; Wolters, V.M.; Bennitz, J.; van Limbergen, J.; et al. Single nucleotide polymorphisms that increase expression of the guanosine triphosphatase rac1 are associated with ulcerative colitis. Gastroenterology 2011, 141, 633–641. [Google Scholar] [CrossRef]
- Nagase, M.; Kurihara, H.; Aiba, A.; Young, M.J.; Sakai, T. Deletion of rac1gtpase in the myeloid lineage protects against inflammation-mediated kidney injury in mice. PLoS ONE 2016, 11, e0150886. [Google Scholar] [CrossRef]
- Cuadrado, A.; Martín-Moldes, Z.; Ye, J.; Lastres-Becker, I. Transcription factors nrf2 and nf-κb are coordinated effectors of the rho family, gtp-binding protein rac1 during inflammation. J. Biol. Chem. 2014, 289, 15244–15258. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Makino, T.; Davidson, M.; Starcevic, D.; Kislat, A.; Nguyen, N.T.; Hashimoto, T.; Homey, B.; Khavari, P.A.; Bradley, M.; et al. Rac1 activation drives pathologic interactions between the epidermis and immune cells. J. Clin. Invest. 2016, 126, 2661–2677. [Google Scholar]
- Heid, I.; Lubeseder-Martellato, C.; Sipos, B.; Mazur, P.K.; Lesina, M.; Schmid, R.M.; Siveke, J.T. Early requirement of rac1 in a mouse model of pancreatic cancer. Gastroenterology 2011, 141, 719–730. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, L.; He, Y.; Xu, X.; Li, K.; Ma, Y.; Gao, Y.; Wei, D.; Wei, L. Knockdown of rac1 and vasp gene expression inhibits breast cancer cell migration. Oncol Lett. 2018, 16, 2151–2160. [Google Scholar] [CrossRef]
- Yoon, C.; Cho, S.J.; Chang, K.K.; Park, D.J.; Ryeom, S.W.; Yoon, S.S. Role of rac1 pathway in epithelial-to-mesenchymal transition and cancer stem-like cell phenotypes in gastric adenocarcinoma. Mol. Cancer Res. 2017, 15, 1106–1116. [Google Scholar] [CrossRef]
- Ferri, N.; Contini, A.; Bernini, S.K.; Corsini, A. Role of small gtpase protein rac1 in cardiovascular diseases: Development of new selective pharmacological inhibitors. J. Cardiovasc. Pharmacol. 2013, 62, 425–435. [Google Scholar] [CrossRef]
- Zhu, S.; Dai, J.; Liu, H.; Cong, X.; Chen, Y.; Wu, Y.; Hu, H.; Heng, B.C.; Ouyang, H.W.; Zhou, Y. Down-regulation of rac gtpase-activating protein ocrl1 causes aberrant activation of rac1 in osteoarthritis development. Arthritis. Rheumatol. 2015, 67, 2154–2163. [Google Scholar] [CrossRef]
- Shibata, S.; Mu, S.; Kawarazaki, H.; Muraoka, K.; Ishizawa, K.; Yoshida, S.; Kawarazaki, W.; Takeuchi, M.; Ayuzawa, N.; Miyoshi, J.; et al. Rac1 gtpase in rodent kidneys is essential for salt-sensitive hypertension via a mineralocorticoid receptor-dependent pathway. J. Clin. Invest. 2011, 121, 3233–3243. [Google Scholar] [CrossRef]
- Shibata, S.; Nagase, M.; Yoshida, S.; Kawarazaki, W.; Kurihara, H.; Tanaka, H.; Miyoshi, J.; Takai, Y.; Fujita, T. Modification of mineralocorticoid receptor function by rac1 gtpase: Implication in proteinuric kidney disease. Nat. Med. 2008, 14, 1370–1376. [Google Scholar] [CrossRef]
- Kawarazaki, W.; Nagase, M.; Yoshida, S.; Takeuchi, M.; Ishizawa, K.; Ayuzawa, N.; Ueda, K.; Fujita, T. Angiotensin ii- and salt-induced kidney injury through rac1-mediated mineralocorticoid receptor activation. J. Am. Soc. Nephrol. 2012, 23, 997–1007. [Google Scholar] [CrossRef]
- Lim, J.S.; Shin, M.; Kim, H.J.; Kim, K.S.; Choy, H.E.; Cho, K.A. Caveolin-1 mediates salmonella invasion via the regulation of sope-dependent rac1 activation and actin reorganization. J. Infect. Dis. 2014, 210, 793–802. [Google Scholar] [CrossRef]
- Criss, A.K.; Ahlgren, D.M.; Jou, T.S.; McCormick, B.A.; Casanova, J.E. The gtpase rac1 selectively regulates salmonella invasion at the apical plasma membrane of polarized epithelial cells. J. Cell Sci. 2001, 114, 1331–1341. [Google Scholar]
- Stankiewicz, T.R.; Linseman, D.A. Rho family gtpases: Key players in neuronal development, neuronal survival, and neurodegeneration. Front Cell Neurosci. 2014, 8, 314. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, N. Rab and rho gtpases regulate intestinal crypt cell homeostasis and enterocyte function. Small GTPases 2016, 7, 59–64. [Google Scholar] [CrossRef]
- Imamura, H.; Takaishi, K.; Nakano, K.; Kodama, A.; Oishi, H.; Shiozaki, H.; Monden, M.; Sasaki, T.; Takai, Y. Rho and rab small g proteins coordinately reorganize stress fibers and focal adhesions in mdck cells. Mol. Biol. Cell 1998, 9, 2561–2575. [Google Scholar] [CrossRef]
- Sun, Y.; Buki, K.G.; Ettala, O.; Vaaraniemi, J.P.; Vaananen, H.K. Possible role of direct rac1-rab7 interaction in ruffled border formation of osteoclasts. J. Biol. Chem. 2005, 280, 32356–32361. [Google Scholar] [CrossRef]
- Margiotta, A.; Progida, C.; Bakke, O.; Bucci, C. Rab7a regulates cell migration through rac1 and vimentin. Biochim. Biophys. Acta 2017, 1864, 367–381. [Google Scholar] [CrossRef]
- Mascia, A.; Gentile, F.; Izzo, A.; Mollo, N.; De Luca, M.; Bucci, C.; Nitsch, L.; Calì, G. Rab7 regulates cdh1 endocytosis, circular dorsal ruffles genesis and thyroglobulin internalization in a thyroid cell line. J. Cell Physiol. 2016, 231, 1695–1708. [Google Scholar] [CrossRef]
- Jian, Q.; Miao, Y.; Tang, L.; Huang, M.; Yang, Y.; Ba, W.; Liu, Y.; Chi, S.; Li, C. Rab23 promotes squamous cell carcinoma cell migration and invasion via integrin beta1/rac1 pathway. Oncotarget 2016, 7, 5342–5352. [Google Scholar] [CrossRef]
- Kunita, R.; Otomo, A.; Mizumura, H.; Suzuki-Utsunomiya, K.; Hadano, S.; Ikeda, J.E. The rab5 activator als2/alsin acts as a novel rac1 effector through rac1-activated endocytosis. J. Biol. Chem. 2007, 282, 16599–16611. [Google Scholar] [CrossRef]
- Morrison Joly, M.; Williams, M.M.; Hicks, D.J.; Jones, B.; Sanchez, V.; Young, C.D.; Sarbassov, D.D.; Muller, W.J.; Brantley-Sieders, D.; Cook, R.S. Two distinct mtorc2-dependent pathways converge on rac1 to drive breast cancer metastasis. Breast Cancer Res. 2017, 19, 74. [Google Scholar] [CrossRef]
- Hong, M.; Zhang, Z.; Chen, Q.; Lu, Y.; Zhang, J.; Lin, C.; Zhang, F.; Zhang, W.; Li, X.; Zhang, W.; et al. Irf1 inhibits the proliferation and metastasis of colorectal cancer by suppressing the ras-rac1 pathway. Cancer Manag. Res. 2019, 11, 369–378. [Google Scholar] [CrossRef]
- Peng, J.X.; Liang, S.Y.; Li, L. Sfrp1 exerts effects on gastric cancer cells through gsk3beta/rac1mediated restraint of tgfbeta/smad3 signaling. Oncol. Rep. 2019, 41, 224–234. [Google Scholar] [PubMed]
- Caggia, S.; Chunduri, H.; Millena, A.C.; Perkins, J.N.; Venugopal, S.V.; Vo, B.T.; Li, C.; Tu, Y.; Khan, S.A. Novel role of gialpha2 in cell migration: Downstream of pi3-kinase-akt and rac1 in prostate cancer cells. J. Cell Physiol. 2018, 234, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Meng, Y.; Zeng, J.; Zeng, B.; Jiang, X. Ubiquitin e3 ligase march7 promotes proliferation and invasion of cervical cancer cells through vav2-rac1-cdc42 pathway. Oncol. Lett. 2018, 16, 2312–2318. [Google Scholar] [CrossRef]
- Zhou, K.; Rao, J.; Zhou, Z.H.; Yao, X.H.; Wu, F.; Yang, J.; Yang, L.; Zhang, X.; Cui, Y.H.; Bian, X.W.; et al. Rac1-gtp promotes epithelial-mesenchymal transition and invasion of colorectal cancer by activation of stat3. Lab. Invest. 2018, 98, 989–998. [Google Scholar] [CrossRef]
- Jamieson, C.; Lui, C.; Brocardo, M.G.; Martino-Echarri, E.; Henderson, B.R. Rac1 augments wnt signaling by stimulating beta-catenin-lymphoid enhancer factor-1 complex assembly independent of beta-catenin nuclear import. J. Cell Sci. 2015, 128, 3933–3946. [Google Scholar] [CrossRef]
- Bopp, A.; Wartlick, F.; Henninger, C.; Schwarz, M.; Kaina, B.; Fritz, G. Rac1 promotes diethylnitrosamine (den)-induced formation of liver tumors. Carcinogenesis 2015, 36, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Xue, Y.; Liu, W.; Yue, C.; Bi, F.; Xu, J.; Zhang, J.; Li, Y.; Zhong, C.; Chen, Y. Role of activated rac1/cdc42 in mediating endothelial cell proliferation and tumor angiogenesis in breast cancer. PLoS ONE 2013, 8, e66275. [Google Scholar]
- Liang, Y.; Wang, S.; Zhang, Y. Downregulation of dock1 and elmo1 suppresses the migration and invasion of triple-negative breast cancer epithelial cells through the rhoa/rac1 pathway. Oncol. Lett. 2018, 16, 3481–3488. [Google Scholar] [CrossRef] [PubMed]
- Recouvreux, M.V.; Commisso, C. Macropinocytosis: A metabolic adaptation to nutrient stress in cancer. Front Endocrinol. (Lausanne) 2017, 8, 261. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, T.N.; Huysentruyt, L.C. On the origin of cancer metastasis. Crit. Rev. Oncog. 2013, 18, 43–73. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Manzanares, M.; Webb, D.J.; Horwitz, A.R. Cell migration at a glance. J. Cell Sci 2005, 118, 4917–4919. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.S.; Chen, X.M.; Zheng, H.X.; Shi, S.L.; Li, Y. Knockdown of rab5a expression decreases cancer cell motility and invasion through integrin-mediated signaling pathway. J. Biomed. Sci. 2011, 18, 58. [Google Scholar] [CrossRef]
- Chen, P.I.; Schauer, K.; Kong, C.; Harding, A.R.; Goud, B.; Stahl, P.D. Rab5 isoforms orchestrate a "division of labor" in the endocytic network; rab5c modulates rac-mediated cell motility. PLoS ONE 2014, 9, e90384. [Google Scholar] [CrossRef]
- Bravo-Cordero, J.J.; Cordani, M.; Soriano, S.F.; Diez, B.; Munoz-Agudo, C.; Casanova-Acebes, M.; Boullosa, C.; Guadamillas, M.C.; Ezkurdia, I.; Gonzalez-Pisano, D.; et al. A novel high-content analysis tool reveals rab8-driven cytoskeletal reorganization through rho gtpases, calpain and mt1-mmp. J. Cell Sci. 2016, 129, 1734–1749. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Wei, W.C.; Hung, C.N.; Kuo, J.F.; Hsu, C.P.; Chang, K.J.; Chao, W.T. Rab11 collaborates e-cadherin to promote collective cell migration and indicates a poor prognosis in colorectal carcinoma. Eur J. Clin. Invest. 2016, 46, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yuan, Y.; Wu, W.; Zhou, M.; Jiang, Q.; Niu, L.; Ji, J.; Liu, N.; Zhang, L.; Wang, X. Hypoxia stimulates invasion and migration of human cervical cancer cell lines hela/siha through the rab11 trafficking of integrin alphavbeta3/fak/pi3k pathway-mediated rac1 activation. J. Biosci. 2017, 42, 491–499. [Google Scholar] [CrossRef]
- Wang, M.; Dong, Q.; Wang, Y. Rab23 is overexpressed in human astrocytoma and promotes cell migration and invasion through regulation of rac1. Tumour Biol. 2016, 37, 11049–11055. [Google Scholar] [CrossRef]
- Zhu, Y.; Shen, T.; Liu, J.; Zheng, J.; Zhang, Y.; Xu, R.; Sun, C.; Du, J.; Chen, Y.; Gu, L. Rab35 is required for wnt5a/dvl2-induced rac1 activation and cell migration in mcf-7 breast cancer cells. Cell Signal 2013, 25, 1075–1085. [Google Scholar] [CrossRef]
- Bucci, C.; Parton, R.G.; Mather, I.H.; Stunnenberg, H.; Simons, K.; Hoflack, B.; Zerial, M. The small gtpase rab5 functions as a regulatory factor in the early endocytic pathway. Cell 1992, 70, 715–728. [Google Scholar] [CrossRef]
- Bucci, C.; Wandinger-Ness, A.; Lutcke, A.; Chiariello, M.; Bruni, C.; Zerial, M. Rab5a is a common component of the apical and basolateral endocytic machinery in polarized epithelial cells. Proc. Natl. Acad. Sci. USA. 1994, 91, 5061–5065. [Google Scholar] [CrossRef]
- Sandri, C.; Caccavari, F.; Valdembri, D.; Camillo, C.; Veltel, S.; Santambrogio, M.; Lanzetti, L.; Bussolino, F.; Ivaska, J.; Serini, G. The r-ras/rin2/rab5 complex controls endothelial cell adhesion and morphogenesis via active integrin endocytosis and rac signaling. Cell Res. 2012, 22, 1479–1501. [Google Scholar] [CrossRef]
- Pfeffer, S.R. Rab gtpase regulation of membrane identity. Curr. Opin. Cell Biol. 2013, 25, 414–419. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, L.; Wang, S.; Wang, T. Rab7: Roles in membrane trafficking and disease. Biosci. Rep. 2009, 29, 193–209. [Google Scholar] [CrossRef]
- Guerra, F.; Bucci, C. Multiple roles of the small gtpase rab7. Cells 2016, 5, 34. [Google Scholar] [CrossRef]
- Zhou, Y.; Toth, M.; Hamman, M.S.; Monahan, S.J.; Lodge, P.A.; Boynton, A.L.; Salgaller, M.L. Serological cloning of paris-1: A new tbc domain-containing, immunogenic tumor antigen from a prostate cancer cell line. Biochem. Biophys. Res. Commun. 2002, 290, 830–838. [Google Scholar] [CrossRef]
- Croizet-Berger, K.; Daumerie, C.; Couvreur, M.; Courtoy, P.J.; van den Hove, M.F. The endocytic catalysts, rab5a and rab7, are tandem regulators of thyroid hormone production. Proc. Natl. Acad. Sci. USA 2002, 99, 8277–8282. [Google Scholar] [CrossRef]
- Cantalupo, G.; Alifano, P.; Roberti, V.; Bruni, C.B.; Bucci, C. Rab-interacting lysosomal protein (rilp): The rab7 effector required for transport to lysosomes. EMBO J. 2001, 20, 683–693. [Google Scholar] [CrossRef]
- Margiotta, A.; Progida, C.; Bakke, O.; Bucci, C. Characterization of the role of rilp in cell migration. Eur. J. Histochem. 2017, 61, 2783. [Google Scholar] [CrossRef]
- Huber, L.A.; Pimplikar, S.; Parton, R.G.; Virta, H.; Zerial, M.; Simons, K. Rab8, a small gtpase involved in vesicular traffic between the tgn and the basolateral plasma membrane. J. Cell Biol. 1993, 123, 35–45. [Google Scholar] [CrossRef]
- Hattula, K.; Furuhjelm, J.; Arffman, A.; Peranen, J. A rab8-specific gdp/gtp exchange factor is involved in actin remodeling and polarized membrane transport. Mol. Biol. Cell 2002, 13, 3268–3280. [Google Scholar] [CrossRef] [PubMed]
- Peranen, J.; Auvinen, P.; Virta, H.; Wepf, R.; Simons, K. Rab8 promotes polarized membrane transport through reorganization of actin and microtubules in fibroblasts. J. Cell Biol. 1996, 135, 153–167. [Google Scholar] [CrossRef]
- Palamidessi, A.; Frittoli, E.; Garré, M.; Faretta, M.; Mione, M.; Testa, I.; Diaspro, A.; Lanzetti, L.; Scita, G.; Di Fiore, P.P. Endocytic trafficking of rac is required for the spatial restriction of signaling in cell migration. Cell 2008, 134, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.M.; Datta, A.; Rodriguez-Fraticelli, A.E.; Peranen, J.; Martin-Belmonte, F.; Mostov, K.E. A molecular network for de novo generation of the apical surface and lumen. Nat. Cell Biol. 2010, 12, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Ramel, D.; Wang, X.; Laflamme, C.; Montell, D.J.; Emery, G. Rab11 regulates cell-cell communication during collective cell movements. Nat. Cell Biol. 2013, 15, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wang, J.; Tang, Y.; Wang, Y.; Xiong, F.; Zhang, S.; Li, X.; Xiang, B.; Wu, X.; Guo, C.; et al. Long non-coding rna loc284454 promotes migration and invasion of nasopharyngeal carcinoma via modulating the rho/rac signaling pathway. Carcinogenesis 2018. [Google Scholar] [CrossRef]
- Tanabe, K.; Bonilla, I.; Winkles, J.A.; Strittmatter, S.M. Fibroblast growth factor-inducible-14 is induced in axotomized neurons and promotes neurite outgrowth. J. Neurosci. 2003, 23, 9675–9686. [Google Scholar] [CrossRef]
- Zamboni, V.; Armentano, M.; Berto, G.; Ciraolo, E.; Ghigo, A.; Garzotto, D.; Umbach, A.; DiCunto, F.; Parmigiani, E.; Boido, M.; et al. Hyperactivity of rac1-gtpase pathway impairs neuritogenesis of cortical neurons by altering actin dynamics. Sci. Rep. 2018, 8, 7254. [Google Scholar] [CrossRef]
- Kaufmann, N.; Wills, Z.P.; Van Vactor, D. Drosophila rac1 controls motor axon guidance. Development 1998, 125, 453–461. [Google Scholar]
- Aoki, K.; Nakamura, T.; Matsuda, M. Spatio-temporal regulation of rac1 and cdc42 activity during nerve growth factor-induced neurite outgrowth in pc12 cells. J. Biol. Chem. 2004, 279, 713–719. [Google Scholar] [CrossRef]
- Woo, S.; Gomez, T.M. Rac1 and rhoa promote neurite outgrowth through formation and stabilization of growth cone point contacts. J. Neurosci. 2006, 26, 1418–1428. [Google Scholar] [CrossRef]
- Bucci, C.; Alifano, P.; Cogli, L. The role of rab proteins in neuronal cells and in the trafficking of neurotrophin receptors. Membranes 2014, 4, 642–677. [Google Scholar] [CrossRef]
- Kiral, F.R.; Kohrs, F.E.; Jin, E.J.; Hiesinger, P.R. Rab gtpases and membrane trafficking in neurodegeneration. Curr. Biol. 2018, 28, R471–R486. [Google Scholar] [CrossRef]
- Kawauchi, T.; Chihama, K.; Nabeshima, Y.; Hoshino, M. The in vivo roles of stef/tiam1, rac1 and jnk in cortical neuronal migration. EMBO J. 2003, 22, 4190–4201. [Google Scholar] [CrossRef]
- Yang, T.; Sun, Y.; Zhang, F.; Zhu, Y.; Shi, L.; Li, H.; Xu, Z. Posh localizes activated rac1 to control the formation of cytoplasmic dilation of the leading process and neuronal migration. Cell Rep. 2012, 2, 640–651. [Google Scholar] [CrossRef]
- Kholmanskikh, S.S.; Dobrin, J.S.; Wynshaw-Boris, A.; Letourneau, P.C.; Ross, M.E. Disregulated rhogtpases and actin cytoskeleton contribute to the migration defect in lis1-deficient neurons. J. Neurosci. 2003, 23, 8673–8681. [Google Scholar] [CrossRef]
- Hirotsune, S.; Fleck, M.W.; Gambello, M.J.; Bix, G.J.; Chen, A.; Clark, G.D.; Ledbetter, D.H.; McBain, C.J.; Wynshaw-Boris, A. Graded reduction of pafah1b1 (lis1) activity results in neuronal migration defects and early embryonic lethality. Nat. Genet. 1998, 19, 333–339. [Google Scholar] [CrossRef]
- Houalla, T.; Shi, L.; van Meyel, D.J.; Rao, Y. Rab-mediated vesicular transport is required for neuronal positioning in the developing drosophila visual system. Mol. Brain 2010, 3, 19. [Google Scholar] [CrossRef]
- Park, H.T.; Feltri, M.L. Rac1 gtpase controls myelination and demyelination. Bioarchitecture 2011, 1, 110–113. [Google Scholar] [CrossRef]
- Casey, L.; Riyadh, M.A.; Yan Ho, X.; Neumann, B.; Giordano-Santini, R.; Hilliard, M.A. Disruption of rab-5 increases eff-1 fusogen availability at the cell surface and promotes the regenerative axonal fusion capacity of the neuron. J. Neurosci. 2019, 39, 2823–2836. [Google Scholar]
- Egami, Y.; Kiryu-Seo, S.; Yoshimori, T.; Kiyama, H. Induced expressions of rab24 gtpase and lc3 in nerve-injured motor neurons. Biochem. Biophys. Res. Commun. 2005, 337, 1206–1213. [Google Scholar] [CrossRef]
- Topp, J.D.; Gray, N.W.; Gerard, R.D.; Horazdovsky, B.F. Alsin is a rab5 and rac1 guanine nucleotide exchange factor. J. Biol. Chem. 2004, 279, 24612–24623. [Google Scholar] [CrossRef] [PubMed]
- Bucci, C.; De Luca, M. Molecular basis of charcot-marie-tooth type 2b disease. Biochem. Soc. Trans. 2012, 40, 1368–1372. [Google Scholar] [CrossRef]
- Deinhardt, K.; Salinas, S.; Verastegui, C.; Watson, R.; Worth, D.; Hanrahan, S.; Bucci, C.; Schiavo, G. Rab5 and rab7 control endocytic sorting along the axonal retrograde transport pathway. Neuron 2006, 52, 293–305. [Google Scholar] [CrossRef]
- Saxena, S.; Bucci, C.; Weis, J.; Kruttgen, A. The small gtpase rab7 controls the endosomal trafficking and neuritogenic signaling of the nerve growth factor receptor trka. J. Neurosci. 2005, 25, 10930–10940. [Google Scholar] [CrossRef]
- Cogli, L.; Progida, C.; Lecci, R.; Bramato, R.; Krüttgen, A.; Bucci, C. Cmt2b-associated rab7 mutants inhibit neurite outgrowth. Acta Neuropathol. 2010, 120, 491–501. [Google Scholar] [CrossRef]
- Yamauchi, J.; Torii, T.; Kusakawa, S.; Sanbe, A.; Nakamura, K.; Takashima, S.; Hamasaki, H.; Kawaguchi, S.; Miyamoto, Y.; Tanoue, A. The mood stabilizer valproic acid improves defective neurite formation caused by charcot-marie-tooth disease-associated mutant rab7 through the jnk signaling pathway. J. Neurosci. Res. 2010, 88, 3189–3197. [Google Scholar] [CrossRef] [PubMed]
- Ponomareva, O.Y.; Eliceiri, K.W.; Halloran, M.C. Charcot-marie-tooth 2b associated rab7 mutations cause axon growth and guidance defects during vertebrate sensory neuron development. Neural. Dev. 2016, 11, 2. [Google Scholar] [CrossRef]
- Kawauchi, T.; Sekine, K.; Shikanai, M.; Chihama, K.; Tomita, K.; Kubo, K.; Nakajima, K.; Nabeshima, Y.; Hoshino, M. Rab gtpases-dependent endocytic pathways regulate neuronal migration and maturation through n-cadherin trafficking. Neuron 2010, 67, 588–602. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, T. Regulation of cell adhesion and migration in cortical neurons: Not only rho but also rab family small gtpases. Small GTPases 2011, 2, 36–40. [Google Scholar] [CrossRef]
- Colecchia, D.; Stasi, M.; Leonardi, M.; Manganelli, F.; Nolano, M.; Veneziani, B.M.; Santoro, L.; Eskelinen, E.-L.; Chiariello, M.; Bucci, C. Alterations of autophagy in charcot-marie-tooth type 2b. Autophagy 2018, 14, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.M.; Li, X.; Xu, M.; Abais, J.M.; Chen, Y.; Riebling, C.R.; Boini, K.M.; Li, P.L.; Zhang, Y. Enhancement of autophagy by simvastatin through inhibition of rac1-mtor signaling pathway in coronary arterial myocytes. Cell Physiol. Biochem. 2013, 31, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Hu, X.; Li, N.; Hu, F.; Chang, F.; Xu, H.F.; Liu, Y.J. Distinctive roles of rac1 and rab29 in lrrk2 mediated membrane trafficking and neurite outgrowth. J. Biomed. Res. 2018, 32, 145–156. [Google Scholar] [PubMed]
- Chevallier, J.; Koop, C.; Srivastava, A.; Petrie, R.J.; Lamarche-Vane, N.; Presley, J.F. Rab35 regulates neurite outgrowth and cell shape. FEBS Lett. 2009, 583, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Turvey, S.E.; Broide, D.H. Innate immunity. J. Allergy Clin. Immunol. 2010, 125, S24–S32. [Google Scholar] [CrossRef]
- Netea, M.G.; Schlitzer, A.; Placek, K.; Joosten, L.A.B.; Schultze, J.L. Innate and adaptive immune memory: An evolutionary continuum in the host's response to pathogens. Cell Host. Microbe. 2019, 25, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Kuijl, C.; Pilli, M.; Alahari, S.K.; Janssen, H.; Khoo, P.S.; Ervin, K.E.; Calero, M.; Jonnalagadda, S.; Scheller, R.H.; Neefjes, J.; et al. Rac and rab gtpases dual effector nischarin regulates vesicle maturation to facilitate survival of intracellular bacteria. EMBO J. 2013, 32, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Nehme, N.T.; Quintin, J.; Cho, J.H.; Lee, J.; Lafarge, M.C.; Kocks, C.; Ferrandon, D. Relative roles of the cellular and humoral responses in the drosophila host defense against three gram-positive bacterial infections. PLoS ONE 2011, 6, e14743. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Lee, S.M.; Lee, M.S.; Yoon, J.; Kweon, H.S.; Kim, Y.J. Rab35 mediates transport of cdc42 and rac1 to the plasma membrane during phagocytosis. Mol. Cell Biol. 2010, 30, 1421–1433. [Google Scholar] [CrossRef]
- Zhao, H.; Laitala-Leinonen, T.; Parikka, V.; Väänänen, H.K. Downregulation of small gtpase rab7 impairs osteoclast polarization and bone resorption. J. Biol. Chem. 2001, 276, 39295–39302. [Google Scholar] [CrossRef]
- Mensah, K.A.; Schwarz, E.M.; Ritchlin, C.T. Altered bone remodeling in psoriatic arthritis. Curr. Rheumatol. Rep. 2008, 10, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Gennari, L.; Rendina, D.; Falchetti, A.; Merlotti, D. Paget's disease of bone. Calcif. Tissue Int. 2019, 1–18. [Google Scholar] [CrossRef]
- Ng, P.Y.; Brigitte Patricia Ribet, A.; Pavlos, N.J. Membrane trafficking in osteoclasts and implications for osteoporosis. Biochem. Soc. Trans. 2019. [Google Scholar] [CrossRef] [PubMed]
- Vallet, S.; Filzmoser, J.M.; Pecherstorfer, M.; Podar, K. Myeloma bone disease: Update on pathogenesis and novel treatment strategies. Pharmaceutics 2018, 10, 202. [Google Scholar] [CrossRef]
- Bouchet, J.; Del Rio-Iniguez, I.; Lasserre, R.; Aguera-Gonzalez, S.; Cuche, C.; Danckaert, A.; McCaffrey, M.W.; Di Bartolo, V.; Alcover, A. Rac1-rab11-fip3 regulatory hub coordinates vesicle traffic with actin remodeling and t-cell activation. EMBO J. 2016, 35, 1160–1174. [Google Scholar] [CrossRef]
- Ong, S.T.; Freeley, M.; Skubis-Zegadlo, J.; Fazil, M.H.; Kelleher, D.; Fresser, F.; Baier, G.; Verma, N.K.; Long, A. Phosphorylation of rab5a protein by protein kinase c is crucial for t-cell migration. J. Biol. Chem. 2014, 289, 19420–19434. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of Modification | Chemical Group Added | Aminoacidic Residue | Effect | Reference |
|---|---|---|---|---|
| Prenylation | Prenyl group | Cys178 | Rac1 localization and activity | [2] |
| Palmitoylation | Palmitoyl group | |||
| Ubiquitination | Ubiquitin | Lys147 | Rac1 degradation and regulation of its activity | [4,5,9] |
| SUMOylation | Small Ubiquitin-related Modifier (SUMO) proteins | Lysine residues; C-terminal domain | Rac1 activity and cell migration | [6] |
| Phosphorylation | Phosphate group | Ser71 | Rac1 GTP-binding activity | [7] |
| Phosphorylation | Phosphate group | Thr108 | Rac1 activity, interaction with PLC-γ1 and cell migration | [8] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margiotta, A.; Bucci, C. Coordination between Rac1 and Rab Proteins: Functional Implications in Health and Disease. Cells 2019, 8, 396. https://doi.org/10.3390/cells8050396
Margiotta A, Bucci C. Coordination between Rac1 and Rab Proteins: Functional Implications in Health and Disease. Cells. 2019; 8(5):396. https://doi.org/10.3390/cells8050396
Chicago/Turabian StyleMargiotta, Azzurra, and Cecilia Bucci. 2019. "Coordination between Rac1 and Rab Proteins: Functional Implications in Health and Disease" Cells 8, no. 5: 396. https://doi.org/10.3390/cells8050396
APA StyleMargiotta, A., & Bucci, C. (2019). Coordination between Rac1 and Rab Proteins: Functional Implications in Health and Disease. Cells, 8(5), 396. https://doi.org/10.3390/cells8050396