The Vav GEF Family: An Evolutionary and Functional Perspective



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
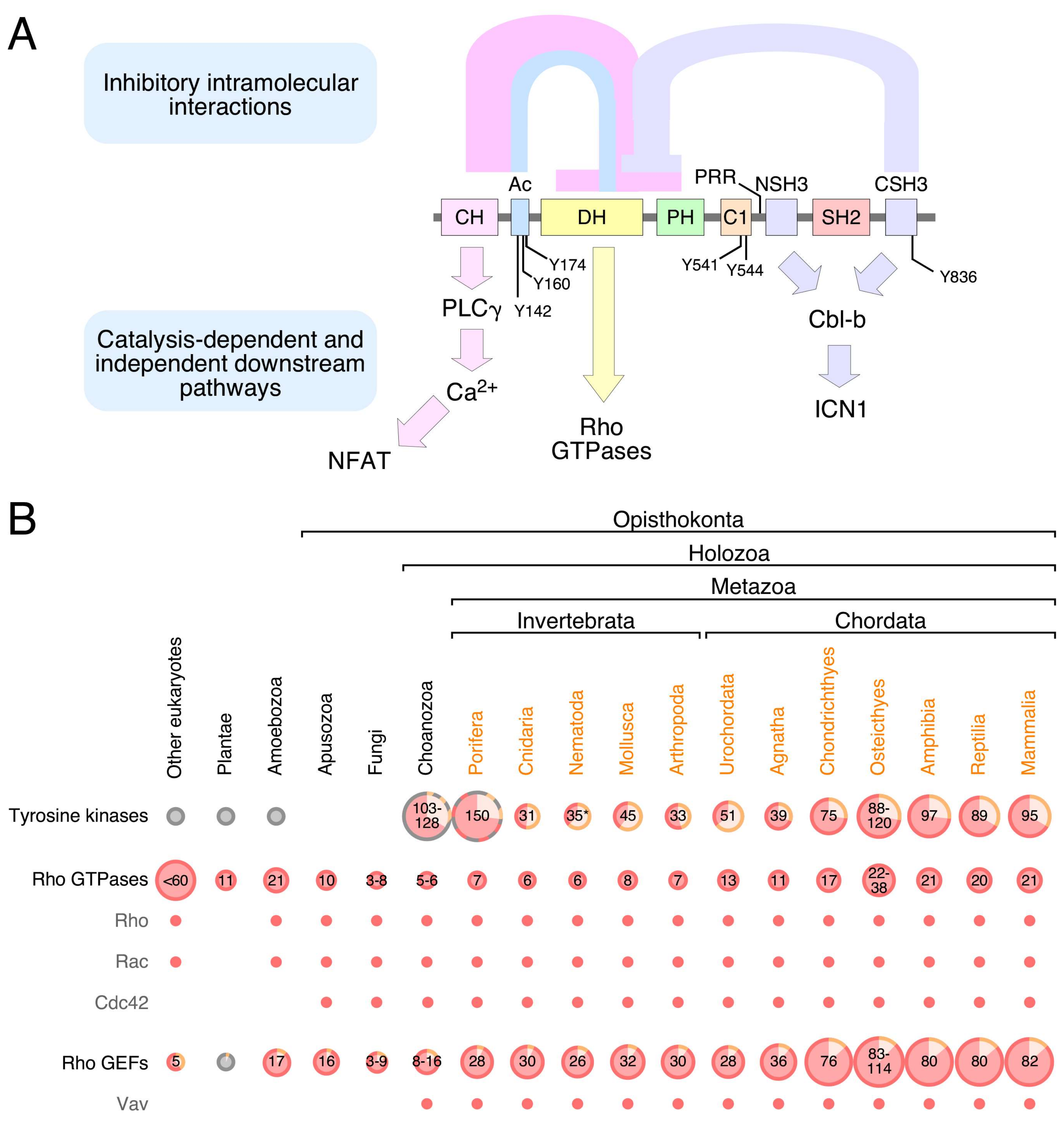
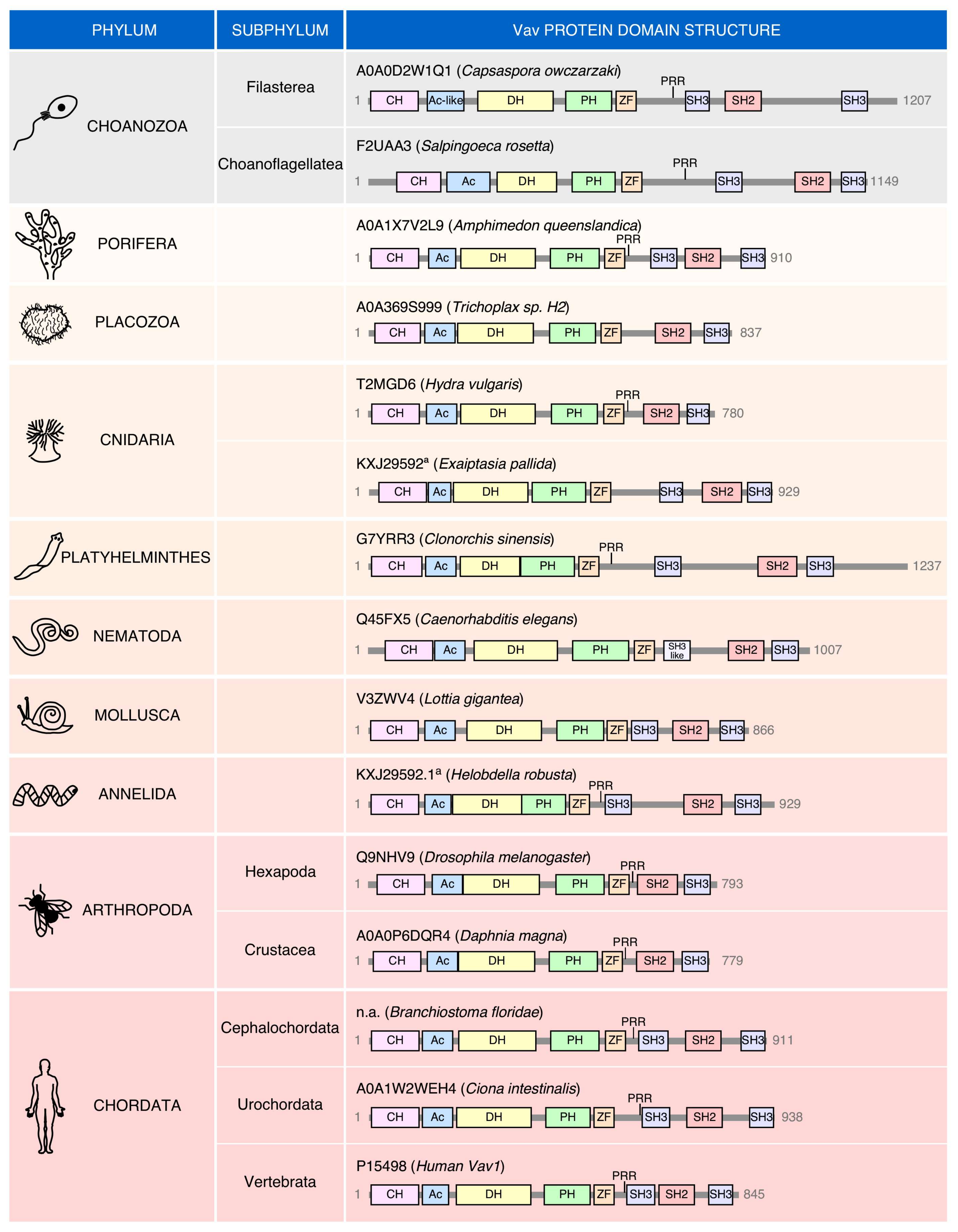
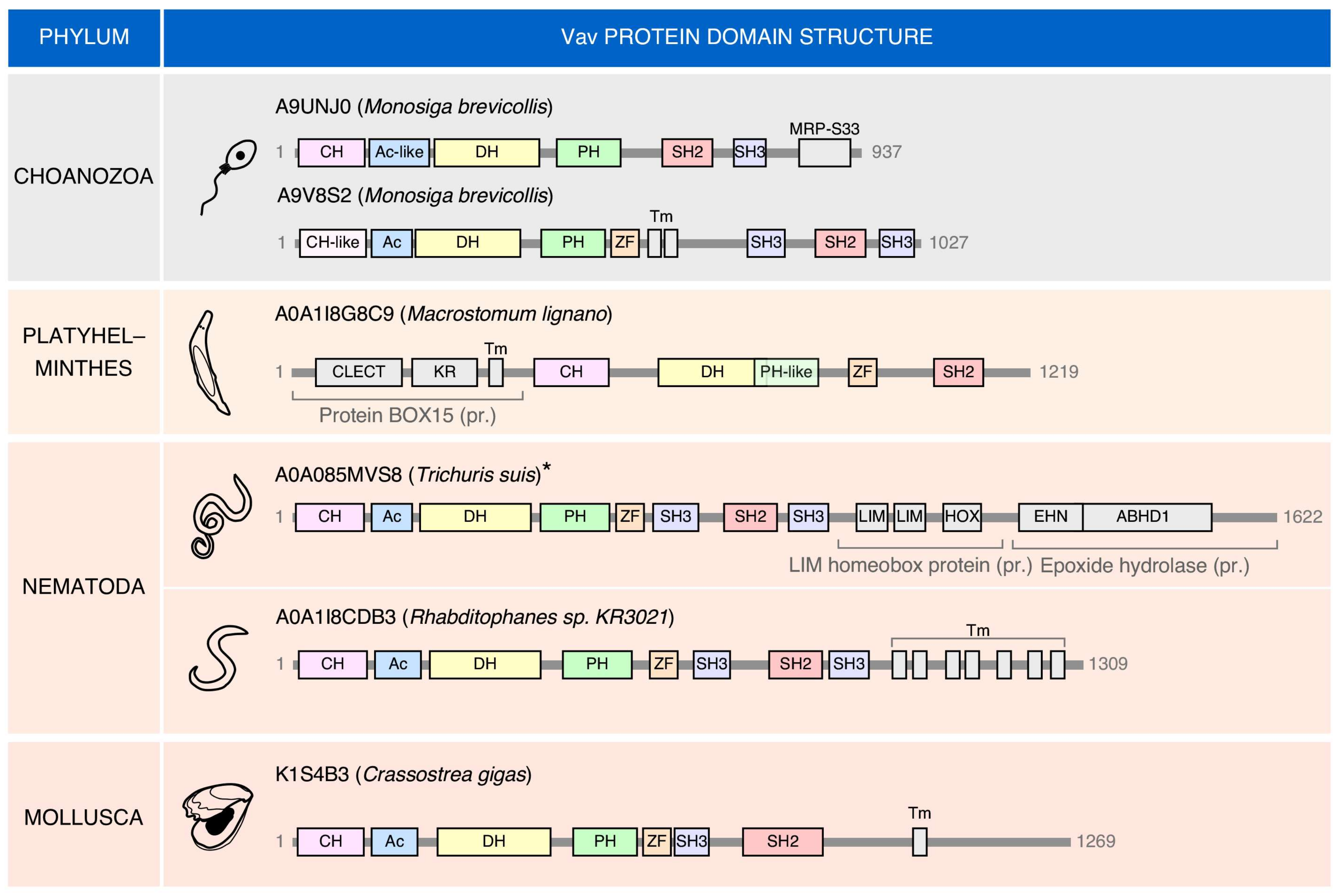
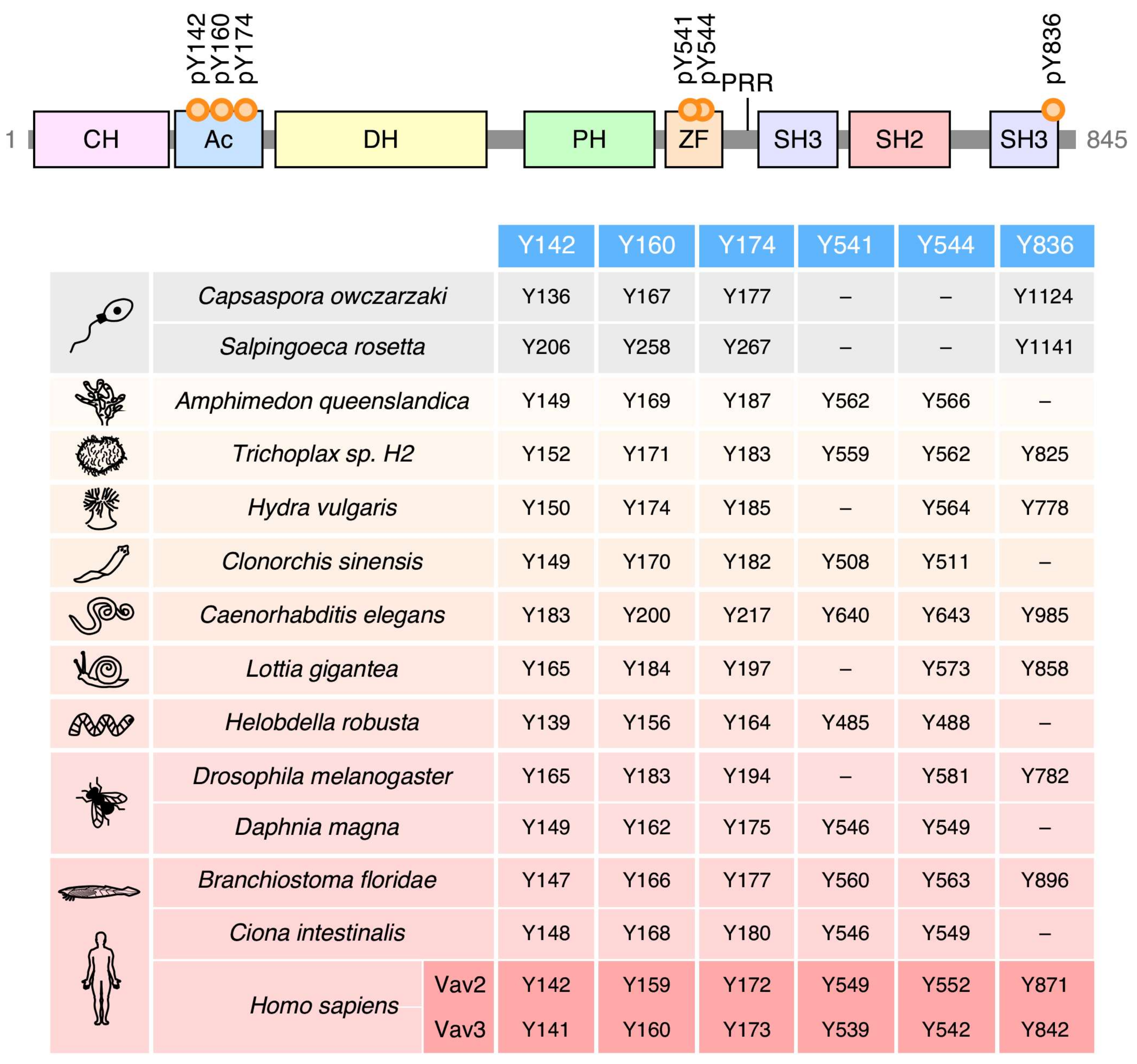
2. Structure
3. Signaling Functions
4. Regulation
5. Phylogenetic Evolution
6. Physiological Roles
7. Final Remarks
Funding
Conflicts of Interest
References
- Rossman, K.L.; Der, C.J.; Sondek, J. GEF means go: Turning on RHO GTPases with guanine nucleotide–exchange factors. Nat. Rev. Mol. Cell Biol. 2005, 6, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP–binding proteins of the Rho/Rac family: Regulation, effectors and functions in vivo. Bioessays 2007, 29, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R. RHO GTPases in cancer: Known facts, open questions, and therapeutic challenges. Biochem. Soc. Trans. 2018, 46, 741–760. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R. Vav family exchange factors: An integrated regulatory and functional view. Small GTPases 2014, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Katzav, S.; Martin-Zanca, D.; Barbacid, M. Vav, a novel human oncogene derived from a locus ubiquitously expressed in hematopoietic cells. EMBO J. 1989, 8, 2283–2290. [Google Scholar] [CrossRef] [PubMed]
- Henske, E.P.; Short, M.P.; Jozwiak, S.; Bovey, C.M.; Ramlakhan, S.; Haines, J.L.; Kwiatkowski, D.J. Identification of VAV2 on 9q34 and its exclusion as the tuberous sclerosis gene TSC1. Ann. Hum. Genet. 1995, 59, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Schuebel, K.E.; Bustelo, X.R.; Nielsen, D.A.; Song, B.J.; Barbacid, M.; Goldman, D.; Lee, I.J. Isolation and characterization of murine vav2, a member of the vav family of proto–oncogenes. Oncogene 1996, 13, 363–371. [Google Scholar] [PubMed]
- Movilla, N.; Bustelo, X.R. Biological and regulatory properties of Vav–3, a new member of the Vav family of oncoproteins. Mol. Cell Biol. 1999, 19, 7870–7885. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R.; Dosil, M. The Vav family. In Encyclopedia of Signaling Molecules; Choi, S., Ed.; Springer: New York, NY, USA, 2016; pp. 1–15. [Google Scholar]
- Yu, B.; Martins, I.R.; Li, P.; Amarasinghe, G.K.; Umetani, J.; Fernandez-Zapico, M.E.; Billadeau, D.D.; Machius, M.; Tomchick, D.R.; Rosen, M.K. Structural and energetic mechanisms of cooperative autoinhibition and activation of Vav1. Cell 2010, 140, 246–256. [Google Scholar] [CrossRef]
- Rapley, J.; Tybulewicz, V.L.; Rittinger, K. Crucial structural role for the PH and C1 domains of the Vav1 exchange factor. EMBO Rep. 2008, 9, 655–661. [Google Scholar] [CrossRef] [Green Version]
- Chrencik, J.E.; Brooun, A.; Zhang, H.; Mathews, I.I.; Hura, G.L.; Foster, S.A.; Perry, J.J.; Streiff, M.; Ramage, P.; Widmer, H.; et al. Structural basis of guanine nucleotide exchange mediated by the T–cell essential Vav1. J. Mol. Biol. 2008, 380, 828–843. [Google Scholar] [CrossRef] [PubMed]
- Barreira, M.; Fabbiano, S.; Couceiro, J.R.; Torreira, E.; Martinez-Torrecuadrada, J.L.; Montoya, G.; Llorca, O.; Bustelo, X.R. The C–terminal SH3 domain contributes to the intramolecular inhibition of Vav family proteins. Sci. Signal 2014, 7, ra35. [Google Scholar] [CrossRef]
- Schuebel, K.E.; Movilla, N.; Rosa, J.L.; Bustelo, X.R. Phosphorylation–dependent and constitutive activation of Rho proteins by wild–type and oncogenic Vav–2. EMBO J. 1998, 17, 6608–6621. [Google Scholar] [CrossRef] [PubMed]
- Zugaza, J.L.; Lopez-Lago, M.A.; Caloca, M.J.; Dosil, M.; Movilla, N.; Bustelo, X.R. Structural determinants for the biological activity of Vav proteins. J. Biol. Chem. 2002, 277, 45377–45392. [Google Scholar] [CrossRef]
- Nishida, M.; Nagata, K.; Hachimori, Y.; Horiuchi, M.; Ogura, K.; Mandiyan, V.; Schlessinger, J.; Inagaki, F. Novel recognition mode between Vav and Grb2 SH3 domains. EMBO J. 2001, 20, 2995–3007. [Google Scholar] [CrossRef] [Green Version]
- Ogura, K.; Nagata, K.; Horiuchi, M.; Ebisui, E.; Hasuda, T.; Yuzawa, S.; Nishida, M.; Hatanaka, H.; Inagaki, F. Solution structure of N–terminal SH3 domain of Vav and the recognition site for Grb2 C–terminal SH3 domain. J. Biomol. NMR 2002, 22, 37–46. [Google Scholar] [CrossRef]
- Mayer, B.J. What Have We Learned from SH2 Domains? Methods Mol. Biol. 2017, 1555, 37–43. [Google Scholar] [CrossRef]
- Fort, P.; Blangy, A. The Evolutionary Landscape of Dbl–Like RhoGEF Families: Adapting Eukaryotic Cells to Environmental Signals. Genome Biol. Evol. 2017, 9, 1471–1486. [Google Scholar] [CrossRef]
- Brunet, F.G.; Volff, J.N.; Schartl, M. Whole Genome Duplications Shaped the Receptor Tyrosine Kinase Repertoire of Jawed Vertebrates. Genome Biol. Evol. 2016, 8, 1600–1613. [Google Scholar] [CrossRef] [Green Version]
- Pincus, D.; Letunic, I.; Bork, P.; Lim, W.A. Evolution of the phospho–tyrosine signaling machinery in premetazoan lineages. Proc. Natl. Acad. Sci. USA 2008, 105, 9680–9684. [Google Scholar] [CrossRef]
- Liu, A.; He, F.; Gu, X. Identification and characterization of tyrosine kinases in anole lizard indicate the conserved tyrosine kinase repertoire in vertebrates. Mol. Genet. Genom. 2017, 292, 1405–1418. [Google Scholar] [CrossRef]
- Rojas, A.M.; Fuentes, G.; Rausell, A.; Valencia, A. The Ras protein superfamily: Evolutionary tree and role of conserved amino acids. J. Cell Biol. 2012, 196, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boureux, A.; Vignal, E.; Faure, S.; Fort, P. Evolution of the Rho family of ras–like GTPases in eukaryotes. Mol. Biol. Evol. 2007, 24, 203–216. [Google Scholar] [CrossRef]
- Elias, M.; Klimes, V. Rho GTPases: deciphering the evolutionary history of a complex protein family. Methods Mol. Biol. 2012, 827, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.A.; Nash, P.D. Evolution of SH2 domains and phosphotyrosine signalling networks. Philos. Trans. R. Soc. Lond B Biol. Sci. 2012, 367, 2556–2573. [Google Scholar] [CrossRef] [Green Version]
- Crespo, P.; Schuebel, K.E.; Ostrom, A.A.; Gutkind, J.S.; Bustelo, X.R. Phosphotyrosine–dependent activation of Rac–1 GDP/GTP exchange by the vav proto–oncogene product. Nature 1997, 385, 169–172. [Google Scholar] [CrossRef]
- Movilla, N.; Dosil, M.; Zheng, Y.; Bustelo, X.R. How Vav proteins discriminate the GTPases Rac1 and RhoA from Cdc42. Oncogene 2001, 20, 8057–8065. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Rossman, K.L.; Liu, B.; Ritola, K.D.; Chiang, D.; Campbell, S.L.; Burridge, K.; Der, C.J. Vav2 is an activator of Cdc42, Rac1, and RhoA. J. Biol. Chem. 2000, 275, 10141–10149. [Google Scholar] [CrossRef]
- Wu, J.; Katzav, S.; Weiss, A. A functional T–cell receptor signaling pathway is required for p95vav activity. Mol. Cell. Biol. 1995, 15, 4337–4346. [Google Scholar] [CrossRef]
- Kuhne, M.R.; Ku, G.; Weiss, A. A guanine nucleotide exchange factor–independent function of Vav1 in transcriptional activation. J. Biol. Chem. 2000, 275, 2185–2190. [Google Scholar] [CrossRef]
- Zhou, Z.; Yin, J.; Dou, Z.; Tang, J.; Zhang, C.; Cao, Y. The calponin homology domain of Vav1 associates with calmodulin and is prerequisite to T cell antigen receptor–induced calcium release in Jurkat T lymphocytes. J. Biol. Chem. 2007, 282, 23737–23744. [Google Scholar] [CrossRef]
- Saveliev, A.; Vanes, L.; Ksionda, O.; Rapley, J.; Smerdon, S.J.; Rittinger, K.; Tybulewicz, V.L. Function of the nucleotide exchange activity of vav1 in T cell development and activation. Sci. Signal 2009, 2, ra83. [Google Scholar] [CrossRef]
- Tybulewicz, V.L. Vav–family proteins in T–cell signalling. Curr. Opin. Immunol. 2005, 17, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.F.; Smyth, L.A.; Norton, T.; Freshney, N.; Downward, J.; Kioussis, D.; Tybulewicz, V.L. Vav1 transduces T cell receptor signals to the activation of phospholipase C–gamma1 via phosphoinositide 3–kinase–dependent and –independent pathways. J. Exp. Med. 2002, 195, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, N.; Wong, J.F.; Tan, K.P.; Chung, H.H.; Yau, Y.H.; Cukuroglu, E.; Allahverdi, A.; Nordenskiold, L.; Goke, J.; Geifman-Shochat, S.; et al. EZH2 promotes neoplastic transformation through VAV interaction–dependent extranuclear mechanisms. Oncogene 2018, 37, 461–477. [Google Scholar] [CrossRef] [PubMed]
- Robles-Valero, J.; Lorenzo-Martin, L.F.; Menacho-Marquez, M.; Fernandez-Pisonero, I.; Abad, A.; Camos, M.; Toribio, M.L.; Espinosa, L.; Bigas, A.; Bustelo, X.R. A Paradoxical Tumor–Suppressor Role for the Rac1 Exchange Factor Vav1 in T Cell Acute Lymphoblastic Leukemia. Cancer Cell 2017, 32, 608–623.e9. [Google Scholar] [CrossRef]
- Robles-Valero, J.; Lorenzo-Martin, L.F.; Fernandez-Pisonero, I.; Bustelo, X.R. Rho guanosine nucleotide exchange factors are not such bad guys after all in cancer. Small GTPases 2018, 1–7. [Google Scholar] [CrossRef]
- Bustelo, X.R.; Lorenzo-Martin, L.F.; Cuadrado, M.; Fernandez-Pisonero, I.; Robles-Valero, J. An unexpected tumor suppressor role for VAV1. Mol. Cell. Oncol. 2018, 5, e1432257. [Google Scholar] [CrossRef]
- Citterio, C.; Menacho-Marquez, M.; Garcia-Escudero, R.; Larive, R.M.; Barreiro, O.; Sanchez-Madrid, F.; Paramio, J.M.; Bustelo, X.R. The rho exchange factors vav2 and vav3 control a lung metastasis–specific transcriptional program in breast cancer cells. Sci. Signal 2012, 5, ra71. [Google Scholar] [CrossRef] [PubMed]
- Houlard, M.; Arudchandran, R.; Regnier-Ricard, F.; Germani, A.; Gisselbrecht, S.; Blank, U.; Rivera, J.; Varin-Blank, N. Vav1 is a component of transcriptionally active complexes. J. Exp. Med. 2002, 195, 1115–1127. [Google Scholar] [CrossRef]
- Wang, R.; Wang, J.; Zhang, N.; Wan, Y.; Liu, Y.; Zhang, L.; Pan, S.; Zhang, C.; Zhang, H.; Cao, Y. The interaction between Vav1 and EBNA1 promotes survival of Burkitt’s lymphoma cells by down–regulating the expression of Bim. Biochem. Biophys. Res. Commun. 2019, 511, 787–793. [Google Scholar] [CrossRef]
- Lyons, L.S.; Burnstein, K.L. Vav3, a Rho GTPase guanine nucleotide exchange factor, increases during progression to androgen independence in prostate cancer cells and potentiates androgen receptor transcriptional activity. Mol. Endocrinol. 2006, 20, 1061–1072. [Google Scholar] [CrossRef]
- Rao, S.; Lyons, L.S.; Fahrenholtz, C.D.; Wu, F.; Farooq, A.; Balkan, W.; Burnstein, K.L. A novel nuclear role for the Vav3 nucleotide exchange factor in androgen receptor coactivation in prostate cancer. Oncogene 2012, 31, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Liu, Y.; Lu, S.; Wang, A.; Lee, K.; Wang, L.H.; Revelo, M.; Lu, S. Vav3 oncogene is overexpressed and regulates cell growth and androgen receptor activity in human prostate cancer. Mol. Endocrinol. 2006, 20, 2315–2325. [Google Scholar] [CrossRef]
- Sauzeau, V.; Sevilla, M.A.; Montero, M.J.; Bustelo, X.R. The Rho/Rac exchange factor Vav2 controls nitric oxide–dependent responses in mouse vascular smooth muscle cells. J. Clin. Investig. 2010, 120, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Caloca, M.J.; Zugaza, J.L.; Bustelo, X.R. Mechanistic Analysis of the Amplification and Diversification Events Induced by Vav Proteins in B–lymphocytes. J. Biol. Chem. 2008, 283, 36454–36464. [Google Scholar] [CrossRef]
- Norman, K.R.; Fazzio, R.T.; Mellem, J.E.; Espelt, M.V.; Strange, K.; Beckerle, M.C.; Maricq, A.V. The Rho/Rac–family guanine nucleotide exchange factor VAV–1 regulates rhythmic behaviors in C. elegans. Cell 2005, 123, 119–132. [Google Scholar] [CrossRef]
- Caloca, M.J.; Zugaza, J.L.; Matallanas, D.; Crespo, P.; Bustelo, X.R. Vav mediates Ras stimulation by direct activation of the GDP/GTP exchange factor Ras GRP1. EMBO J. 2003, 22, 3326–3336. [Google Scholar] [CrossRef] [Green Version]
- Zugaza, J.L.; Caloca, M.J.; Bustelo, X.R. Inverted signaling hierarchy between RAS and RAC in T–lymphocytes. Oncogene 2004, 23, 5823–5833. [Google Scholar] [CrossRef]
- Costello, P.S.; Walters, A.E.; Mee, P.J.; Turner, M.; Reynolds, L.F.; Prisco, A.; Sarner, N.; Zamoyska, R.; Tybulewicz, V.L. The Rho–family GTP exchange factor Vav is a critical transducer of T cell receptor signals to the calcium, ERK, and NF–kappaB pathways. Proc. Natl. Acad. Sci. USA 1999, 96, 3035–3040. [Google Scholar] [CrossRef]
- Reynolds, L.F.; de Bettignies, C.; Norton, T.; Beeser, A.; Chernoff, J.; Tybulewicz, V.L. Vav1 transduces T cell receptor signals to the activation of the Ras/ERK pathway via LAT, Sos, and RasGRP1. J. Biol. Chem. 2004, 279, 18239–18246. [Google Scholar] [CrossRef]
- Doody, G.M.; Billadeau, D.D.; Clayton, E.; Hutchings, A.; Berland, R.; McAdam, S.; Leibson, P.J.; Turner, M. Vav–2 controls NFAT–dependent transcription in B– but not T–lymphocytes. EMBO J. 2000, 19, 6173–6184. [Google Scholar] [CrossRef]
- Lorenzo-Martin, L.F.; Citterio, C.; Menacho-Marquez, M.; Conde, J.; Larive, R.M.; Rodriguez-Fdez, S.; Garcia-Escudero, R.; Robles-Valero, J.; Cuadrado, M.; Fernandez-Pisonero, I.; et al. Vav proteins maintain epithelial traits in breast cancer cells using miR–200c–dependent and independent mechanisms. Oncogene 2019, 38, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh, B.; Lowry, W.E.; Huang, X.Y.; Rosen, M.K. Structural basis for relief of autoinhibition of the Dbl homology domain of proto–oncogene Vav by tyrosine phosphorylation. Cell 2000, 102, 625–633. [Google Scholar] [CrossRef]
- Lopez-Lago, M.; Lee, H.; Cruz, C.; Movilla, N.; Bustelo, X.R. Tyrosine phosphorylation mediates both activation and downmodulation of the biological activity of Vav. Mol. Cell. Biol. 2000, 20, 1678–1691. [Google Scholar] [CrossRef]
- Barreira, M.; Rodriguez-Fdez, S.; Bustelo, X.R. New insights into the Vav1 activation cycle in lymphocytes. Cell. Signal. 2018, 45, 132–144. [Google Scholar] [CrossRef]
- Bustelo, X.R.; Ledbetter, J.A.; Barbacid, M. Product of vav proto–oncogene defines a new class of tyrosine protein kinase substrates. Nature 1992, 356, 68–71. [Google Scholar] [CrossRef]
- Margolis, B.; Hu, P.; Katzav, S.; Li, W.; Oliver, J.M.; Ullrich, A.; Weiss, A.; Schlessinger, J. Tyrosine phosphorylation of vav proto–oncogene product containing SH2 domain and transcription factor motifs. Nature 1992, 356, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Weng, W.K.; Jarvis, L.; LeBien, T.W. Signaling through CD19 activates Vav/mitogen–activated protein kinase pathway and induces formation of a CD19/Vav/phosphatidylinositol 3–kinase complex in human B cell precursors. J. Biol. Chem. 1994, 269, 32514–32521. [Google Scholar]
- O’Rourke, L.M.; Tooze, R.; Turner, M.; Sandoval, D.M.; Carter, R.H.; Tybulewicz, V.L.; Fearon, D.T. CD19 as a membrane–anchored adaptor protein of B lymphocytes: Costimulation of lipid and protein kinases by recruitment of Vav. Immunity 1998, 8, 635–645. [Google Scholar] [CrossRef]
- Son, M.; Park, I.; Lee, O.H.; Rhee, I.; Park, C.; Yun, Y. LIME mediates immunological synapse formation through activation of VAV. Mol. Cells 2012, 33, 407–414. [Google Scholar] [CrossRef]
- Johmura, S.; Oh-hora, M.; Inabe, K.; Nishikawa, Y.; Hayashi, K.; Vigorito, E.; Kitamura, D.; Turner, M.; Shingu, K.; Hikida, M.; et al. Regulation of Vav localization in membrane rafts by adaptor molecules Grb2 and BLNK. Immunity 2003, 18, 777–787. [Google Scholar] [CrossRef]
- Wu, J.; Motto, D.G.; Koretzky, G.A.; Weiss, A. Vav and SLP–76 interact and functionally cooperate in IL–2 gene activation. Immunity 1996, 4, 593–602. [Google Scholar] [CrossRef]
- Tuosto, L.; Michel, F.; Acuto, O. p95vav associates with tyrosine–phosphorylated SLP–76 in antigen–stimulated T cells. J. Exp. Med. 1996, 184, 1161–1166. [Google Scholar] [CrossRef]
- Lindholm, C.K.; Henriksson, M.L.; Hallberg, B.; Welsh, M. Shb links SLP–76 and Vav with the CD3 complex in Jurkat T cells. Eur. J. Biochem. 2002, 269, 3279–3288. [Google Scholar] [CrossRef]
- Pauker, M.H.; Hassan, N.; Noy, E.; Reicher, B.; Barda-Saad, M. Studying the dynamics of SLP–76, Nck, and Vav1 multimolecular complex formation in live human cells with triple–color FRET. Sci. Signal 2012, 5, rs3. [Google Scholar] [CrossRef]
- Barda-Saad, M.; Shirasu, N.; Pauker, M.H.; Hassan, N.; Perl, O.; Balbo, A.; Yamaguchi, H.; Houtman, J.C.; Appella, E.; Schuck, P.; et al. Cooperative interactions at the SLP–76 complex are critical for actin polymerization. EMBO J. 2010, 29, 2315–2328. [Google Scholar] [CrossRef]
- Gomez, T.S.; Hamann, M.J.; McCarney, S.; Savoy, D.N.; Lubking, C.M.; Heldebrant, M.P.; Labno, C.M.; McKean, D.J.; McNiven, M.A.; Burkhardt, J.K.; et al. Dynamin 2 regulates T cell activation by controlling actin polymerization at the immunological synapse. Nat. Immunol. 2005, 6, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Sylvain, N.R.; Nguyen, K.; Bunnell, S.C. Vav1–mediated scaffolding interactions stabilize SLP–76 microclusters and contribute to antigen–dependent T cell responses. Sci. Signal 2011, 4, ra14. [Google Scholar] [CrossRef]
- Charvet, C.; Canonigo, A.J.; Billadeau, D.D.; Altman, A. Membrane localization and function of Vav3 in T cells depend on its association with the adapter SLP–76. J. Biol. Chem. 2005, 280, 15289–15299. [Google Scholar] [CrossRef] [PubMed]
- Couceiro, J.R.; Martin-Bermudo, M.D.; Bustelo, X.R. Phylogenetic conservation of the regulatory and functional properties of the Vav oncoprotein family. Exp. Cell. Res. 2005, 308, 364–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abate, F.; da Silva-Almeida, A.C.; Zairis, S.; Robles-Valero, J.; Couronne, L.; Khiabanian, H.; Quinn, S.A.; Kim, M.Y.; Laginestra, M.A.; Kim, C.; et al. Activating mutations and translocations in the guanine exchange factor VAV1 in peripheral T–cell lymphomas. Proc. Natl. Acad. Sci. USA 2017, 114, 764–769. [Google Scholar] [CrossRef]
- Boddicker, R.L.; Razidlo, G.L.; Dasari, S.; Zeng, Y.; Hu, G.; Knudson, R.A.; Greipp, P.T.; Davila, J.I.; Johnson, S.H.; Porcher, J.C.; et al. Integrated mate–pair and RNA sequencing identifies novel, targetable gene fusions in peripheral T–cell lymphoma. Blood 2016, 128, 1234–1245. [Google Scholar] [CrossRef]
- Campbell, J.D.; Alexandrov, A.; Kim, J.; Wala, J.; Berger, A.H.; Pedamallu, C.S.; Shukla, S.A.; Guo, G.; Brooks, A.N.; Murray, B.A. Distinct patterns of somatic genome alterations in lung adenocarcinomas and squamous cell carcinomas. Nat. Genet. 2016, 48, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, K.; Nagata, Y.; Kitanaka, A.; Shiraishi, Y.; Shimamura, T.; Yasunaga, J.; Totoki, Y.; Chiba, K.; Sato-Otsubo, A.; Nagae, G.; et al. Integrated molecular analysis of adult T cell leukemia/lymphoma. Nat. Genet. 2015, 47, 1304–1315. [Google Scholar] [CrossRef]
- Park, J.; Yang, J.; Wenzel, A.T.; Ramachandran, A.; Lee, W.J.; Daniels, J.C.; Kim, J.; Martinez-Escala, E.; Amankulor, N.; Pro, B.; et al. Genomic analysis of 220 CTCLs identifies a novel recurrent gain–of–function alteration in RLTPR (p.Q575E). Blood 2017, 130, 1430–1440. [Google Scholar] [CrossRef]
- Choudhary, C.; Kumar, C.; Gnad, F.; Nielsen, M.L.; Rehman, M.; Walther, T.C.; Olsen, J.V.; Mann, M. Lysine acetylation targets protein complexes and co–regulates major cellular functions. Science 2009, 325, 834–840. [Google Scholar] [CrossRef]
- Puga, I.; Rao, A.; Macian, F. Targeted cleavage of signaling proteins by caspase 3 inhibits T cell receptor signaling in anergic T cells. Immunity 2008, 29, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.G.; Hehner, S.P.; Droge, W.; Schmitz, M.L. Caspase–dependent cleavage and inactivation of the Vav1 proto–oncogene product during apoptosis prevents IL–2 transcription. Oncogene 2000, 19, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Razidlo, G.L.; Wang, Y.; Chen, J.; Krueger, E.W.; Billadeau, D.D.; McNiven, M.A. Dynamin 2 potentiates invasive migration of pancreatic tumor cells through stabilization of the Rac1 GEF Vav1. Dev. Cell 2013, 24, 573–585. [Google Scholar] [CrossRef]
- Yoo, A.S.; Greenwald, I. LIN–12/Notch activation leads to microRNA–mediated down–regulation of Vav in C. elegans. Science 2005, 310, 1330–1333. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, N.; Li, S.; Fang, J.H.; Chen, M.X.; Yang, J.; Jia, W.H.; Yuan, Y.; Zhuang, S.M. MicroRNA–195 suppresses angiogenesis and metastasis of hepatocellular carcinoma by inhibiting the expression of VEGF, VAV2, and CDC42. Hepatology 2013, 58, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Ding, L.; Bennewith, K.L.; Tong, R.T.; Welford, S.M.; Ang, K.K.; Story, M.; Le, Q.T.; Giaccia, A.J. Hypoxia–inducible mir–210 regulates normoxic gene expression involved in tumor initiation. Mol. Cell 2009, 35, 856–867. [Google Scholar] [CrossRef]
- Fernandez-Zapico, M.E.; Gonzalez-Paz, N.C.; Weiss, E.; Savoy, D.N.; Molina, J.R.; Fonseca, R.; Smyrk, T.C.; Chari, S.T.; Urrutia, R.; Billadeau, D.D. Ectopic expression of VAV1 reveals an unexpected role in pancreatic cancer tumorigenesis. Cancer Cell 2005, 7, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvajal-Gonzalez, J.M.; Mulero-Navarro, S.; Roman, A.C.; Sauzeau, V.; Merino, J.M.; Bustelo, X.R.; Fernandez-Salguero, P.M. The dioxin receptor regulates the constitutive expression of the vav3 proto–oncogene and modulates cell shape and adhesion. Mol. Biol. Cell 2009, 20, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Ilan, L.; Katzav, S. Human Vav1 expression in hematopoietic and cancer cell lines is regulated by c–Myb and by CpG methylation. PLoS ONE 2012, 7, e29939. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Raja, S.M.; Chen, G.; Virmani, S.; Williams, S.H.; Clubb, R.J.; Mukhopadhyay, C.; Rainey, M.A.; Ying, G.; Dimri, M.; et al. Negative regulation of EGFR–Vav2 signaling axis by Cbl ubiquitin ligase controls EGF receptor–mediated epithelial cell adherens junction dynamics and cell migration. J. Biol. Chem. 2011, 286, 620–633. [Google Scholar] [CrossRef]
- Sauzeau, V.; Carvajal-Gonzalez, J.M.; Riolobos, A.S.; Sevilla, M.A.; Menacho-Marquez, M.; Roman, A.C.; Abad, A.; Montero, M.J.; Fernandez-Salguero, P.; Bustelo, X.R. Transcriptional factor aryl hydrocarbon receptor (Ahr) controls cardiovascular and respiratory functions by regulating the expression of the Vav3 proto–oncogene. J. Biol. Chem. 2011, 286, 2896–2909. [Google Scholar] [CrossRef]
- Blanchet, F.; Cardona, A.; Letimier, F.A.; Hershfield, M.S.; Acuto, O. CD28 costimulatory signal induces protein arginine methylation in T cells. J. Exp. Med. 2005, 202, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Jagodic, M.; Colacios, C.; Nohra, R.; Dejean, A.S.; Beyeen, A.D.; Khademi, M.; Casemayou, A.; Lamouroux, L.; Duthoit, C.; Papapietro, O.; et al. A role for VAV1 in experimental autoimmune encephalomyelitis and multiple sclerosis. Sci. Transl. Med. 2009, 1, 10ra21. [Google Scholar] [CrossRef]
- Kassem, S.; Gaud, G.; Bernard, I.; Benamar, M.; Dejean, A.S.; Liblau, R.; Fournie, G.J.; Colacios, C.; Malissen, B.; Saoudi, A. A Natural Variant of the T Cell Receptor–Signaling Molecule Vav1 Reduces Both Effector T Cell Functions and Susceptibility to Neuroinflammation. PLoS Genet. 2016, 12, e1006185. [Google Scholar] [CrossRef]
- Colacios, C.; Casemayou, A.; Dejean, A.S.; Gaits-Iacovoni, F.; Pedros, C.; Bernard, I.; Lagrange, D.; Deckert, M.; Lamouroux, L.; Jagodic, M.; et al. The p.Arg63Trp polymorphism controls Vav1 functions and Foxp3 regulatory T cell development. J. Exp. Med. 2011, 208, 2183–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, I.; Sacquin, A.; Kassem, S.; Benamar, M.; Colacios, C.; Gador, M.; Perals, C.; Fazilleau, N.; Saoudi, A. A Natural Variant of the Signaling Molecule Vav1 Enhances Susceptibility to Myasthenia Gravis and Influences the T Cell Receptor Repertoire. Front. Immunol. 2018, 9, 2399. [Google Scholar] [CrossRef]
- Roth, S.; Bergmann, H.; Jaeger, M.; Yeroslaviz, A.; Neumann, K.; Koenig, P.A.; Prazeres da Costa, C.; Vanes, L.; Kumar, V.; Johnson, M.; et al. Vav Proteins Are Key Regulators of Card9 Signaling for Innate Antifungal Immunity. Cell Rep. 2016, 17, 2572–2583. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Li, G.; Wang, L. The interaction effect of rs4077515 and rs17019602 increases the susceptibility to IgA nephropathy. Oncotarget 2017, 8, 76492–76497. [Google Scholar] [CrossRef] [Green Version]
- Perretta-Tejedor, N.; Fernandez-Mateos, J.; Garcia-Ortiz, L.; Gomez-Marcos, M.A.; Recio-Rodriguez, J.I.; Agudo-Conde, C.; Rodriguez-Sanchez, E.; Morales, A.I.; Lopez-Hernandez, F.J.; Lopez-Novoa, J.M.; et al. Association of VAV2 and VAV3 polymorphisms with cardiovascular risk factors. Sci. Rep. 2017, 7, 41875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, H.Y.; Hwangbo, Y.; Yoo, S.K.; Im, S.W.; Yang, S.D.; Kwak, S.J.; Park, M.S.; Kwak, S.H.; Cho, S.W.; Ryu, J.S.; et al. Genome–wide association and expression quantitative trait loci studies identify multiple susceptibility loci for thyroid cancer. Nat. Commun. 2017, 8, 15966. [Google Scholar] [CrossRef] [PubMed]
- Kiryluk, K.; Li, Y.; Scolari, F.; Sanna-Cherchi, S.; Choi, M.; Verbitsky, M.; Fasel, D.; Lata, S.; Prakash, S.; Shapiro, S.; et al. Discovery of new risk loci for IgA nephropathy implicates genes involved in immunity against intestinal pathogens. Nat. Genet. 2014, 46, 1187–1196. [Google Scholar] [CrossRef] [Green Version]
- Manning, G.; Young, S.L.; Miller, W.T.; Zhai, Y. The protist, Monosiga brevicollis, has a tyrosine kinase signaling network more elaborate and diverse than found in any known metazoan. Proc. Natl. Acad. Sci. USA 2008, 105, 9674–9679. [Google Scholar] [CrossRef] [Green Version]
- Brunet, T.; King, N. The Origin of Animal Multicellularity and Cell Differentiation. Dev. Cell 2017, 43, 124–140. [Google Scholar] [CrossRef]
- Ksionda, O.; Saveliev, A.; Kochl, R.; Rapley, J.; Faroudi, M.; Smith-Garvin, J.E.; Wulfing, C.; Rittinger, K.; Carter, T.; Tybulewicz, V.L. Mechanism and function of Vav1 localisation in TCR signalling. J. Cell Sci. 2012, 125, 5302–5314. [Google Scholar] [CrossRef] [Green Version]
- Flajnik, M.F.; Kasahara, M. Origin and evolution of the adaptive immune system: Genetic events and selective pressures. Nat. Rev. Genet. 2010, 11, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, C.; Li, Q.; Li, B.; Larkin, D.M.; Lee, C.; Storz, J.F.; Antunes, A.; Greenwold, M.J.; Meredith, R.W.; et al. Comparative genomics reveals insights into avian genome evolution and adaptation. Science 2014, 346, 1311–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prum, R.O.; Berv, J.S.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A comprehensive phylogeny of birds (Aves) using targeted next–generation DNA sequencing. Nature 2015, 526, 569–573. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Origin of animal multicellularity: Precursors, causes, consequences–the choanoflagellate/sponge transition, neurogenesis and the Cambrian explosion. Philos. Trans. R Soc. Lond B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef]
- Sebe-Pedros, A.; Burkhardt, P.; Sanchez-Pons, N.; Fairclough, S.R.; Lang, B.F.; King, N.; Ruiz-Trillo, I. Insights into the origin of metazoan filopodia and microvilli. Mol. Biol. Evol. 2013, 30, 2013–2023. [Google Scholar] [CrossRef]
- Govindan, J.A.; Cheng, H.; Harris, J.E.; Greenstein, D. Galphao/i and Galphas signaling function in parallel with the MSP/Eph receptor to control meiotic diapause in C. elegans. Curr. Biol. 2006, 16, 1257–1268. [Google Scholar] [CrossRef] [PubMed]
- Fry, A.L.; Laboy, J.T.; Norman, K.R. VAV–1 acts in a single interneuron to inhibit motor circuit activity in Caenorhabditis elegans. Nat. Commun. 2014, 5, 5579. [Google Scholar] [CrossRef] [PubMed]
- Fry, A.L.; Laboy, J.T.; Huang, H.; Hart, A.C.; Norman, K.R. A Conserved GEF for Rho–Family GTPases Acts in an EGF Signaling Pathway to Promote Sleep–like Quiescence in Caenorhabditis elegans. Genetics 2016, 202, 1153–1166. [Google Scholar] [CrossRef] [PubMed]
- Sauzeau, V.; Sevilla, M.A.; Rivas-Elena, J.V.; de Alava, E.; Montero, M.J.; Lopez-Novoa, J.M.; Bustelo, X.R. Vav3 proto–oncogene deficiency leads to sympathetic hyperactivity and cardiovascular dysfunction. Nat. Med. 2006, 12, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Quevedo, C.; Sauzeau, V.; Menacho-Marquez, M.; Castro-Castro, A.; Bustelo, X.R. Vav3–deficient mice exhibit a transient delay in cerebellar development. Mol. Biol. Celll 2010, 21, 1125–1139. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.W.; Shao, Y.R.; Sahin, M.; Shamah, S.M.; Lin, M.Z.; Greer, P.L.; Gao, S.; Griffith, E.C.; Brugge, J.S.; Greenberg, M.E. Vav family GEFs link activated Ephs to endocytosis and axon guidance. Neuron 2005, 46, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Sauzeau, V.; Horta-Junior, J.A.; Riolobos, A.S.; Fernandez, G.; Sevilla, M.A.; Lopez, D.E.; Montero, M.J.; Rico, B.; Bustelo, X.R. Vav3 is involved in GABAergic axon guidance events important for the proper function of brainstem neurons controlling cardiovascular, respiratory, and renal parameters. Mol. Biol. Cell 2010, 21, 4251–4263. [Google Scholar] [CrossRef]
- Malartre, M.; Ayaz, D.; Amador, F.F.; Martin-Bermudo, M.D. The guanine exchange factor vav controls axon growth and guidance during Drosophila development. J. Neurosci. 2010, 30, 2257–2267. [Google Scholar] [CrossRef]
- Fernandez-Espartero, C.H.; Ramel, D.; Farago, M.; Malartre, M.; Luque, C.M.; Limanovich, S.; Katzav, S.; Emery, G.; Martin-Bermudo, M.D. GTP exchange factor Vav regulates guided cell migration by coupling guidance receptor signalling to local Rac activation. J. Cell Sci. 2013, 126, 2285–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schardt, L.; Ander, J.J.; Lohmann, I.; Papagiannouli, F. Stage–specific control of niche positioning and integrity in the Drosophila testis. Mech. Dev. 2015, 138, 336–348. [Google Scholar] [CrossRef]
- Sarkar, A.; Parikh, N.; Hearn, S.A.; Fuller, M.T.; Tazuke, S.I.; Schulz, C. Antagonistic roles of Rac and Rho in organizing the germ cell microenvironment. Curr. Biol. 2007, 17, 1253–1258. [Google Scholar] [CrossRef]
- Martin-Bermudo, M.D.; Bardet, P.L.; Bellaiche, Y.; Malartre, M. The vav oncogene antagonises EGFR signalling and regulates adherens junction dynamics during Drosophila eye development. Development 2015, 142, 1492–1501. [Google Scholar] [CrossRef] [Green Version]
- Ruan, W.; Srinivasan, A.; Lin, S.; Kara k, I.; Barker, P.A. Eiger–induced cell death relies on Rac1–dependent endocytosis. Cell Death Dis. 2016, 7, e2181. [Google Scholar] [CrossRef]
- Doody, G.M.; Bell, S.E.; Vigorito, E.; Clayton, E.; McAdam, S.; Tooze, R.; Fernandez, C.; Lee, I.J.; Turner, M. Signal transduction through Vav–2 participates in humoral immune responses and B cell maturation. Nat. Immunol. 2001, 2, 542–547. [Google Scholar] [CrossRef]
- Sauzeau, V.; Jerkic, M.; Lopez-Novoa, J.M.; Bustelo, X.R. Loss of Vav2 proto–oncogene causes tachycardia and cardiovascular disease in mice. Mol. Biol. Cell 2007, 18, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Zhao, C.; Wu, Y.; Yang, Q.; Shao, A.; Wang, T.; Wu, J.; Yin, Y.; Li, Y.; Hou, J.; et al. Identification of a Vav2–dependent mechanism for GDNF/Ret control of mesolimbic DAT trafficking. Nat. Neurosci. 2015, 18, 1084–1093. [Google Scholar] [CrossRef]
- Guyenet, P.G. The sympathetic control of blood pressure. Nat. Rev. Neurosci. 2006, 7, 335–346. [Google Scholar] [CrossRef]
- Menacho-Marquez, M.; Nogueiras, R.; Fabbiano, S.; Sauzeau, V.; Al-Massadi, O.; Dieguez, C.; Bustelo, X.R. Chronic sympathoexcitation through loss of Vav3, a Rac1 activator, results in divergent effects on metabolic syndrome and obesity depending on diet. Cell Metab. 2013, 18, 199–211. [Google Scholar] [CrossRef]
- Lorenzo-Martin, L.F.; Menacho-Marquez, M.; Fabbiano, S.; Al-Massadi, O.; Abad, A.; Rodriguez-Fdez, S.; Sevilla, M.A.; Montero, M.J.; Dieguez, C.; Nogueiras, R.; et al. Vagal afferents contribute to sympathoexcitation–driven metabolic dysfunctions. J. Endocrinol. 2019, 240, 483–496. [Google Scholar] [CrossRef]
- Ulc, A.; Zeug, A.; Bauch, J.; van Leeuwen, S.; Kuhlmann, T.; Ffrench-Constant, C.; Ponimaskin, E.; Faissner, A. The guanine nucleotide exchange factor Vav3 modulates oligodendrocyte precursor differentiation and supports remyelination in white matter lesions. GLIA 2019, 67, 376–392. [Google Scholar] [CrossRef] [PubMed]
- Hilfenhaus, G.; Nguyen, D.P.; Freshman, J.; Prajapati, D.; Ma, F.; Song, D.; Ziyad, S.; Cuadrado, M.; Pellegrini, M.; Bustelo, X.R.; et al. Vav3–induced cytoskeletal dynamics contribute to heterotypic properties of endothelial barriers. J. Cell Biol. 2018, 217, 2813–2830. [Google Scholar] [CrossRef]
- Faccio, R.; Teitelbaum, S.L.; Fujikawa, K.; Chappel, J.; Zallone, A.; Tybulewicz, V.L.; Ross, F.P.; Swat, W. Vav3 regulates osteoclast function and bone mass. Nat. Med. 2005, 11, 284–290. [Google Scholar] [CrossRef]
- Luft, V.; Reinhard, J.; Shibuya, M.; Fischer, K.D.; Faissner, A. The guanine nucleotide exchange factor Vav3 regulates differentiation of progenitor cells in the developing mouse retina. Cell Tissue Res. 2015, 359, 423–440. [Google Scholar] [CrossRef]
- Turner, M.; Billadeau, D.D. VAV proteins as signal integrators for multi–subunit immune–recognition receptors. Nat. Rev. Immunol. 2002, 2, 476–486. [Google Scholar] [CrossRef]
- Fabbiano, S.; Menacho-Marquez, M.; Robles-Valero, J.; Pericacho, M.; Matesanz-Marin, A.; Garcia-Macias, C.; Sevilla, M.A.; Montero, M.J.; Alarcon, B.; Lopez-Novoa, J.M.; et al. Immunosuppression–Independent Role of Regulatory T Cells against Hypertension–Driven Renal Dysfunctions. Mol. Cell Biol. 2015, 35, 3528–3546. [Google Scholar] [CrossRef]
- Gaud, G.; Roncagalli, R.; Chaoui, K.; Bernard, I.; Familiades, J.; Colacios, C.; Kassem, S.; Monsarrat, B.; Burlet-Schiltz, O.; de Peredo, A.G.; et al. The costimulatory molecule CD226 signals through VAV1 to amplify TCR signals and promote IL–17 production by CD4(+) T cells. Sci. Signal 2018, 11. [Google Scholar] [CrossRef]
- Guerreiro-Cacais, A.O.; Norin, U.; Gyllenberg, A.; Berglund, R.; Beyeen, A.D.; Rheumatoid Arthritis Consortium International (RACI); Petit-Teixeira, E.; Cornelis, F.; Saoudi, A.; Fournie, G.J.; et al. VAV1 regulates experimental autoimmune arthritis and is associated with anti–CCP negative rheumatoid arthritis. Genes. Immun. 2017, 18, 109. [Google Scholar] [CrossRef] [PubMed]
- Hale, C.F.; Dietz, K.C.; Varela, J.A.; Wood, C.B.; Zirlin, B.C.; Leverich, L.S.; Greene, R.W.; Cowan, C.W. Essential role for vav Guanine nucleotide exchange factors in brain–derived neurotrophic factor–induced dendritic spine growth and synapse plasticity. J. Neurosci. 2011, 31, 12426–12436. [Google Scholar] [CrossRef]
- Hunter, S.G.; Zhuang, G.; Brantley-Sieders, D.; Swat, W.; Cowan, C.W.; Chen, J. Essential role of Vav family guanine nucleotide exchange factors in EphA receptor–mediated angiogenesis. Mol. Cell Biol. 2006, 26, 4830–4842. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Seno, H.; Miletic, A.V.; Mills, J.C.; Swat, W.; Stappenbeck, T.S. Vav proteins are necessary for correct differentiation of mouse cecal and colonic enterocytes. J. Cell Sci. 2009, 122, 324–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Fdez, S.; Bustelo, X.R. The Vav GEF Family: An Evolutionary and Functional Perspective. Cells 2019, 8, 465. https://doi.org/10.3390/cells8050465
Rodríguez-Fdez S, Bustelo XR. The Vav GEF Family: An Evolutionary and Functional Perspective. Cells. 2019; 8(5):465. https://doi.org/10.3390/cells8050465
Chicago/Turabian StyleRodríguez-Fdez, Sonia, and Xosé R. Bustelo. 2019. "The Vav GEF Family: An Evolutionary and Functional Perspective" Cells 8, no. 5: 465. https://doi.org/10.3390/cells8050465
APA StyleRodríguez-Fdez, S., & Bustelo, X. R. (2019). The Vav GEF Family: An Evolutionary and Functional Perspective. Cells, 8(5), 465. https://doi.org/10.3390/cells8050465