mTOR Signaling Pathway Regulates Sperm Quality in Older Men

 ,
,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Ethical Approval
2.2. Study Design
2.3. Semen Analysis and Processing
2.4. Protein Quantification
2.5. Antibody Array
2.6. Network and Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
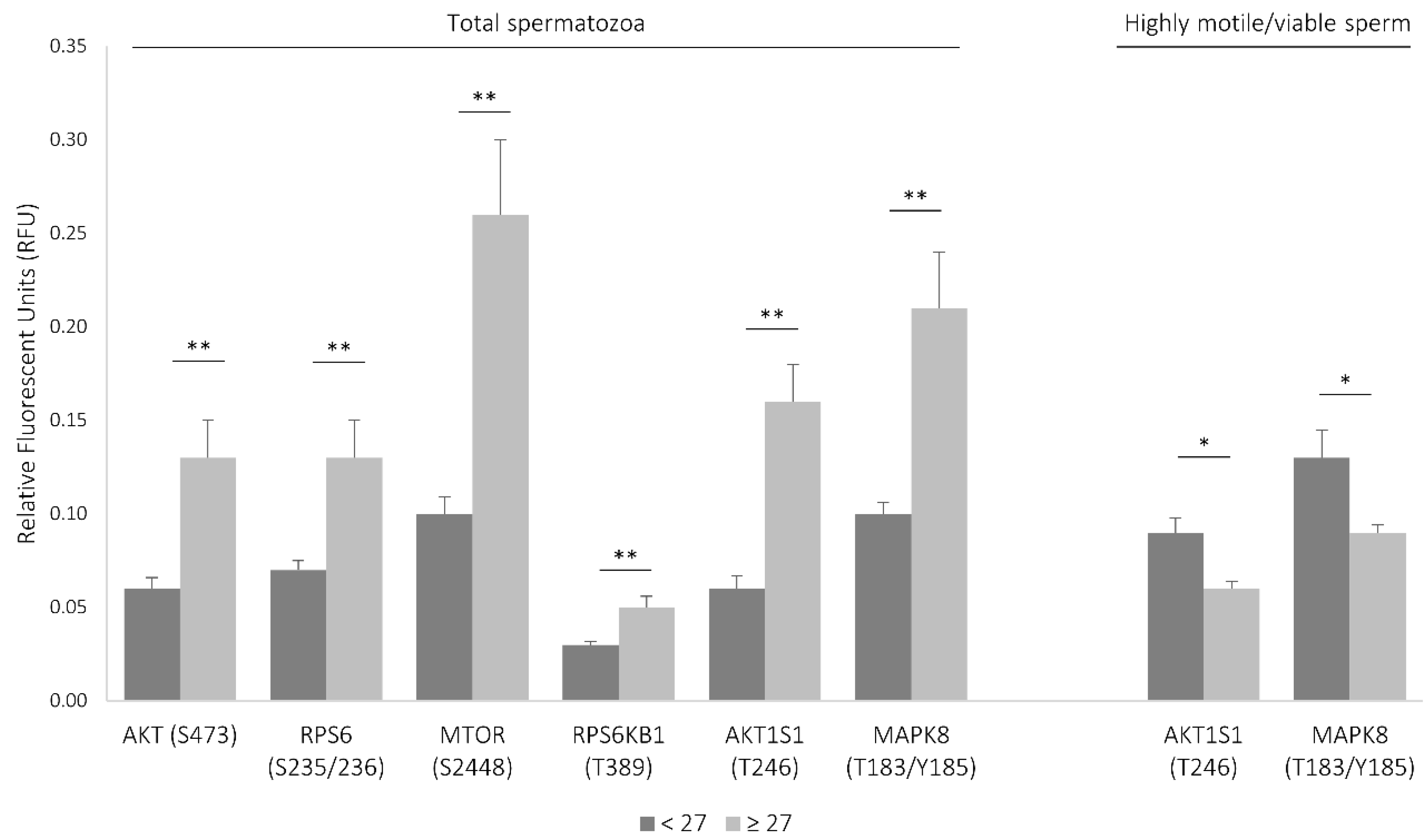
3.1. Age Impacts the Activity of Several Signaling Proteins
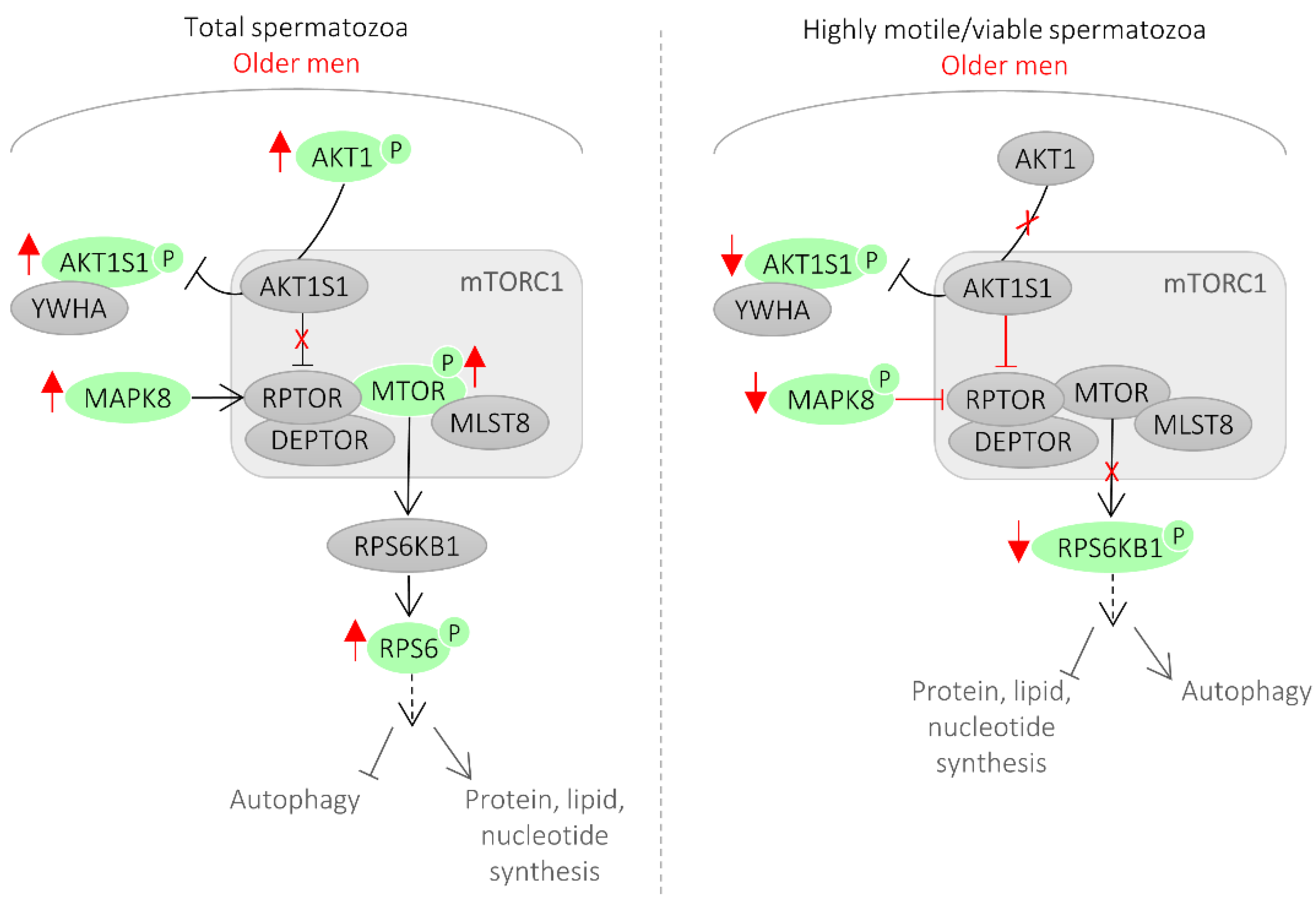
3.2. Network of Sperm Proteins Associated with Male Age
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lutz, W.; Leridon, H.; Aitken, R.J.; Von Eyben, F.E. Fertility rates and future population trends: Will Europe’s birth rate recover or continue to decline? Int. J. Androl. 2006, 29, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.W. Male infertility. Endocrinol. Metab. Clin. N. Am. 1994, 23, 783–793. [Google Scholar] [CrossRef]
- Hamada, A.; Esteves, S.C.; Nizza, M.; Agarwal, A. Unexplained male infertility: Diagnosis and management. Int. Braz. J. Urol. 2012, 38, 576–594. [Google Scholar] [CrossRef] [PubMed]
- Cedars, M.I. Childhood implications of parental aging. Fertil. Steril. 2015, 103, 1379–1380. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M.; Steiner, A.Z. Age-related infertility. Obstet. Gynecol. Clin. N. Am. 2015, 42, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.L.; Dunleavy, J.; Gemmell, N.J.; Nakagawa, S. Consistent age-dependent declines in human semen quality: A systematic review and meta-analysis. Ageing Res. Rev. 2015, 19, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Kidd, S.A.; Eskenazi, B.; Wyrobek, A.J. Effects of male age on semen quality and fertility: A review of the literature. Fertil. Steril. 2001, 75, 237–248. [Google Scholar] [CrossRef]
- Zhu, Q.-X.; Meads, C.; Lu, M.-L.; Wu, J.-Q.; Zhou, W.-J.; Gao, E.-S. Turning point of age for semen quality: A population-based study in Chinese men. Fertil. Steril. 2011, 96, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Stone, B.A.; Alex, A.; Werlin, L.B.; Marrs, R.P. Age thresholds for changes in semen parameters in men. Fertil. Steril. 2013, 100, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Age, the environment and our reproductive future: Bonking baby boomers and the future of sex. Reproduction 2014, 147, S1–S11. [Google Scholar] [CrossRef]
- Sartorius, G.A.; Nieschlag, E. Paternal age and reproduction. Hum. Reprod. Update 2010, 16, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, B. Reproductive functions of the ageing male. Hum. Reprod. Update 2004, 10, 327–339. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010; ISBN 9789241547789. [Google Scholar]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Alanis-Lobato, G.; Andrade-Navarro, M.A.; Schaefer, M.H. HIPPIE v2.0: Enhancing meaningfulness and reliability of protein–protein interaction networks. Nucleic Acids Res. 2017, 45, D408–D414. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Bravo, À.; Queralt-Rosinach, N.; Gutiérrez-Sacristán, A.; Deu-Pons, J.; Centeno, E.; García-García, J.; Sanz, F.; Furlong, L.I. DisGeNET: A comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017, 45, D833–D839. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.L.; Blake, J.A.; Kadin, J.A.; Richardson, J.E.; Bult, C.J. Mouse Genome Database Group. Mouse Genome Database (MGD)-2018: Knowledgebase for the laboratory mouse. Nucleic Acids Res. 2018, 46, D836–D842. [Google Scholar] [CrossRef]
- Liu, F.-J.; Liu, X.; Han, J.-L.; Wang, Y.-W.; Jin, S.-H.; Liu, X.-X.; Liu, J.; Wang, W.-T.; Wang, W.-J. Aged men share the sperm protein PATE1 defect with young asthenozoospermia patients. Hum. Reprod. 2015, 30, 861–869. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Kwak, D.; Choi, S.; Jeong, H.; Jang, J.-H.; Lee, Y.; Jeon, H.; Lee, M.N.; Noh, J.; Cho, K.; Yoo, J.S.; et al. Osmotic Stress Regulates Mammalian Target of Rapamycin (mTOR) Complex 1 via c-Jun N-terminal Kinase (JNK)-mediated Raptor Protein Phosphorylation. J. Biol. Chem. 2012, 287, 18398–18407. [Google Scholar] [CrossRef] [Green Version]
- Frey, J.W.; Jacobs, B.L.; Goodman, C.A.; Hornberger, T.A. A role for Raptor phosphorylation in the mechanical activation of mTOR signaling. Cell. Signal. 2014, 26, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.K.; Lamming, D.W. The Mechanistic Target of Rapamycin: The Grand ConducTOR of Metabolism and Aging. Cell Metab. 2016, 23, 990–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, V.D.; Antebi, A.; Bartke, A.; Barzilai, N.; Brown-Borg, H.M.; Caruso, C.; Curiel, T.J.; de Cabo, R.; Franceschi, C.; Gems, D.; et al. Interventions to Slow Aging in Humans: Are We Ready? Aging Cell 2015, 14, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Huang, L.; Geng, Y.; He, J.; Chen, X.; Xu, H.; Li, R.; Wang, Y.; Ding, Y.; Liu, X. Rapamycin inhibits spermatogenesis by changing the autophagy status through suppressing mechanistic target of rapamycin-p70S6 kinase in male rats. Mol. Med. Rep. 2017, 16, 4029–4037. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, I.M.; Espino, J.; Bejarano, I.; Gallardo-Soler, A.; Campo, M.L.; Salido, G.M.; Pariente, J.A.; Peña, F.J.; Tapia, J.A. Autophagy-related proteins are functionally active in human spermatozoa and may be involved in the regulation of cell survival and motility. Sci. Rep. 2016, 6, 33647. [Google Scholar] [CrossRef]
- Ip, C.K.M.; Wong, A.S.T. p70 S6 kinase and actin dynamics: A perspective. Spermatogenesis 2012, 2, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.; Cardoso, M.F.; Sousa, M.; Viana, P.; Gonçalves, A.; Silva, J.; Barros, A. Quantitative study of caspase-3 activity in semen and after swim-up preparation in relation to sperm quality. Hum. Reprod. 2005, 20, 1307–1313. [Google Scholar] [CrossRef] [Green Version]
- Roessner, C.; Paasch, U.; Kratzsch, J.; Glander, H.-J.; Grunewald, S. Sperm apoptosis signalling in diabetic men. Reprod. Biomed. Online 2012, 25, 292–299. [Google Scholar] [CrossRef]
- Chang, F.-W.; Sun, G.-H.; Cheng, Y.-Y.; Chen, I.-C.; Chien, H.-H.; Wu, G.-J. Effects of varicocele upon the expression of apoptosis-related proteins. Andrologia 2010, 42, 225–230. [Google Scholar] [CrossRef]
- Raimondo, S.; Aprea, G.; Cuomo, F.; De Filippo, S.; Gentile, T.; Guida, J. Quantitative evaluation of p53 as a new indicator of DNA damage in human spermatozoa. J. Hum. Reprod. Sci. 2014, 7, 212. [Google Scholar] [CrossRef]
- Almog, T.; Naor, Z. The role of Mitogen activated protein kinase (MAPK) in sperm functions. Mol. Cell. Endocrinol. 2010, 314, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Li, M.W.M.; Mruk, D.D.; Cheng, C.Y. Mitogen-activated protein kinases in male reproductive function. Trends Mol. Med. 2009, 15, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Almog, T.; Lazar, S.; Reiss, N.; Etkovitz, N.; Milch, E.; Rahamim, N.; Dobkin-Bekman, M.; Rotem, R.; Kalina, M.; Ramon, J.; et al. Identification of extracellular signal-regulated kinase 1/2 and p38 MAPK as regulators of human sperm motility and acrosome reaction and as predictors of poor spermatozoan quality. J. Biol. Chem. 2008, 283, 14479–14489. [Google Scholar] [CrossRef]
- Kostenko, S.; Moens, U. Heat shock protein 27 phosphorylation: Kinases, phosphatases, functions and pathology. Cell. Mol. Life Sci. 2009, 66, 3289–3307. [Google Scholar] [CrossRef]
- McReynolds, S.; Dzieciatkowska, M.; Stevens, J.; Hansen, K.C.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Toward the identification of a subset of unexplained infertility: A sperm proteomic approach. Fertil. Steril. 2014, 102, 692–699. [Google Scholar] [CrossRef]
- Intasqui, P.; Camargo, M.; Del Giudice, P.T.; Spaine, D.M.; Carvalho, V.M.; Cardozo, K.H.M.; Cedenho, A.P.; Bertolla, R.P. Unraveling the sperm proteome and post-genomic pathways associated with sperm nuclear DNA fragmentation. J. Assist. Reprod. Genet. 2013, 30, 1187–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqualotto, F.F.; Sobreiro, B.P.; Hallak, J.; Pasqualotto, E.B.; Lucon, A.M. Sperm concentration and normal sperm morphology decrease and follicle-stimulating hormone level increases with age. BJU Int. 2005, 96, 1087–1091. [Google Scholar] [CrossRef]
- Levitas, E.; Lunenfeld, E.; Weisz, N.; Friger, M.; Potashnik, G. Relationship between age and semen parameters in men with normal sperm concentration: Analysis of 6022 semen samples. Andrologia 2007, 39, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Kalyani, R.; Basavaraj, P.B.; Kumar, M.L.H. Factors influencing quality of semen: A two year prospective study. Indian J. Pathol. Microbiol. 2007, 50, 890–895. [Google Scholar]
- Cocuzza, M.; Athayde, K.S.; Agarwal, A.; Sharma, R.; Pagani, R.; Lucon, A.M.; Srougi, M.; Hallak, J. Age-Related Increase of Reactive Oxygen Species in Neat Semen in Healthy Fertile Men. Urology 2008, 71, 490–494. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Uniprot ID | Abbreviation | Protein Name | Phosphorylation/Cleavage, Residue, Status | Pearson’s Correlation Coefficient | |
|---|---|---|---|---|---|
| Formerly | Presently | ||||
| Total Spermatozoa Populations | |||||
| P04637 | p53 | TP53 | Cellular tumor antigen p53 | Phospho S15 Activation | 0.451 ** |
| Q92934 | Bad | BAD | Bcl2-associated agonist of cell death | Phospho S112 Inhibition | 0.434 ** |
| Q16539 | p38 | MAPK 11/12/13/14 | Mitogen-activated protein kinase 14/11/12/13 | Phospho T180/Y182 Activation | 0.430 ** |
| Q15759 | |||||
| P53778 | |||||
| O15264 | |||||
| P42345 | mTOR | MTOR | Serine/threonine-protein kinase mTOR | Phospho S2448 Activation | 0.409 ** |
| Q96B36 | PRAS40 | AKT1S1 | Proline-rich AKT1 substrate 1 | Phospho T246 Inhibition | 0.406 ** |
| P04792 | HSP27 | HSPB1 | Heat shock protein beta-1 | Phospho S78 Activation | 0.373 ** |
| P31749 | Akt | AKT | RAC-alpha serine/threonine-protein kinase | Phospho S473 Activation | 0.365 ** |
| P42574 | Caspase-3 | CASP3 | Caspase-3 | Cleavage D175 Activation | 0.362 ** |
| P49841 | GSK-3β | GSK3B | Glycogen synthase kinase-3 beta | Phospho S9 Inhibition | 0.354 ** |
| P45983 | SAPK/JNK | MAPK8 | Mitogen-activated protein kinase 8 | Phospho T183/Y185 Activation | 0.335 ** |
| P62753 | S6 ribosomal protein | RPS6 | 40S ribosomal protein S6 | Phospho S235/236 Activation | 0.317 ** |
| Highly Motile/Viable Spermatozoa | |||||
| Q96B36 | PRAS40 | AKT1S1 | Proline-rich AKT1 substrate 1 | Phospho T246 Inhibition | −0.523 ** |
| P23443 | p70 S6 kinase | RPS6KB1 | Ribosomal protein S6 kinase beta | Phospho T389 Activation | −0.382 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, J.V.; Cabral, M.; Correia, B.R.; Carvalho, P.; Sousa, M.; Oliveira, P.F.; Fardilha, M. mTOR Signaling Pathway Regulates Sperm Quality in Older Men. Cells 2019, 8, 629. https://doi.org/10.3390/cells8060629
Silva JV, Cabral M, Correia BR, Carvalho P, Sousa M, Oliveira PF, Fardilha M. mTOR Signaling Pathway Regulates Sperm Quality in Older Men. Cells. 2019; 8(6):629. https://doi.org/10.3390/cells8060629
Chicago/Turabian StyleSilva, Joana Vieira, Madalena Cabral, Bárbara Regadas Correia, Pedro Carvalho, Mário Sousa, Pedro Fontes Oliveira, and Margarida Fardilha. 2019. "mTOR Signaling Pathway Regulates Sperm Quality in Older Men" Cells 8, no. 6: 629. https://doi.org/10.3390/cells8060629
APA StyleSilva, J. V., Cabral, M., Correia, B. R., Carvalho, P., Sousa, M., Oliveira, P. F., & Fardilha, M. (2019). mTOR Signaling Pathway Regulates Sperm Quality in Older Men. Cells, 8(6), 629. https://doi.org/10.3390/cells8060629