Assessing the Epigenetic Status of Human Telomeres


Abstract
:1. The Epigenetic Nature of Human Telomeres Has Remained Controversial
2. Analysis of the Epigenetic Features of Human Telomeres by Microscopy
3. The Chromatin Immunoprecipitation Technique
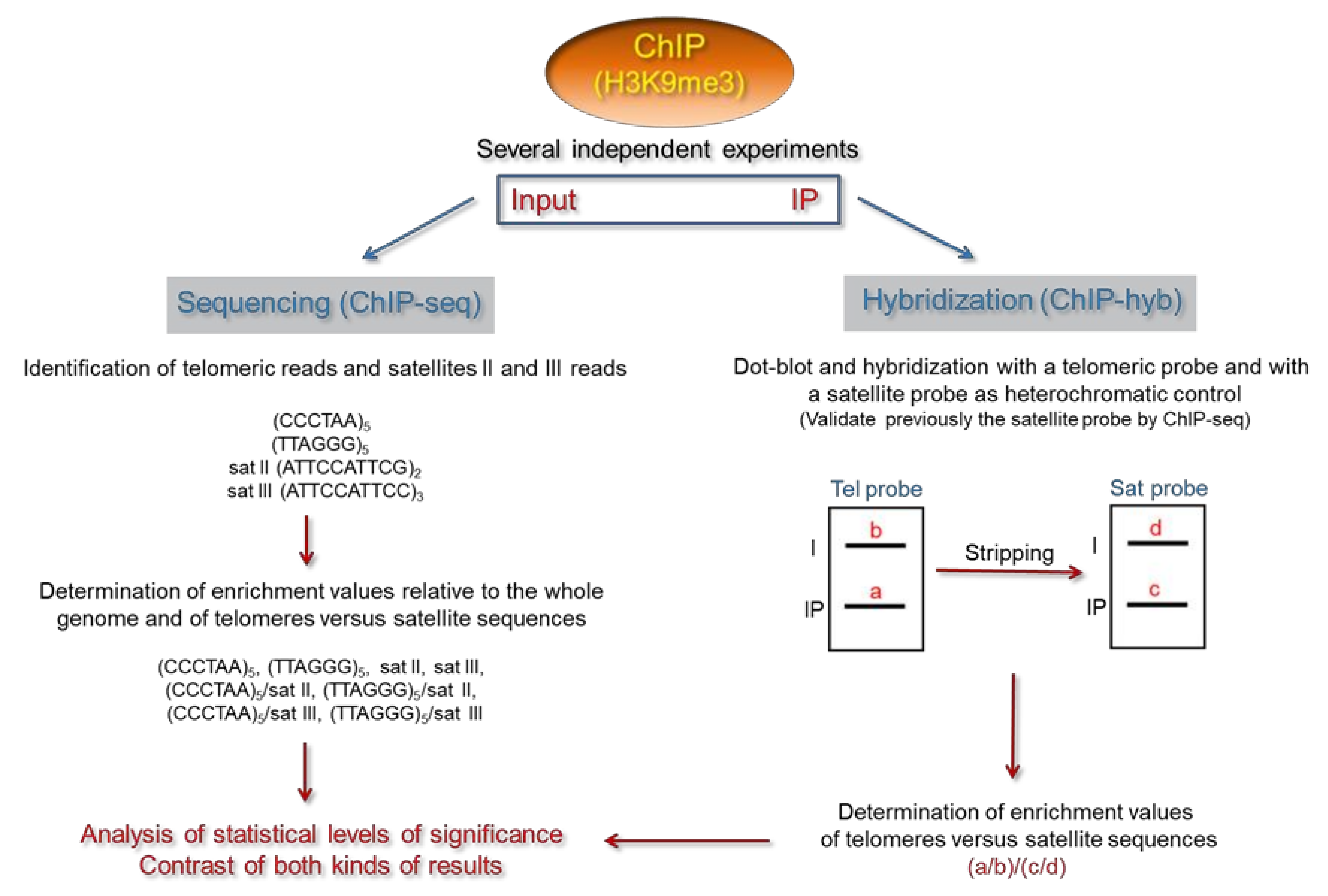
4. Analysis of the Epigenetic Features of Human Telomeres by ChIP-Hyb
5. Analysis of the Epigenetic Features of Human Telomeres by ChIP-Seq
6. Determining the Heterochromatic Status of Telomeres in Specific Human Cell Lines
7. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Blasco, M.A. The epigenetic regulation of mammalian telomeres. Nat. Rev. Genet. 2007, 8, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Connelly, K.E.; Dykhuizen, E.C. Compositional and functional diversity of canonical PRC1 complexes in mammals. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Holoch, D.; Margueron, R. Mechanisms Regulating PRC2 Recruitment and Enzymatic Activity. Trends Biochem. Sci. 2017, 42, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Ernst, J.; Kheradpour, P.; Mikkelsen, T.S.; Shoresh, N.; Ward, L.D.; Epstein, C.B.; Zhang, X.; Wang, L.; Issner, R.; Coyne, M.; et al. Mapping and analysis of chromatin state dynamics in nine human cell types. Nature 2011, 473, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Nishibuchi, G.; Dejardin, J. The molecular basis of the organization of repetitive DNA-containing constitutive heterochromatin in mammals. Chromosome Res. 2017, 25, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.J.; Cropley, J.E.; Pickett, H.A.; Reddel, R.R.; Suter, C.M. Telomerase activity is associated with an increase in DNA methylation at the proximal subtelomere and a reduction in telomeric transcription. Nucleic Acids Res. 2009, 37, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Brock, G.J.; Charlton, J.; Bird, A. Densely methylated sequences that are preferentially localized at telomere-proximal regions of human chromosomes. Gene 1999, 240, 269–277. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Suzuki, K.; Qu, J.; Wang, P.; Zhou, J.; Liu, X.; Ren, R.; Xu, X.; Ocampo, A.; et al. A Werner syndrome stem cell model unveils heterochromatin alterations as a driver of human aging. Science 2015, 348, 1160–1163. [Google Scholar] [CrossRef] [Green Version]
- Dejardin, J.; Kingston, R.E. Purification of proteins associated with specific genomic Loci. Cell 2009, 136, 175–186. [Google Scholar] [CrossRef]
- Saksouk, N.; Barth, T.K.; Ziegler-Birling, C.; Olova, N.; Nowak, A.; Rey, E.; Mateos-Langerak, J.; Urbach, S.; Reik, W.; Torres-Padilla, M.E.; et al. Redundant mechanisms to form silent chromatin at pericentromeric regions rely on BEND3 and DNA methylation. Mol. Cell 2014, 56, 580–594. [Google Scholar] [CrossRef]
- Grolimund, L.; Aeby, E.; Hamelin, R.; Armand, F.; Chiappe, D.; Moniatte, M.; Lingner, J. A quantitative telomeric chromatin isolation protocol identifies different telomeric states. Nat. Commun. 2013, 4, 2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frescas, D.; de Lange, T. Binding of TPP1 protein to TIN2 protein is required for POT1a,b protein-mediated telomere protection. J. Biol. Chem. 2014, 289, 24180–24187. [Google Scholar] [CrossRef] [PubMed]
- De Lange, T. Shelterin-mediated telomere protection. Annu. Rev. Genet. 2018, 52, 223–247. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, N.; Van Beneden, A.; Decottignies, A. Telomere length regulates TERRA levels through increased trimethylation of telomeric H3K9 and HP1alpha. Nat. Struct. Mol. Biol. 2012, 19, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Liu, L.; Sun, Y.; Xie, P.; Hu, L.; Yuan, D.; Chen, D.; Ouyang, Q.; Lin, G.; Lu, G. Telomerase-mediated telomere elongation from human blastocysts to embryonic stem cells. J. Cell Sci. 2014, 127, 752–762. [Google Scholar] [CrossRef]
- Stong, N.; Deng, Z.; Gupta, R.; Hu, S.; Paul, S.; Weiner, A.K.; Eichler, E.E.; Graves, T.; Fronick, C.C.; Courtney, L.; et al. Subtelomeric CTCF and cohesin binding site organization using improved subtelomere assemblies and a novel annotation pipeline. Genome Res. 2014, 24, 1039–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samassekou, O.; Gadji, M.; Drouin, R.; Yan, J. Sizing the ends: Normal length of human telomeres. Ann. Anat. 2010, 192, 284–291. [Google Scholar] [CrossRef]
- Braunstein, M.; Rose, A.B.; Holmes, S.G.; Allis, C.D.; Broach, J.R. Transcriptional silencing in yeast is associated with reduced nucleosome acetylation. Genes Dev. 1993, 7, 592–604. [Google Scholar] [CrossRef]
- Hecht, A.; Strahl-Bolsinger, S.; Grunstein, M. Spreading of transcriptional repressor SIR3 from telomeric heterochromatin. Nature 1996, 383, 92–96. [Google Scholar] [CrossRef]
- Orlando, V.; Paro, R. Mapping Polycomb-repressed domains in the bithorax complex using in vivo formaldehyde cross-linked chromatin. Cell 1993, 75, 1187–1198. [Google Scholar] [CrossRef]
- Vaquero-Sedas, M.I.; Gamez-Arjona, F.M.; Vega-Palas, M.A. Arabidopsis thaliana telomeres exhibit euchromatic features. Nucleic Acids Res. 2011, 39, 2007–2017. [Google Scholar] [CrossRef] [PubMed]
- Vaquero-Sedas, M.I.; Luo, C.; Vega-Palas, M.A. Analysis of the epigenetic status of telomeres by using ChIP-seq data. Nucleic Acids Res. 2012, 40, e163. [Google Scholar] [CrossRef] [PubMed]
- Vaquero-Sedas, M.I.; Vega-Palas, M.A. Differential association of Arabidopsis telomeres and centromeres with histone H3 variants. Sci. Rep. 2013, 3, 1202. [Google Scholar] [CrossRef] [PubMed]
- Cubiles, M.; Barroso, S.; Enguix, A.; Vaquero-Sedas, M.I.; Aguilera, A.; Vega-Palas, M.A. Epigenetic features of human telomeres. Nucleic Acids Res. 2018, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gámez-Arjona, F.M.; López-López, C.; Vaquero-Sedas, M.I.; Vega-Palas, M.A. On the organization of the nucleosomes associated with telomeric sequences. Biochim. Biophys. Acta 2010, 1803, 1058–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaquero-Sedas, M.I.; Vega-Palas, M.A. On the chromatin structure of eukaryotic telomeres. Epigenetics 2011, 6, 1055–1058. [Google Scholar] [CrossRef] [Green Version]
- Fontenete, S.; Barros, J.; Madureira, P.; Figueiredo, C.; Wengel, J.; Azevedo, N.F. Mismatch discrimination in fluorescent in situ hybridization using different types of nucleic acids. Appl. Microbiol. Biotechnol. 2015, 99, 3961–3969. [Google Scholar] [CrossRef] [PubMed]
- González-García, M.P.; Pavelescu, I.; Canela, A.; Sevillano, X.; Leehy, K.A.; Nelson, A.D.; Ibanes, M.; Shippen, D.E.; Blasco, M.A.; Cano-Delgado, A.I. Single-cell telomere-length quantification couples telomere length to meristem activity and stem cell development in Arabidopsis. Cell Rep. 2015, 11, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.J.; Franklin, F.C.; Jones, G.H. Nucleolus-associated telomere clustering and pairing precede meiotic chromosome synapsis in Arabidopsis thaliana. J. Cell Sci. 2001, 114, 4207–4217. [Google Scholar]
- Armstrong, S.J.; Jones, G.H. Meiotic cytology and chromosome behaviour in wild-type Arabidopsis thaliana. J. Exp. Bot. 2003, 54, 1–10. [Google Scholar] [CrossRef]
- Tommerup, H.; Dousmanis, A.; de Lange, T. Unusual chromatin in human telomeres. Mol. Cell. Biol. 1994, 14, 5777–5785. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, Y.; Nishimura, Y.; Kurumizaka, H.; Shimizu, M. Nucleosome organization and chromatin dynamics in telomeres. Biomol. Concepts 2015, 6, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Pisano, S.; Galati, A.; Cacchione, S. Telomeric nucleosomes: Forgotten players at chromosome ends. Cell. Mol. Life Sci. 2008, 65, 3553–3563. [Google Scholar] [CrossRef] [PubMed]
- Negishi, Y.; Kawaji, H.; Minoda, A.; Usui, K. Identification of chromatin marks at TERRA promoter and encoding region. Biochem. Biophys. Res. Commun. 2015, 467, 1052–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schueler, M.G.; Sullivan, B.A. Structural and functional dynamics of human centromeric chromatin. Annu. Rev. Genomics Hum. Genet. 2006, 7, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.L.; Yewdell, W.T.; Bell, J.C.; McNulty, S.M.; Duda, Z.; O’Neill, R.J.; Sullivan, B.A.; Straight, A.F. RNA-dependent stabilization of SUV39H1 at constitutive heterochromatin. Elife 2017, 6, e25299. [Google Scholar] [CrossRef]
- Capurso, D.; Xiong, H.; Segal, M.R. A histone arginine methylation localizes to nucleosomes in satellite II and III DNA sequences in the human genome. BMC Genomics 2012, 13, 630. [Google Scholar] [CrossRef]
- Fanelli, M.; Caprodossi, S.; Ricci-Vitiani, L.; Porcellini, A.; Tomassoni-Ardori, F.; Amatori, S.; Andreoni, F.; Magnani, M.; De Maria, R.; Santoni, A.; et al. Loss of pericentromeric DNA methylation pattern in human glioblastoma is associated with altered DNA methyltransferases expression and involves the stem cell compartment. Oncogene 2008, 27, 358–365. [Google Scholar] [CrossRef]
- McNulty, S.M.; Sullivan, B.A. Alpha satellite DNA biology: Finding function in the recesses of the genome. Chromosome Res. 2018, 26, 115–138. [Google Scholar] [CrossRef]
- Waye, J.S.; Durfy, S.J.; Pinkel, D.; Kenwrick, S.; Patterson, M.; Davies, K.E.; Willard, H.F. Chromosome-specific alpha satellite DNA from human chromosome 1: Hierarchical structure and genomic organization of a polymorphic domain spanning several hundred kilobase pairs of centromeric DNA. Genomics 1987, 1, 43–51. [Google Scholar] [CrossRef]
- Olins, A.L.; Ishaque, N.; Chotewutmontri, S.; Langowski, J.; Olins, D.E. Retrotransposon Alu is enriched in the epichromatin of HL-60 cells. Nucleus 2014, 5, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, J.A.; Wang, Z.; Schones, D.E.; Zhao, K.; DeSalle, R.; Zhang, M.Q. Determination of enriched histone modifications in non-genic portions of the human genome. BMC Genomics 2009, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Vaquero-Sedas, M.I.; Vega-Palas, M.A. Targeting cancer through the epigenetic features of telomeric regions. Trends Cell Biol. 2019, 29, 281–290. [Google Scholar] [CrossRef] [PubMed]
- O′Sullivan, R.J.; Almouzni, G. Assembly of telomeric chromatin to create ALTernative endings. Trends Cell Biol. 2014, 24, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Gauchier, M.; Kan, S.; Barral, A.; Sauzet, S.; Agirre, E.; Bonnell, E.; Saksouk, N.; Barth, T.K.; Ide, S.; Urbach, S.; et al. SETDB1-dependent heterochromatin stimulates alternative lengthening of telomeres. Sci. Adv. 2019, 5, eaav3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero, J.J.; Lopez-Silanes, I.; Megias, D.; Fraga, M.F.; Castells-Garcia, A.; Blasco, M.A. TERRA recruitment of polycomb to telomeres is essential for histone trymethylation marks at telomeric heterochromatin. Nat. Commun. 2018, 9, 1548. [Google Scholar] [CrossRef]
- Blackburn, E.H. Telomeres and telomerase: The means to the end (Nobel lecture). Angew. Chem. Int. Ed. Engl. 2010, 49, 7405–7421. [Google Scholar] [CrossRef]
- Loo Yau, H.; Ettayebi, I.; De Carvalho, D.D. The Cancer Epigenome: Exploiting Its Vulnerabilities for Immunotherapy. Trends Cell Biol. 2019, 29, 31–43. [Google Scholar] [CrossRef]
- Pompili, L.; Leonetti, C.; Biroccio, A.; Salvati, E. Diagnosis and treatment of ALT tumors: Is Trabectedin a new therapeutic option? J. Exp. Clin. Cancer Res. 2017, 36, 189. [Google Scholar] [CrossRef]
- Tumini, E.; Herrera-Moyano, E.; San Martin-Alonso, M.; Barroso, S.; Galmarini, C.M.; Aguilera, A. The antitumor drugs trabectedin and lurbinectedin induce transcription-dependent replication stress and genome instability. Mol. Cancer Res. 2019, 17, 773–782. [Google Scholar] [CrossRef]
- Sagie, S.; Toubiana, S.; Hartono, S.R.; Katzir, H.; Tzur-Gilat, A.; Havazelet, S.; Francastel, C.; Velasco, G.; Chedin, F.; Selig, S. Telomeres in ICF syndrome cells are vulnerable to DNA damage due to elevated DNA:RNA hybrids. Nat. Commun. 2017, 8, 14015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, R.; Lee, Y.; Wischnewski, H.; Brun, C.M.; Schwarz, T.; Azzalin, C.M. RNaseH1 regulates TERRA-telomeric DNA hybrids and telomere maintenance in ALT tumour cells. Nat. Commun. 2014, 5, 5220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Pereira, J.M.; Aguilera, A. R loops: New modulators of genome dynamics and function. Nat. Rev. Genet. 2015, 16, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA binding to TRF2 facilitates heterochromatin formation and ORC recruitment at telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Graf, M.; Bonetti, D.; Lockhart, A.; Serhal, K.; Kellner, V.; Maicher, A.; Jolivet, P.; Teixeira, M.T.; Luke, B. Telomere Length Determines TERRA and R-Loop Regulation through the Cell Cycle. Cell 2017, 170, 72–85.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balk, B.; Maicher, A.; Dees, M.; Klermund, J.; Luke-Glaser, S.; Bender, K.; Luke, B. Telomeric RNA-DNA hybrids affect telomere-length dynamics and senescence. Nat. Struct. Mol. Biol. 2013, 20, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- O′Sullivan, R.J.; Kubicek, S.; Schreiber, S.L.; Karlseder, J. Reduced histone biosynthesis and chromatin changes arising from a damage signal at telomeres. Nat. Struct. Mol. Biol. 2010, 17, 1218–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Johnson, L.M.; Jacobsen, S.E.; Patel, D.J. DNA methylation pathways and their crosstalk with histone methylation. Nat. Rev. Mol. Cell. Biol. 2015, 16, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Vega-Vaquero, A.; Bonora, G.; Morselli, M.; Vaquero-Sedas, M.I.; Rubbi, L.; Pellegrini, M.; Vega-Palas, M.A. Novel features of telomere biology revealed by the absence of telomeric DNA methylation. Genome Res. 2016, 26, 1047–1056. [Google Scholar] [CrossRef] [Green Version]
- Vancevska, A.; Douglass, K.M.; Pfeiffer, V.; Manley, S.; Lingner, J. The telomeric DNA damage response occurs in the absence of chromatin decompaction. Genes Dev. 2017, 31, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Timashev, L.A.; Babcock, H.; Zhuang, X.; de Lange, T. The DDR at telomeres lacking intact shelterin does not require substantial chromatin decompaction. Genes Dev. 2017, 31, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Bandaria, J.N.; Qin, P.; Berk, V.; Chu, S.; Yildiz, A. Shelterin protects chromosome ends by compacting telomeric chromatin. Cell 2016, 164, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Makarov, V.L.; Lejnine, S.; Bedoyan, J.; Langmore, J.P. Nucleosomal organization of telomere-specific chromatin in rat. Cell 1993, 73, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Xu, L.; Crawford, G.; Wang, Z.; Burgess, S.M. The forkhead transcription factor FoxI1 remains bound to condensed mitotic chromosomes and stably remodels chromatin structure. Mol. Cell. Biol. 2006, 26, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Hyle, J.; Lechner, M.S.; Lahti, J.M. Mammalian ChlR1 has a role in heterochromatin organization. Exp. Cell Res. 2011, 317, 2522–2535. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Evidence | Ref. | |
|---|---|---|
| 1 | Immunofluorescence microscopy studies reveal low co-localization levels of the H3K9me3 or the HP1 heterochromatic marks with telomeres, which could be explained by the presence of H3K9me3 and HP1 in subtelomeric heterochromatin. | [8,14,15,16] |
| 2 | ChIP-hyb experiments reveal low levels of H3K9me3 at telomeric repeats as compared with subtelomeric sequences. | [57] |
| 3 | ChIP-seq experiments show that telomeres are not enriched in H3K9me3 but have enhanced levels of the H3K27ac and H4K20me1 euchromatic marks. In contrast, satellites II and III are enriched in H3K9me3 but not in H3K27ac or H4K20me1. | [24,42] |
| 4 | Human telomeres should not be methylated because the human telomeric sequence is TTAGGG, and DNA methylation in mammals mainly circumscribe to the CG context. In agreement with this assumption, DNA methylation is completely absent from Arabidopsis telomeres. However, the Arabidopsis telomeric sequence, which is TTTAGGG, is methylated by the CHH methylation machinery (where H is A, C, or T) when it is present in interstitial telomeric sequences (ITSs). | [58,59] |
| 5 | Super-resolution microscopy shows that the disruption of shelterin proteins can lead to an increase in telomeres volume that has been related to telomeres decompaction and to a 53BP1-dependent telomere clustering. However, telomeres volume is not affected in human cells treated with inhibitors of heterochromatin formation. Therefore, heterochromatin does not seem to affect the protective structure of telomeres as shelterins do. | [60,61,62] |
| 6 | Telomeric nucleosomes are shorter and more sensitive to overall micrococcal nuclease digestion than the heterochromatic nucleosomes associated with chromocenters. | [31,63,64,65] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaquero-Sedas, M.I.; Vega-Palas, M.A. Assessing the Epigenetic Status of Human Telomeres. Cells 2019, 8, 1050. https://doi.org/10.3390/cells8091050
Vaquero-Sedas MI, Vega-Palas MA. Assessing the Epigenetic Status of Human Telomeres. Cells. 2019; 8(9):1050. https://doi.org/10.3390/cells8091050
Chicago/Turabian StyleVaquero-Sedas, María I., and Miguel A. Vega-Palas. 2019. "Assessing the Epigenetic Status of Human Telomeres" Cells 8, no. 9: 1050. https://doi.org/10.3390/cells8091050
APA StyleVaquero-Sedas, M. I., & Vega-Palas, M. A. (2019). Assessing the Epigenetic Status of Human Telomeres. Cells, 8(9), 1050. https://doi.org/10.3390/cells8091050