Delineating the Molecular Basis of the Calmodulin–bMunc13-2 Interaction by Cross-Linking/Mass Spectrometry—Evidence for a Novel CaM Binding Motif in bMunc13-2

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Expression and Purification of Recombinant CaM and bMunc13-2 Segments
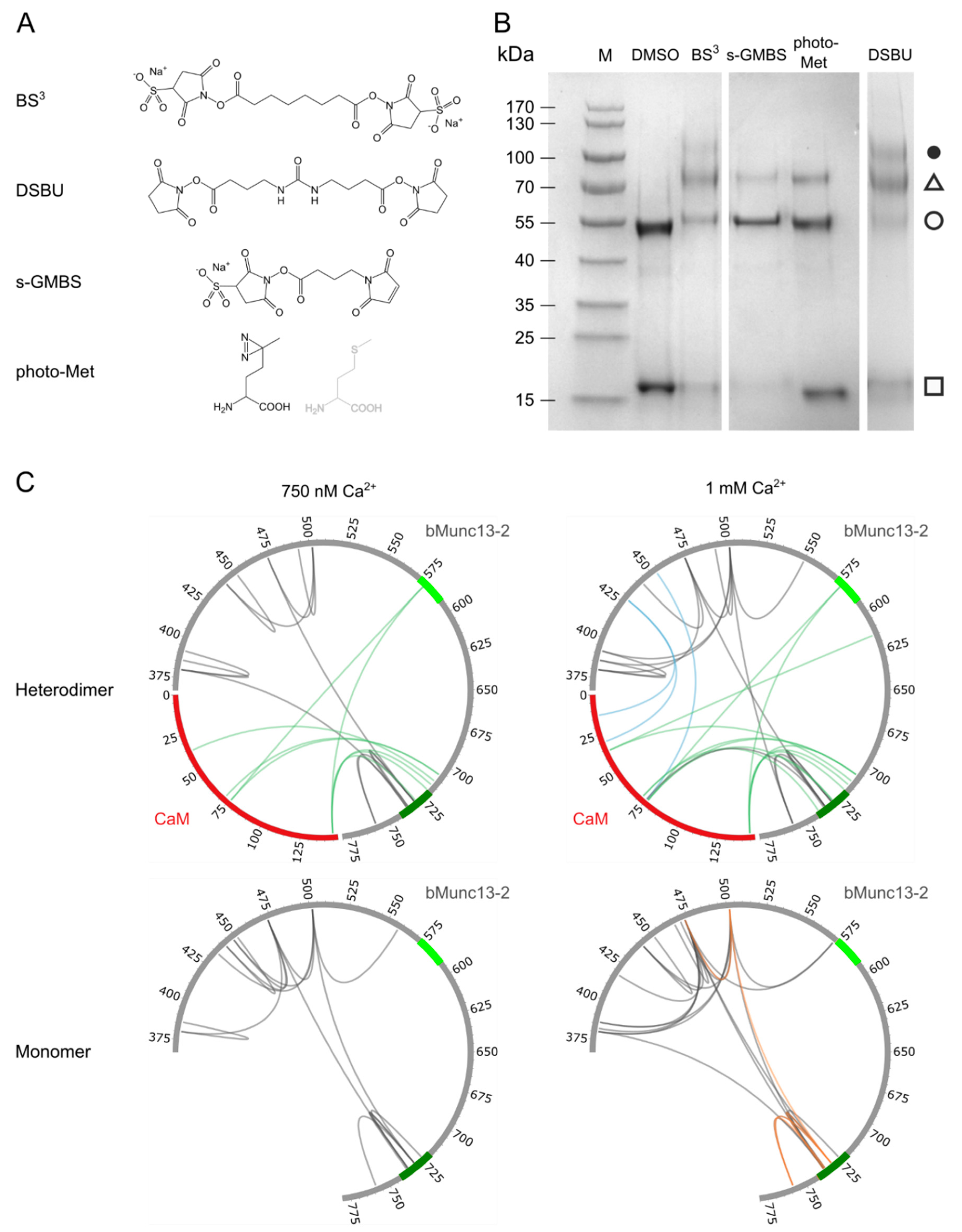
2.3. (Photo-)Chemical Cross-Linking
2.4. Enzymatic Digestion
2.5. Liquid Chromatography/Mass Spectrometry (LC/MS)
2.6. Identification of Cross-Links
2.7. Docking of CaM/bMunc13-2 Segment-C Complexes
2.8. Native Electrospray Ionization Mass Spectrometry
2.9. Isothermal Titration Calorimetry
2.10. Photoaffinity Labeling-Based Competition Assay
2.11. Surface Plasmon Resonance Spectroscopy
3. Results
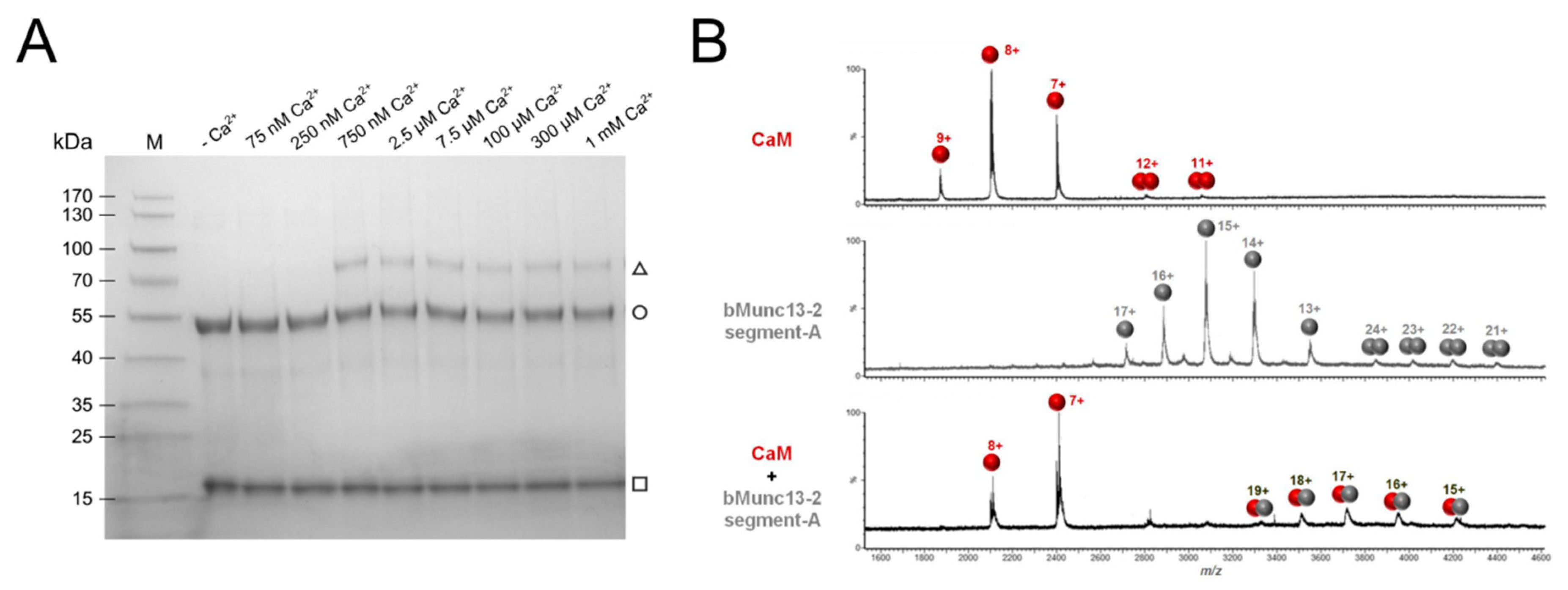
3.1. Ca2+-Dependent Formation of a Stoichiometric Complex between CaM and bMunc13-2
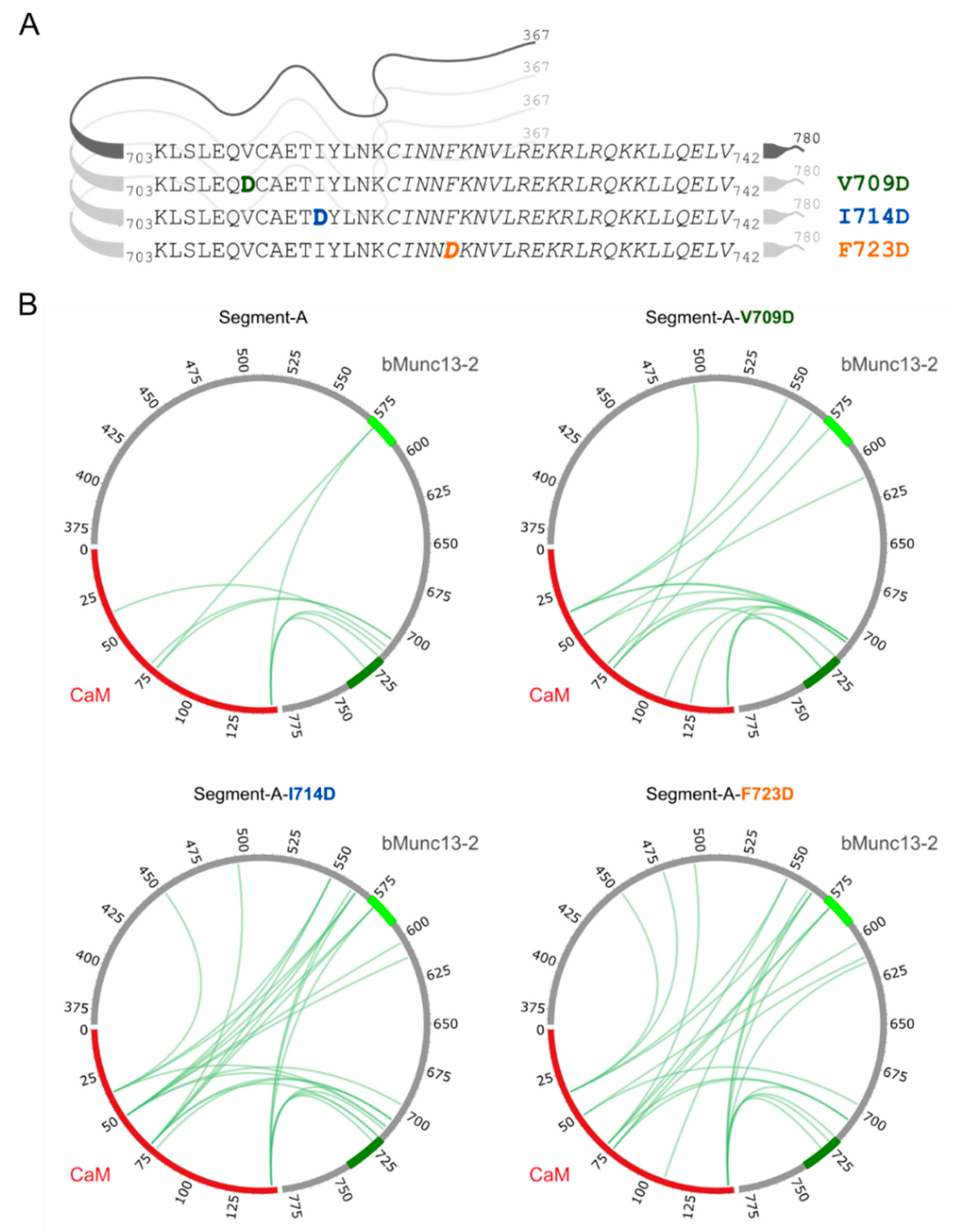
3.2. Structural Characterization of CaM–bMunc13-2 Segment-A Interaction by XL-MS
3.3. Inclusion of the C1 Domain of bMunc13-2 Does Not Influence CaM Binding
3.4. The CaM Binding Site in bMunc13-2 Is N-Terminally Elongated
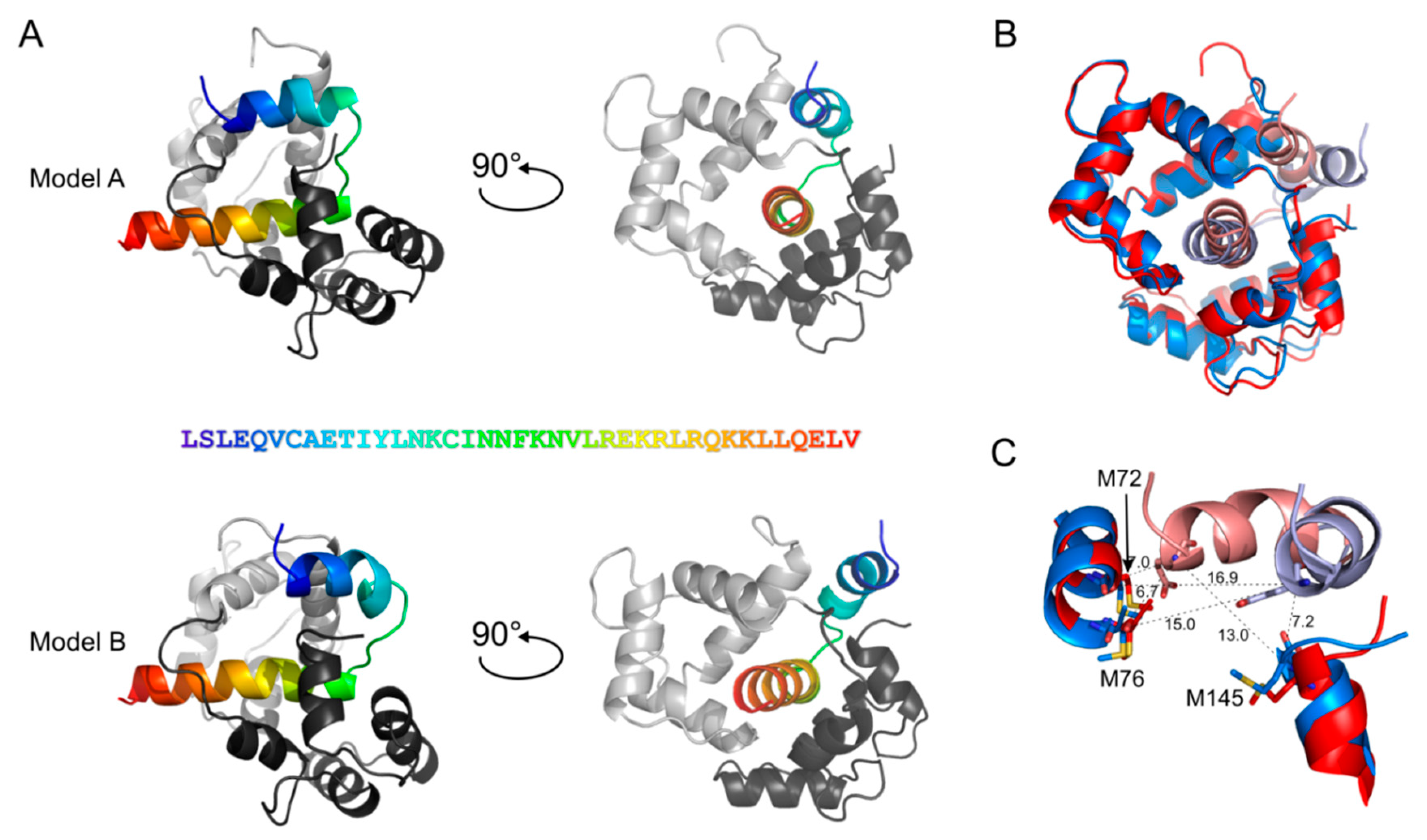
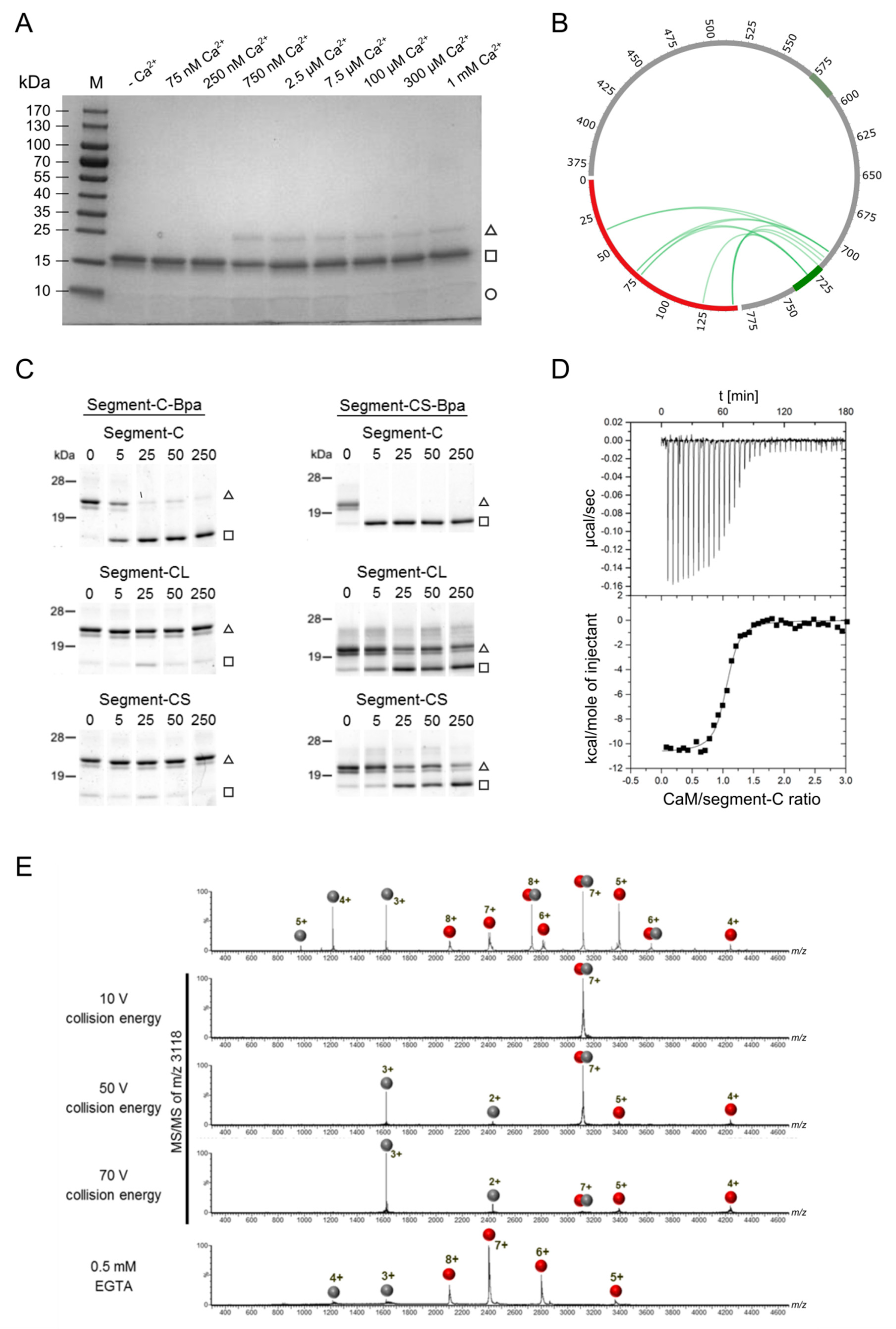
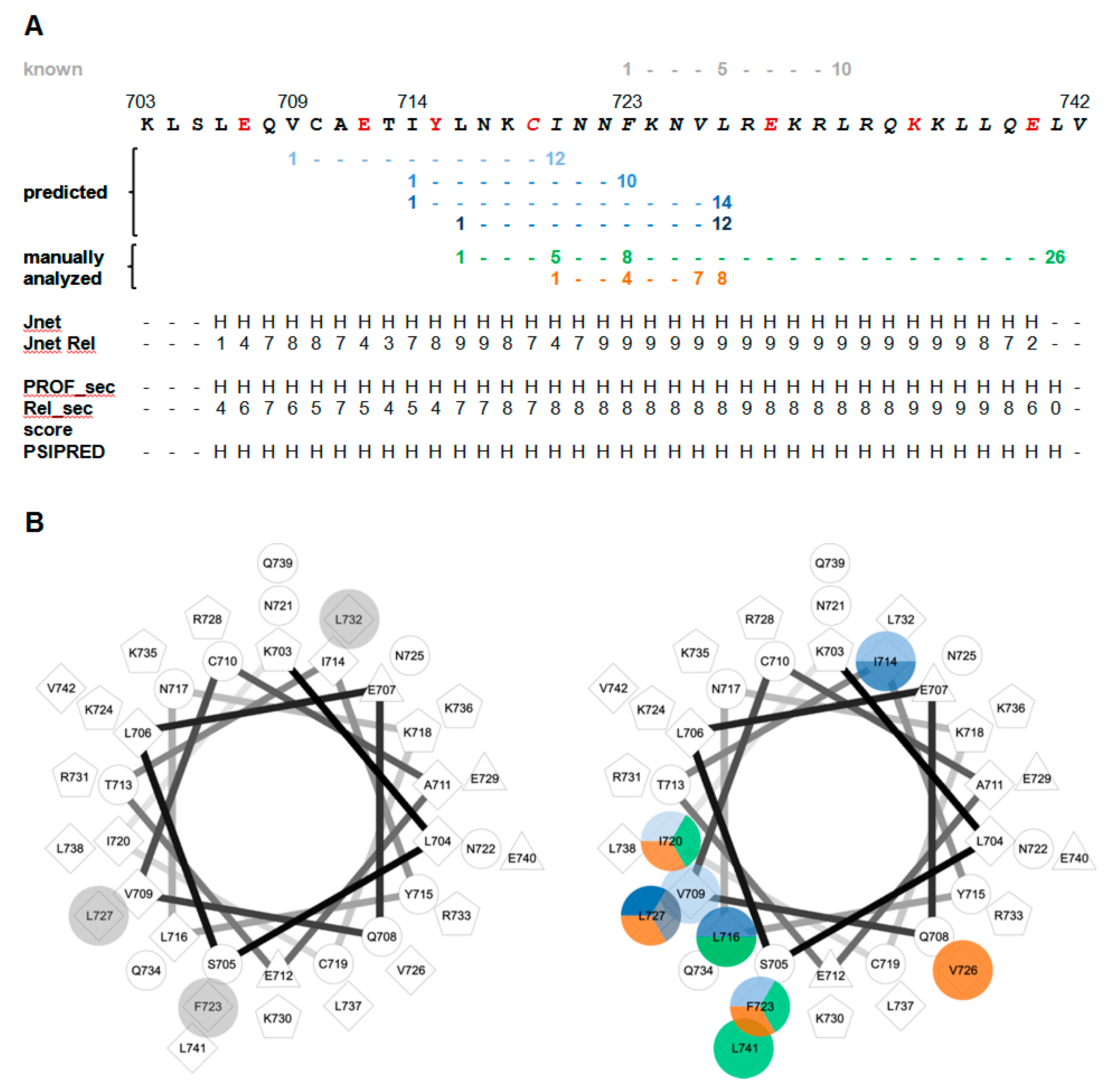
3.5. Detailed Analysis of the N-Terminally Elongated Binding Site
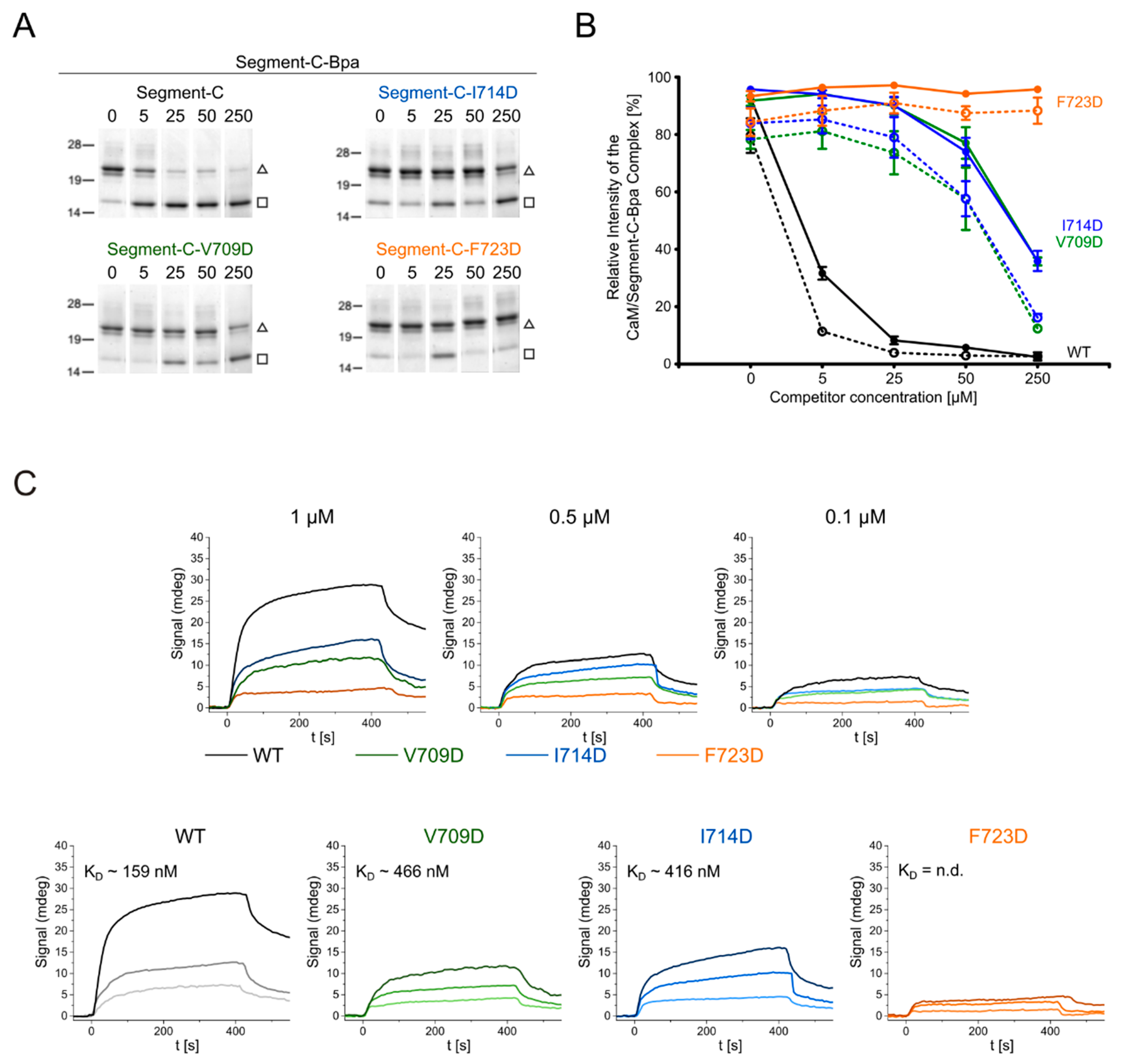
3.6. Hydrophobic Contact Sites in the N-Terminal Part of the bMunc13-2 CaM Binding Sequence Are Crucial for High-Affinity Binding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carafoli, E.; Krebs, J. Why Calcium? How Calcium Became the Best Communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berchtold, M.W.; Villalobo, A. The many faces of calmodulin in cell proliferation, programmed cell death, autophagy, and cancer. Biochim. Et Biophys. Acta 2014, 1843, 398–435. [Google Scholar] [CrossRef] [PubMed]
- Chin, D.; Means, A.R. Calmodulin: A prototypical calcium sensor. Trends Cell Biol. 2000, 10, 322–328. [Google Scholar] [CrossRef]
- Davis, T.N.; Thorner, J. Vertebrate and yeast calmodulin, despite significant sequence divergence, are functionally interchangeable. Proc. Natl. Acad. Sci. USA 1989, 86, 7909–7913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, Y.S.; Bugg, C.E.; Cook, W.J. Structure of calmodulin refined at 2.2 A resolution. J. Mol. Biol. 1988, 204, 191–204. [Google Scholar] [CrossRef]
- Fallon, J.L.; Quiocho, F.A. A closed compact structure of native Ca2+-calmodulin. Structure 2003, 11, 1303–1307. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Tanaka, T.; Ikura, M. Calcium-Induced Conformational Transition Revealed by the Solution Structure of Apo Calmodulin. Nat. Struct. Biol. 1995, 2, 758–767. [Google Scholar] [CrossRef]
- Shen, X.; Valencia, C.A.; Szostak, J.W.; Dong, B.; Liu, R. Scanning the human proteome for calmodulin-binding proteins. Proc. Natl. Acad. Sci. USA 2005, 102, 5969–5974. [Google Scholar] [CrossRef] [Green Version]
- Villarroel, A.; Taglialatela, M.; Bernardo-Seisdedos, G.; Alaimo, A.; Agirre, J.; Alberdi, A.; Gomis-Perez, C.; Soldovieri, M.V.; Ambrosino, P.; Malo, C.; et al. The ever changing moods of calmodulin: How structural plasticity entails transductional adaptability. J. Mol. Biol. 2014, 426, 2717–2735. [Google Scholar] [CrossRef]
- Tidow, H.; Nissen, P. Structural diversity of calmodulin binding to its target sites. FEBS J. 2013, 280, 5551–5565. [Google Scholar] [CrossRef]
- Rhoads, A.R.; Friedberg, F. Sequence motifs for calmodulin recognition. FASEB J. 1997, 11, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Vetter, S.W.; Leclerc, E. Novel aspects of calmodulin target recognition and activation. Eur. J. Biochem. 2003, 270, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Bähler, M.; Rhoads, A. Calmodulin signaling via the IQ motif. FEBS Lett. 2002, 513, 107–113. [Google Scholar] [CrossRef]
- Mruk, K.; Farley, B.M.; Ritacco, A.W.; Kobertz, W.R. Calmodulation meta-analysis: Predicting calmodulin binding via canonical motif clustering. J. Gen. Physiol. 2014, 144, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Yap, K.L.; Kim, J.; Truong, K.; Sherman, M.; Yuan, T.; Ikura, M. Calmodulin target database. J. Struct. Funct. Genom. 2000, 1, 8–14. [Google Scholar] [CrossRef]
- Lipstein, N.; Goth, M.; Piotrowski, C.; Pagel, K.; Sinz, A.; Jahn, O. Presynaptic Calmodulin targets: Lessons from structural proteomics. Expert Rev. Proteom. 2017, 14, 223–242. [Google Scholar] [CrossRef]
- Ma, C.; Su, L.; Seven, A.B.; Xu, Y.; Rizo, J. Reconstitution of the vital functions of Munc18 and Munc13 in neurotransmitter release. Science 2013, 339, 421–425. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Choi, U.B.; Leitz, J.; Rhee, H.J.; Lee, C.; Altas, B.; Zhao, M.; Pfuetzner, R.A.; Wang, A.L.; Brose, N.; et al. Molecular Mechanisms of Synaptic Vesicle Priming by Munc13 and Munc18. Neuron 2017, 95, 591–607. [Google Scholar] [CrossRef]
- Imig, C.; Min, S.W.; Krinner, S.; Arancillo, M.; Rosenmund, C.; Sudhof, T.C.; Rhee, J.; Brose, N.; Cooper, B.H. The morphological and molecular nature of synaptic vesicle priming at presynaptic active zones. Neuron 2014, 84, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Brose, N.; Hofmann, K.; Hata, Y.; Sudhof, T.C. Mammalian homologues of Caenorhabditis elegans unc-13 gene define novel family of C2-domain proteins. J. Biol. Chem. 1995, 270, 25273–25280. [Google Scholar] [CrossRef] [Green Version]
- Koch, H.; Hofmann, K.; Brose, N. Definition of Munc13-homology-domains and characterization of a novel ubiquitously expressed Munc13 isoform. Biochem. J. 2000, 349, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Augustin, I.; Betz, A.; Herrmann, C.; Jo, T.; Brose, N. Differential expression of two novel Munc13 proteins in rat brain. Biochem. J. 1999, 337, 363–371. [Google Scholar] [CrossRef]
- Breustedt, J.; Gundlfinger, A.; Varoqueaux, F.; Reim, K.; Brose, N.; Schmitz, D. Munc13-2 differentially affects hippocampal synaptic transmission and plasticity. Cereb. Cortex 2010, 20, 1109–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiyama, S.; Schmidt, H.; Cooper, B.H.; Brose, N.; Eilers, J. Munc13-3 superprimes synaptic vesicles at granule cell-to-basket cell synapses in the mouse cerebellum. J. Neurosci. 2014, 34, 14687–14696. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, H.; Ariyoshi, T.; Kimpara, N.; Sugao, K.; Taiko, I.; Takikawa, K.; Asanuma, D.; Namiki, S.; Hirose, K. Synaptic weight set by Munc13-1 supramolecular assemblies. Nat. Neurosci. 2018, 21, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Böhme, M.A.; Beis, C.; Reddy-Alla, S.; Reynolds, E.; Mampell, M.M.; Grasskamp, A.T.; Lutzkendorf, J.; Bergeron, D.D.; Driller, J.H.; Babikir, H.; et al. Active zone scaffolds differentially accumulate Unc13 isoforms to tune Ca(2+) channel-vesicle coupling. Nat. Neurosci. 2016, 19, 1311–1320. [Google Scholar] [CrossRef]
- Hu, Z.; Tong, X.J.; Kaplan, J.M. UNC-13L, UNC-13S, and Tomosyn form a protein code for fast and slow neurotransmitter release in Caenorhabditis elegans. eLife 2013, 2, e00967. [Google Scholar] [CrossRef]
- Lipstein, N.; Schaks, S.; Dimova, K.; Kalkhof, S.; Ihling, C.; Kolbel, K.; Ashery, U.; Rhee, J.; Brose, N.; Sinz, A.; et al. Nonconserved Ca2+/Calmodulin Binding Sites in Munc13s Differentially Control Synaptic Short-Term Plasticity. Mol. Cell. Biol. 2012, 32, 4628–4641. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Cooper, B.; Kalla, S.; Varoqueaux, F.; Young, S.M., Jr. The Munc13 proteins differentially regulate readily releasable pool dynamics and calcium-dependent recovery at a central synapse. J. Neurosci. 2013, 33, 8336–8351. [Google Scholar] [CrossRef]
- Junge, H.J.; Rhee, J.S.; Jahn, O.; Varoqueaux, F.; Spiess, J.; Waxham, M.N.; Rosenmund, C.; Brose, N. Calmodulin and Munc13 form a Ca2+ sensor/effector complex that controls short-term synaptic plasticity. Cell 2004, 118, 389–401. [Google Scholar] [CrossRef] [Green Version]
- Lipstein, N.; Sakaba, T.; Cooper, B.H.; Lin, K.H.; Strenzke, N.; Ashery, U.; Rhee, J.S.; Taschenberger, H.; Neher, E.; Brose, N. Dynamic control of synaptic vesicle replenishment and short-term plasticity by Ca(2+)-calmodulin-Munc13-1 signaling. Neuron 2013, 79, 82–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Castaneda, F.; Maestre-Martinez, M.; Coudevylle, N.; Dimova, K.; Junge, H.; Lipstein, N.; Lee, D.; Becker, S.; Brose, N.; Jahn, O.; et al. Modular architecture of Munc13/calmodulin complexes: Dual regulation by Ca2+ and possible function in short-term synaptic plasticity. EMBO J. 2009, 29, 680–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinz, A. The advancement of chemical cross-linking and mass spectrometry for structural proteomics: From single proteins to protein interaction networks. Expert Rev. Proteom. 2014, 11, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Sinz, A. Divide and conquer: Cleavable cross-linkers to study protein conformation and protein-protein interactions. Anal. Bioanal. Chem. 2017, 409, 33–44. [Google Scholar] [CrossRef]
- Rappsilber, J. The beginning of a beautiful friendship: Cross-linking/mass spectrometry and modelling of proteins and multi-protein complexes. J. Struct. Biol. 2011, 173, 530–540. [Google Scholar] [CrossRef] [Green Version]
- Leitner, A.; Walzthoeni, T.; Kahraman, A.; Herzog, F.; Rinner, O.; Beck, M.; Aebersold, R. Probing native protein structures by chemical cross-linking, mass spectrometry, and bioinformatics. Mol. Cell. Proteom. MCP 2010, 9, 1634–1649. [Google Scholar] [CrossRef] [Green Version]
- Petrotchenko, E.V.; Borchers, C.H. Crosslinking combined with mass spectrometry for structural proteomics. Mass Spectrom. Rev. 2010, 29, 862–876. [Google Scholar] [CrossRef]
- Young, M.M.; Tang, N.; Hempel, J.C.; Oshiro, C.M.; Taylor, E.W.; Kuntz, I.D.; Gibson, B.W.; Dollinger, G. High throughput protein fold identification by using experimental constraints derived from intramolecular cross-links and mass spectrometry. Proc. Natl. Acad. Sci. USA 2000, 97, 5802–5806. [Google Scholar] [CrossRef] [Green Version]
- Lössl, P.; Kölbel, K.; Tänzler, D.; Nannemann, D.; Ihling, C.H.; Keller, M.V.; Schneider, M.; Zaucke, F.; Meiler, J.; Sinz, A. Analysis of Nidogen-1/Laminin gamma1 Interaction by Cross-Linking, Mass Spectrometry, and Computational Modeling Reveals Multiple Binding Modes. PLoS ONE 2014, 9, e112886. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Szarecka, A.; Yonkunas, M.; Speranskiy, K.; Kurnikova, M.; Cascio, M. Crosslinking constraints and computational models as complementary tools in modeling the extracellular domain of the glycine receptor. PLoS ONE 2014, 9, e102571. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, T.; Fischer, A.W.; Meiler, J.; Kalkhof, S. Protein structure prediction guided by crosslinking restraints—A systematic evaluation of the impact of the crosslinking spacer length. Methods 2015, 89, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.Q.; Schafer, M.; Dreiocker, F.; Ihling, C.H.; Sinz, A. Cleavable Cross-Linker for Protein Structure Analysis: Reliable Identification of Cross-Linking Products by Tandem MS. Anal. Chem. 2010, 82, 6958–6968. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, C.; Ihling, C.H.; Sinz, A. Extending the cross-linking/mass spectrometry strategy: Facile incorporation of photo-activatable amino acids into the model protein calmodulin in Escherichia coli cells. Methods 2015, 89, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Dimova, K.; Kalkhof, S.; Pottratz, I.; Ihling, C.; Rodriguez-Castaneda, F.; Liepold, T.; Griesinger, C.; Brose, N.; Sinz, A.; Jahn, O. Structural insights into the calmodulin-Munc13 interaction obtained by cross-linking and mass spectrometry. Biochemistry 2009, 48, 5908–5921. [Google Scholar] [CrossRef]
- Lössl, P.; Sinz, A. Combining Amine-Reactive Cross-Linkers and Photo-Reactive Amino Acids for 3D-Structure Analysis of Proteins and Protein Complexes. Methods Mol. Biol. 2016, 1394, 109–127. [Google Scholar] [CrossRef]
- Götze, M.; Pettelkau, J.; Schaks, S.; Bosse, K.; Ihling, C.H.; Krauth, F.; Fritzsche, R.; Kuhn, U.; Sinz, A. StavroX—A software for analyzing crosslinked products in protein interaction studies. J. Am. Soc. Mass Spectrom. 2012, 23, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Götze, M.; Pettelkau, J.; Fritzsche, R.; Ihling, C.H.; Schafer, M.; Sinz, A. Automated assignment of MS/MS cleavable cross-links in protein 3D-structure analysis. J. Am. Soc. Mass Spectrom. 2015, 26, 83–97. [Google Scholar] [CrossRef]
- Kalkhof, S.; Sinz, A. Chances and pitfalls of chemical cross-linking with amine-reactive N-hydroxysuccinimide esters. Anal. Bioanal. Chem. 2008, 392, 305–312. [Google Scholar] [CrossRef]
- Raveh, B.; London, N.; Zimmerman, L.; Schueler-Furman, O. Rosetta FlexPepDock ab-initio: Simultaneous folding, docking and refinement of peptides onto their receptors. PLoS ONE 2011, 6, e18934. [Google Scholar] [CrossRef] [Green Version]
- Valentine, K.G.; Ng, H.L.; Schneeweis, L.; Kranz, J.K.; Frederick, K.K.; Alber, T.; Wand, A.J. Crystal Structure of Calmodulin-Neuronal Nitric Oxide Synthase Complex; RCSB PDB: Piscataway, NJ, USA. [CrossRef]
- Iacobucci, C.; Gotze, M.; Piotrowski, C.; Arlt, C.; Rehkamp, A.; Ihling, C.; Hage, C.; Sinz, A. Carboxyl-Photo-Reactive MS-Cleavable Cross-Linkers: Unveiling a Hidden Aspect of Diazirine-Based Reagents. Anal. Chem. 2018, 90, 2805–2809. [Google Scholar] [CrossRef]
- Zikich, D.; Mezer, A.; Varoqueaux, F.; Sheinin, A.; Junge, H.J.; Nachliel, E.; Melamed, R.; Brose, N.; Gutman, M.; Ashery, U. Vesicle priming and recruitment by ubMunc13-2 are differentially regulated by calcium and calmodulin. J. Neurosci. 2008, 28, 1949–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneggenburger, R.; Neher, E. Presynaptic calcium and control of vesicle fusion. Curr. Opin. Neurobiol. 2005, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Merkley, E.D.; Rysavy, S.; Kahraman, A.; Hafen, R.P.; Daggett, V.; Adkins, J.N. Distance restraints from crosslinking mass spectrometry: Mining a molecular dynamics simulation database to evaluate lysine-lysine distances. Protein Sci. A Publ. Protein Soc. 2014, 23, 747–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimova, K.; Kawabe, H.; Betz, A.; Brose, N.; Jahn, O. Characterization of the Munc13-calmodulin interaction by photoaffinity labeling. Biochim. Et Biophys. Acta 2006, 1763, 1256–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufmann, K.W.; Lemmon, G.H.; Deluca, S.L.; Sheehan, J.H.; Meiler, J. Practically useful: What the Rosetta protein modeling suite can do for you. Biochemistry 2010, 49, 2987–2998. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.; Tanzler, D.; Ihling, C.H.; Sinz, A. Monitoring Solution Structures of Peroxisome Proliferator-Activated Receptor beta/delta upon Ligand Binding. PLoS ONE 2016, 11, e0151412. [Google Scholar] [CrossRef]
- Göth, M. Investigation of Protein-Ligand Complexes by Native Mass Spectrometry and Ion Mobility-Mass Spectrometry. Diss. Freie Univ. Berl. 2017. [Google Scholar] [CrossRef]
- Osawa, M.; Tokumitsu, H.; Swindells, M.B.; Kurihara, H.; Orita, M.; Shibanuma, T.; Furuya, T.; Ikura, M. A novel target recognition revealed by calmodulin in complex with Ca2+-calmodulin-dependent kinase kinase. Nat. Struct. Biol. 1999, 6, 819–824. [Google Scholar] [CrossRef]
- Glaser, P.; Elmaoglou-Lazaridou, A.; Krin, E.; Ladant, D.; Barzu, O.; Danchin, A. Identification of residues essential for catalysis and binding of calmodulin in Bordetella pertussis adenylate cyclase by site-directed mutagenesis. EMBO J. 1989, 8, 967–972. [Google Scholar] [CrossRef]
- Tokumitsu, H.; Wayman, G.A.; Muramatsu, M.; Soderling, T.R. Calcium/calmodulin-dependent protein kinase kinase: Identification of regulatory domains. Biochemistry 1997, 36, 12823–12827. [Google Scholar] [CrossRef]
- De Diego, I.; Kuper, J.; Bakalova, N.; Kursula, P.; Wilmanns, M. Molecular basis of the death-associated protein kinase-calcium/calmodulin regulator complex. Sci. Signal. 2010, 3, ra6. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Stengel, F.; Aebersold, R.; Gold, M.G. Molecular basis of AKAP79 regulation by calmodulin. Nat. Commun. 2017, 8, 1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbst, S.; Maucher, D.; Schneider, M.; Ihling, C.H.; Jahn, O.; Sinz, A. Munc13-like skMLCK variants cannot mimic the unique calmodulin binding mode of Munc13 as evidenced by chemical cross-linking and mass spectrometry. PLoS ONE 2013, 8, e75119. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, K.T.; Erickson-Vlitanen, S.; DeGrado, W.F. Photolabeling of calmodulin with basic, amphiphilic alpha-helical peptides containing benzoylphenylalanine. J. Biol. Chem. 1989, 264, 14571–14578. [Google Scholar]
- Varoqueaux, F.; Sigler, A.; Rhee, J.S.; Brose, N.; Enk, C.; Reim, K.; Rosenmund, C. Total arrest of spontaneous and evoked synaptic transmission but normal synaptogenesis in the absence of Munc13-mediated vesicle priming. Proc. Natl. Acad. Sci. USA 2002, 99, 9037–9042. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cross-Linker | Cross-Linked Amino Acid CaM | Cross-Linked Amino Acid Segment-A | Ca2+ Concentration | |
|---|---|---|---|---|
| 750 nM | 1 mM | |||
| Photo-Met | x36 | D615 | N | Y |
| x36 | E712 | Y | Y | |
| x71/x72 | E707 | Y | Y | |
| x71/x72 | E740 | N | Y | |
| x76 | H578 | Y | Y | |
| x76 | E707 | Y | Y | |
| x76 | E729 | N | Y | |
| x144/x145 | H578/H579 | Y | Y | |
| x144/x145 | E707 | N | Y | |
| x144/x145 | Y715 | Y | Y | |
| x144/x145 | C719 | Y | Y | |
| x144/x145 | E729 | Y | Y | |
| DSBU | K75 | K735 | N | Y |
| s-GMBS | K75 | C453 | N | Y |
| K13 | C428 | N | Y | |
| K30 | C428 | N | Y | |
| Cross-Linked Amino Acid CaM | Cross-Linked Amino Acid bMunc13-2 | Segment-A (aa 367–780) | Segment-B (aa 367–903) | Segment-C (aa 703–742) |
|---|---|---|---|---|
| x36 | E707 | ✓ | ||
| x36 | D615 | ✓ | ||
| x36 | E712 | ✓ | ✓ | |
| x71/x72 | E707 | ✓ | ✓ | ✓ |
| x71/x72 | E729 | ✓ | ||
| x71/x72 | E740 | ✓ | ✓ | |
| x76 | H578 | ✓ | ||
| x76 | E707 | ✓ | ✓ | ✓ |
| x76 | E729 | ✓ | ✓ | |
| x124 | E729 | ✓ | ||
| x144/x145 | H578/H579 | ✓ | ||
| x144/x145 | E707 | ✓ | ✓ | ✓ |
| x144/x145 | Y715 | ✓ | ✓ | ✓ |
| x144/x145 | C719 | ✓ | ✓ | ✓ |
| x144/x145 | E729 | ✓ | ✓ |
| Amino Acid Sequences of Segment-C | Variants of Segment-C |
|---|---|
| 703KLSLEQVCAETIYLNKCINNFKNVLREKRLRQKKLLQELV742 | Segment-C |
| 703KLSLEQVCAETIYLNKCINNfKNVLREKRLRQKKLLQELV742 | Segment-C-Bpa |
| 719CINNFKNVLREKRLRQKKLLQELV742 | Segment-CS |
| 719CINNfKNVLREKRLRQKKLLQELV742 | Segment-CS-Bpa |
| 719CINNFKNVLREKRLRQKKLLQELVQTASHLSVED742 | Segment-CL |
| 703KLSLDQVCAETIYLNKCINNFKNVLREKRLRQKKLLQELV742 | Segment-C-V709D |
| 703KLSLEQVCAETDYLNKCINNFKNVLREKRLRQKKLLQELV742 | Segment-C-I714D |
| 703KLSLEQVCAETIYLNKCINNDKNVLREKRLRQKKLLQELV742 | Segment-C-F723D |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piotrowski, C.; Moretti, R.; Ihling, C.H.; Haedicke, A.; Liepold, T.; Lipstein, N.; Meiler, J.; Jahn, O.; Sinz, A. Delineating the Molecular Basis of the Calmodulin–bMunc13-2 Interaction by Cross-Linking/Mass Spectrometry—Evidence for a Novel CaM Binding Motif in bMunc13-2. Cells 2020, 9, 136. https://doi.org/10.3390/cells9010136
Piotrowski C, Moretti R, Ihling CH, Haedicke A, Liepold T, Lipstein N, Meiler J, Jahn O, Sinz A. Delineating the Molecular Basis of the Calmodulin–bMunc13-2 Interaction by Cross-Linking/Mass Spectrometry—Evidence for a Novel CaM Binding Motif in bMunc13-2. Cells. 2020; 9(1):136. https://doi.org/10.3390/cells9010136
Chicago/Turabian StylePiotrowski, Christine, Rocco Moretti, Christian H. Ihling, André Haedicke, Thomas Liepold, Noa Lipstein, Jens Meiler, Olaf Jahn, and Andrea Sinz. 2020. "Delineating the Molecular Basis of the Calmodulin–bMunc13-2 Interaction by Cross-Linking/Mass Spectrometry—Evidence for a Novel CaM Binding Motif in bMunc13-2" Cells 9, no. 1: 136. https://doi.org/10.3390/cells9010136
APA StylePiotrowski, C., Moretti, R., Ihling, C. H., Haedicke, A., Liepold, T., Lipstein, N., Meiler, J., Jahn, O., & Sinz, A. (2020). Delineating the Molecular Basis of the Calmodulin–bMunc13-2 Interaction by Cross-Linking/Mass Spectrometry—Evidence for a Novel CaM Binding Motif in bMunc13-2. Cells, 9(1), 136. https://doi.org/10.3390/cells9010136