Hydrogen Sulfide Impairs Meiosis Resumption in Xenopus laevis Oocytes

 , , , and
, , , and 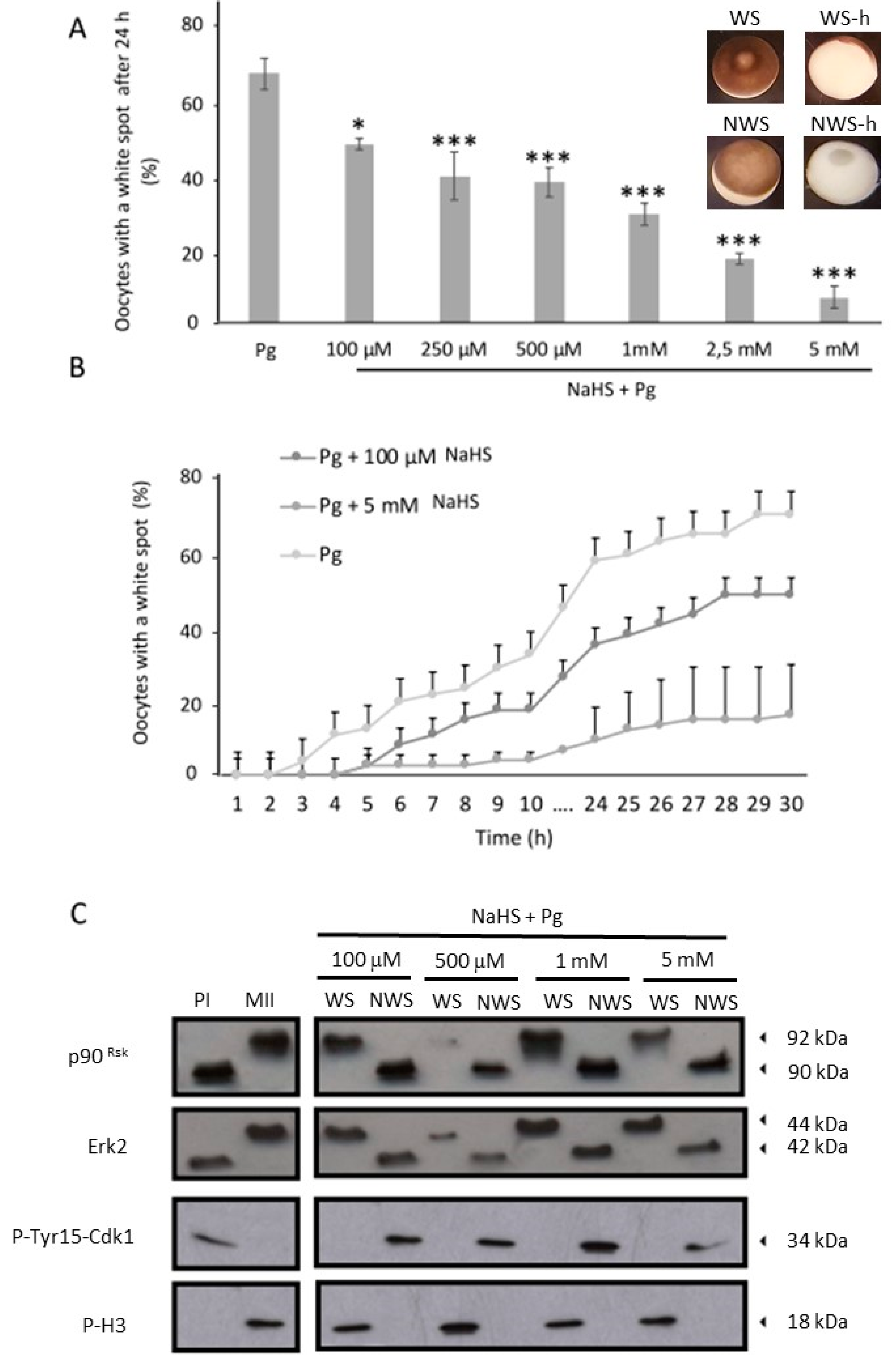{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Frog and Oocyte Handling
2.3. Oocyte treatments, mRNA Micro-Injections, and Meiotic Resumption Analysis
2.4. Mature Oocyte Cytoplasm Micro-Injections
2.5. Heat Fixation and GVBD Analysis
2.6. Electrophoresis and Western blot
2.7. Immunoprecipitations
2.8. Histone H1 Kinase Assay
2.9. Apoptosis Analysis
2.10. H2S Production
2.11. Cdc25C Phosphatase Activity Assay
2.12. Statistical Analysis
3. Results
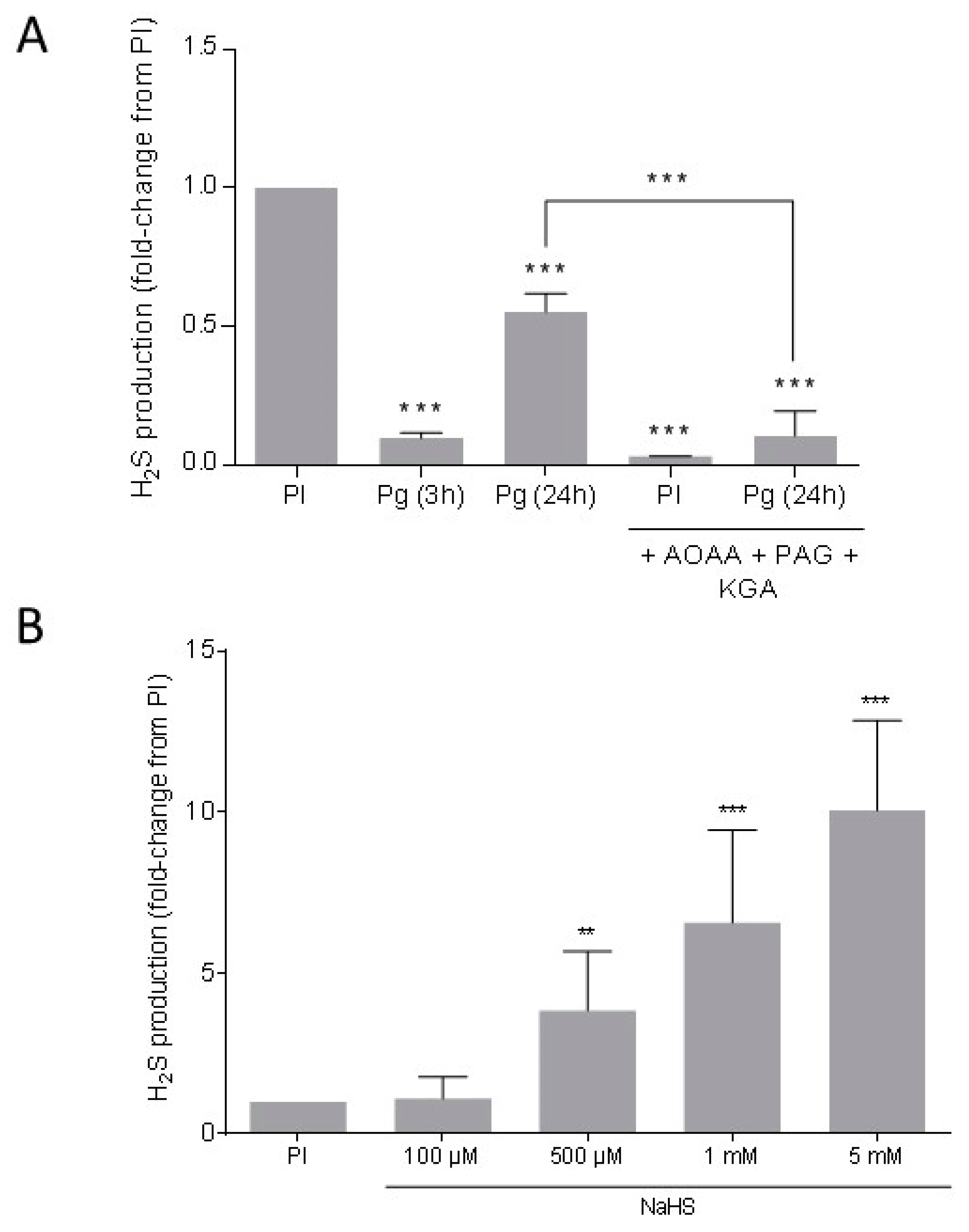
3.1. NaHS, an H2S Donor, Prevents the Xenopus Oocytes Meiotic Resumption in a Dose-Dependent Manner
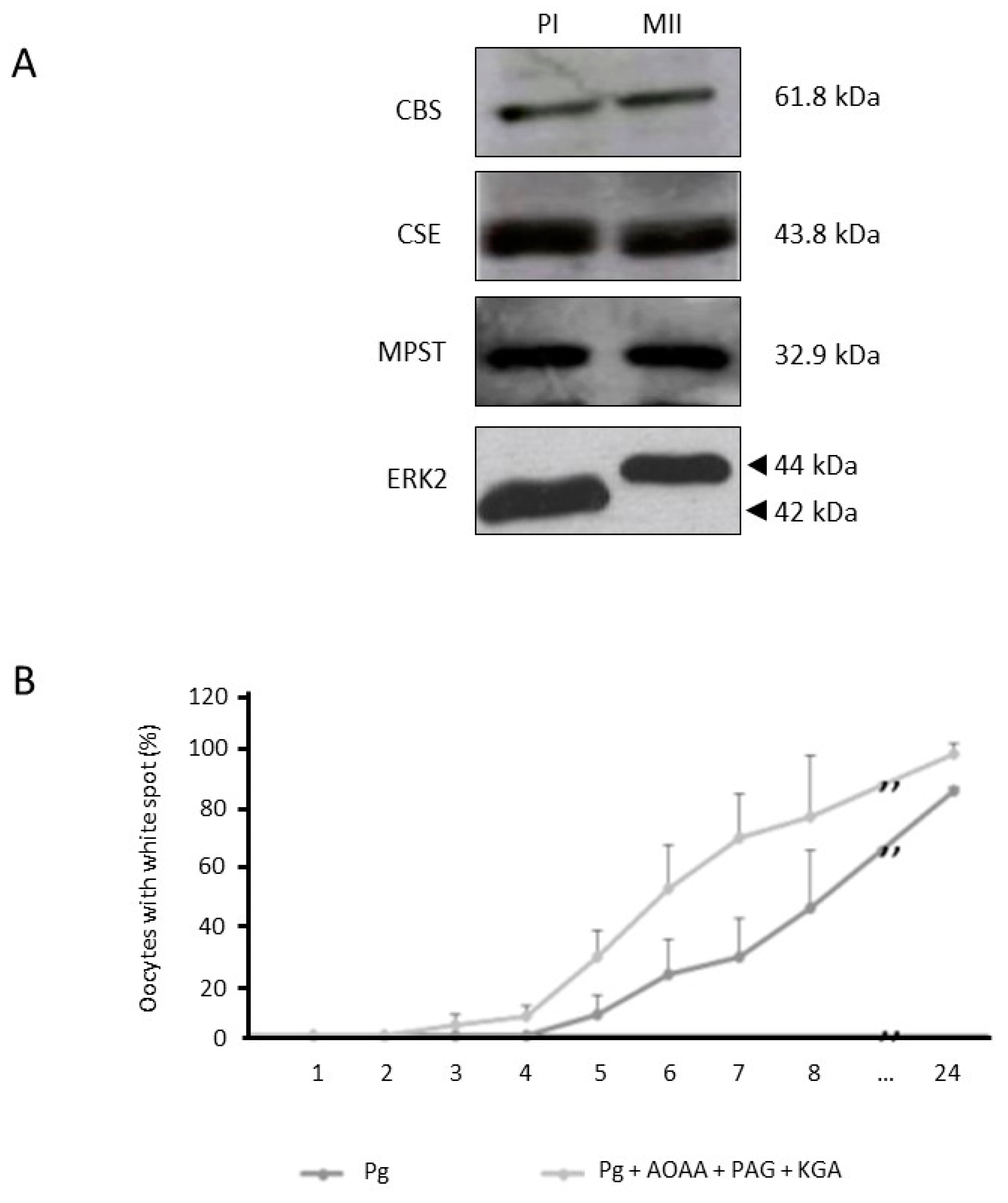
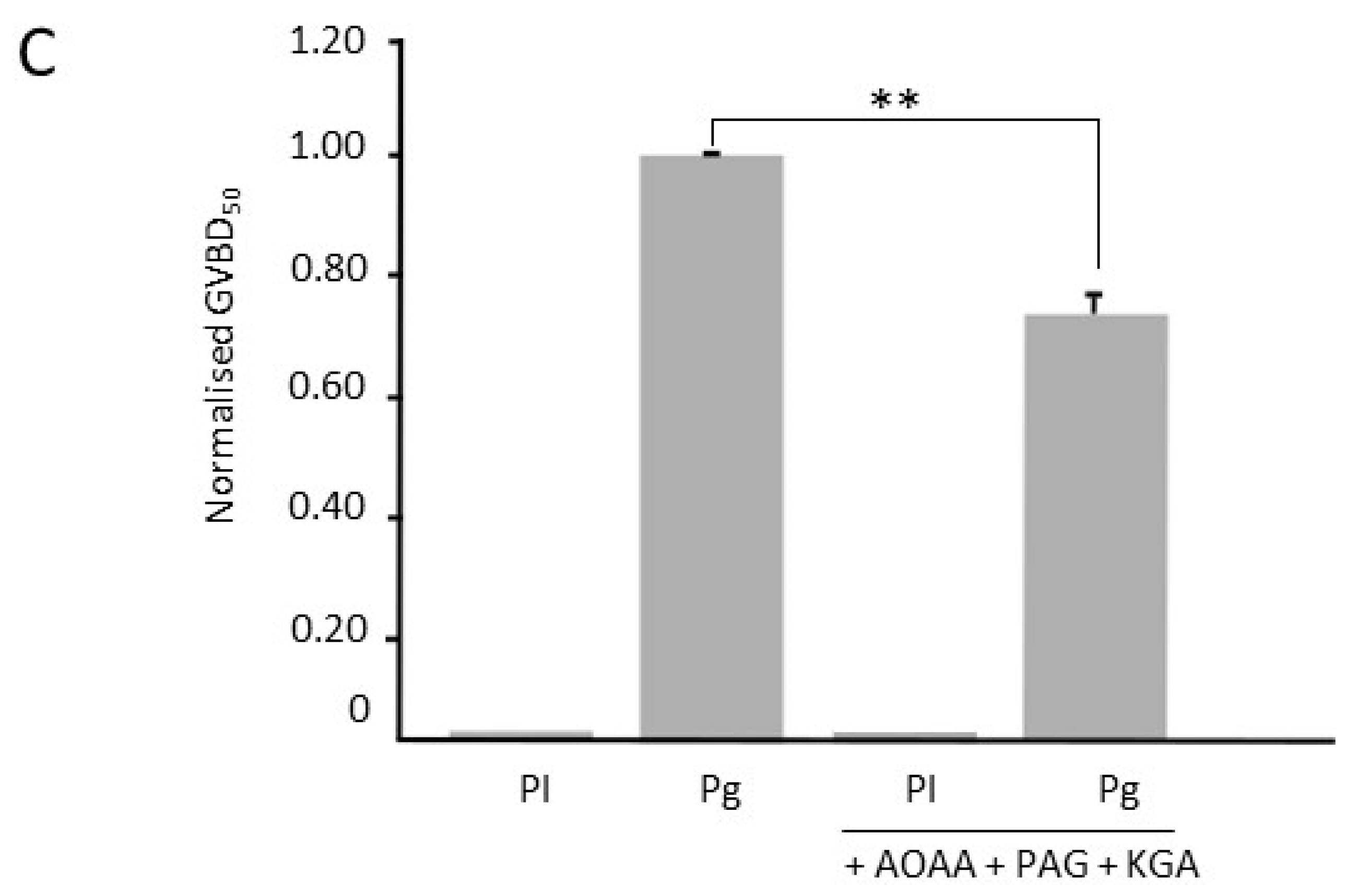
3.2. Inhibition of H2S Metabolism Key Enzymes Accelerates Oocyte Meiotic Resumption
3.3. Endogenous H2S Is Produced in Xenopus Oocytes
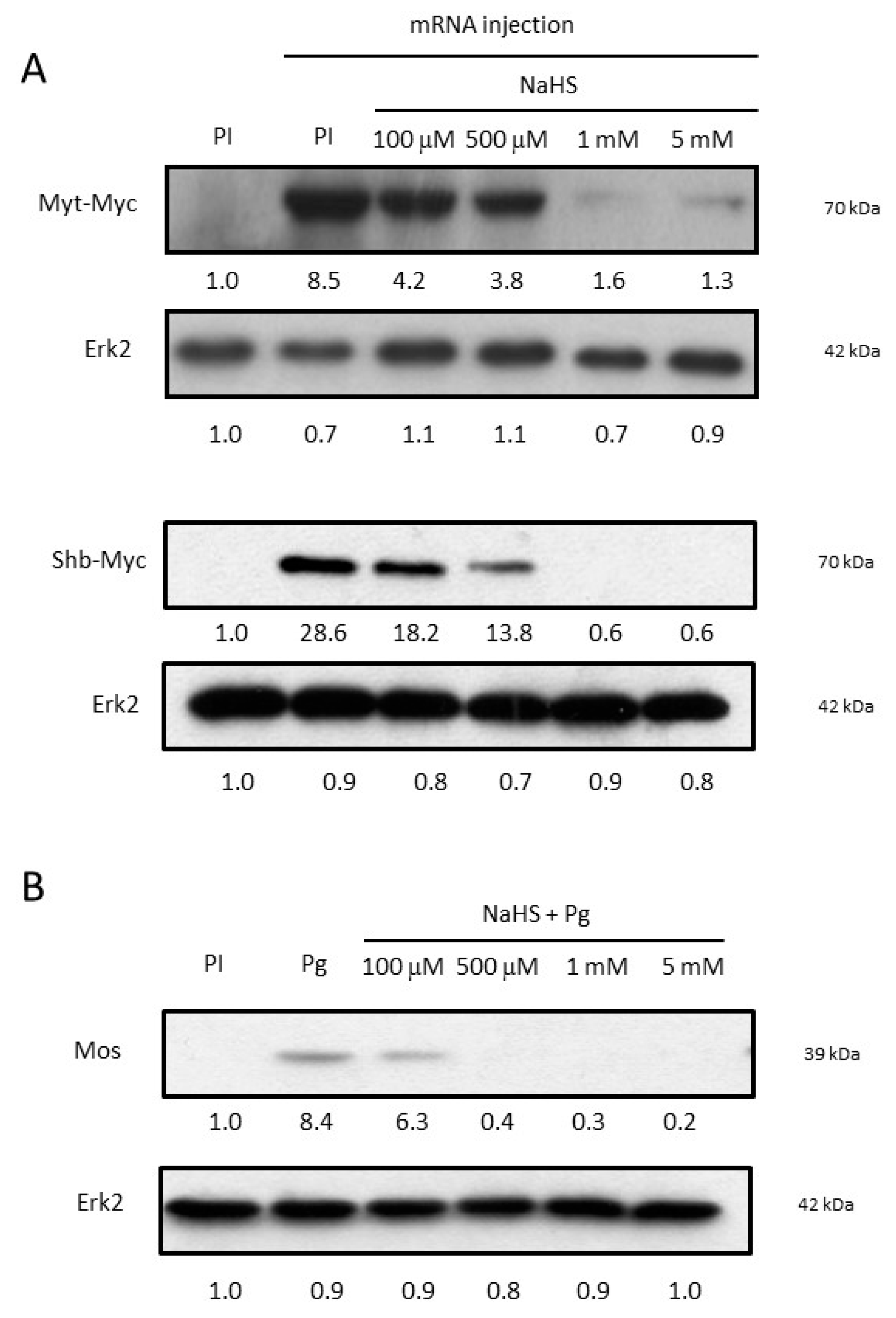
3.4. NaHS Impairs Oocyte Protein Synthesis
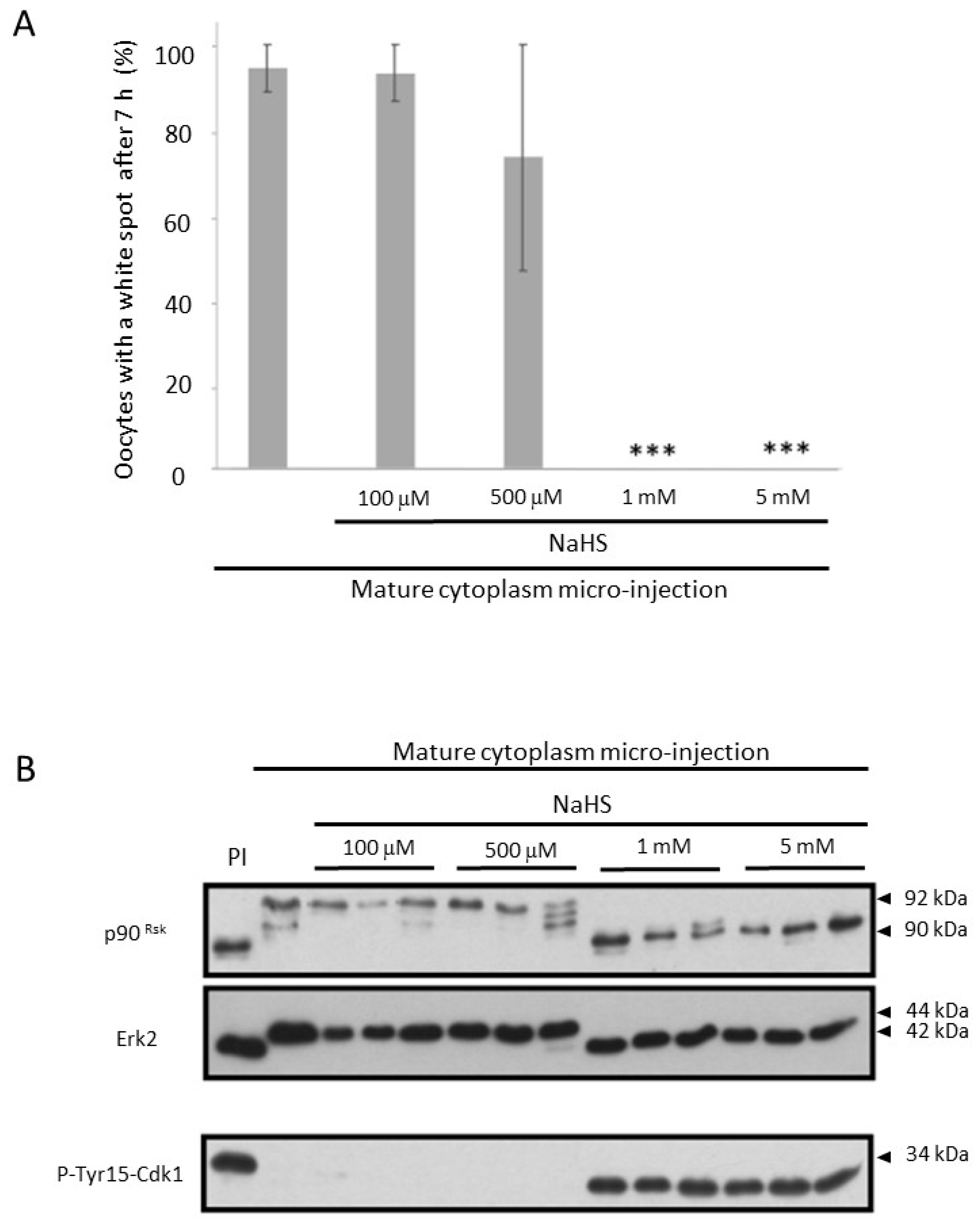
3.5. NaHS Impairs the Self-Amplification Loop of MPF
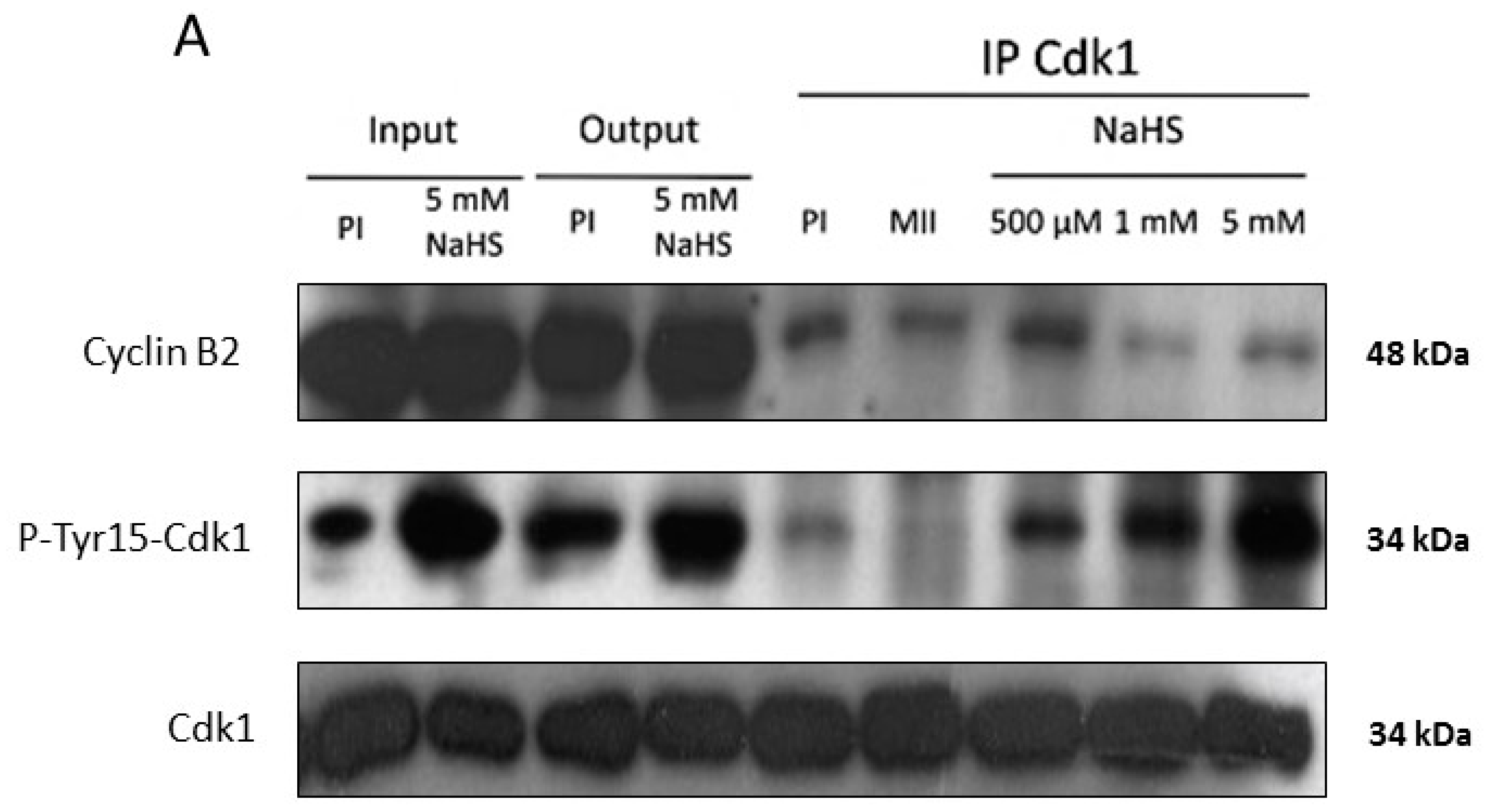
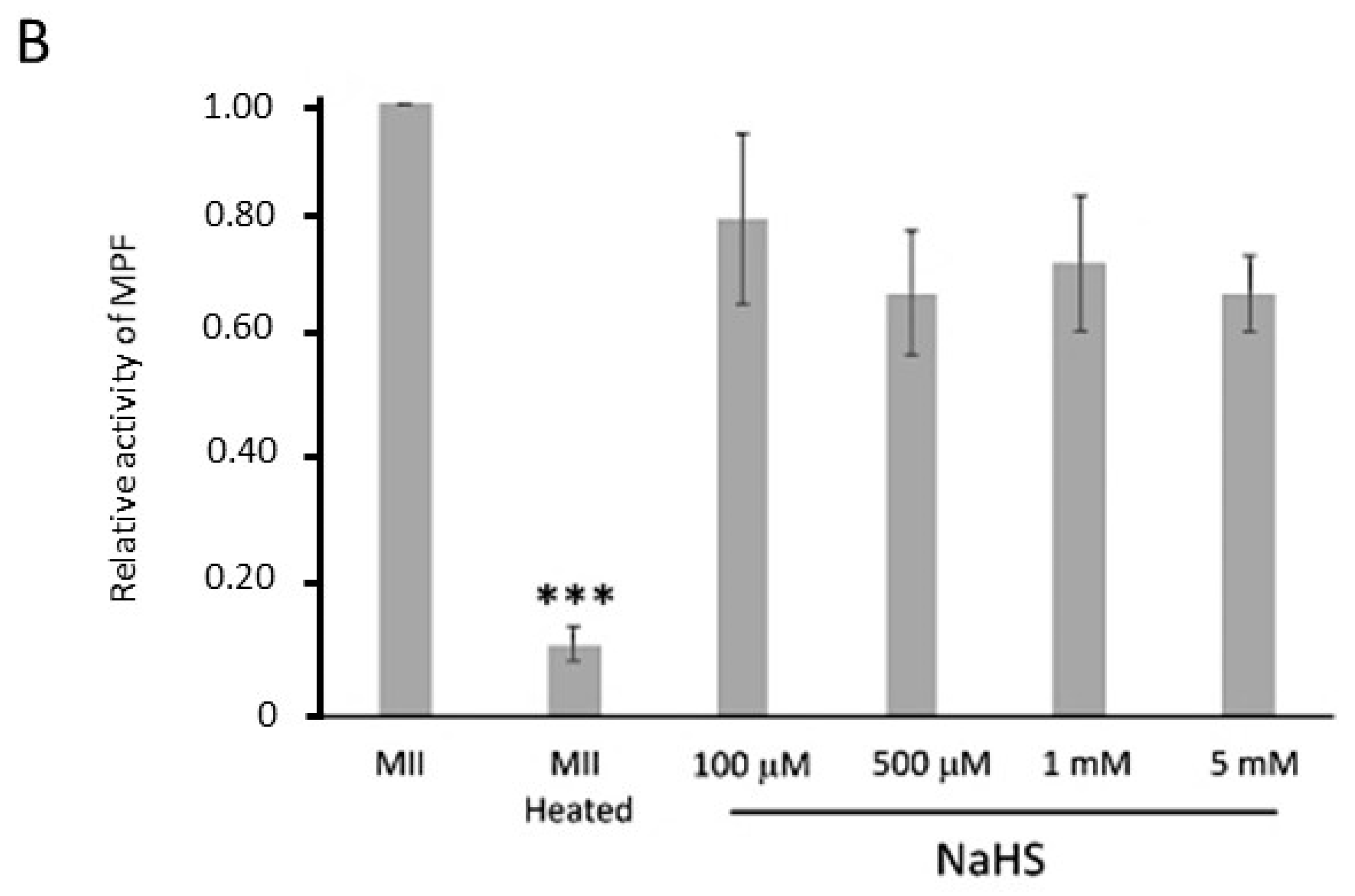
3.6. NaHS Does Not Dissociate the Pre-MPF Complexes Nor Impact MPF Activity
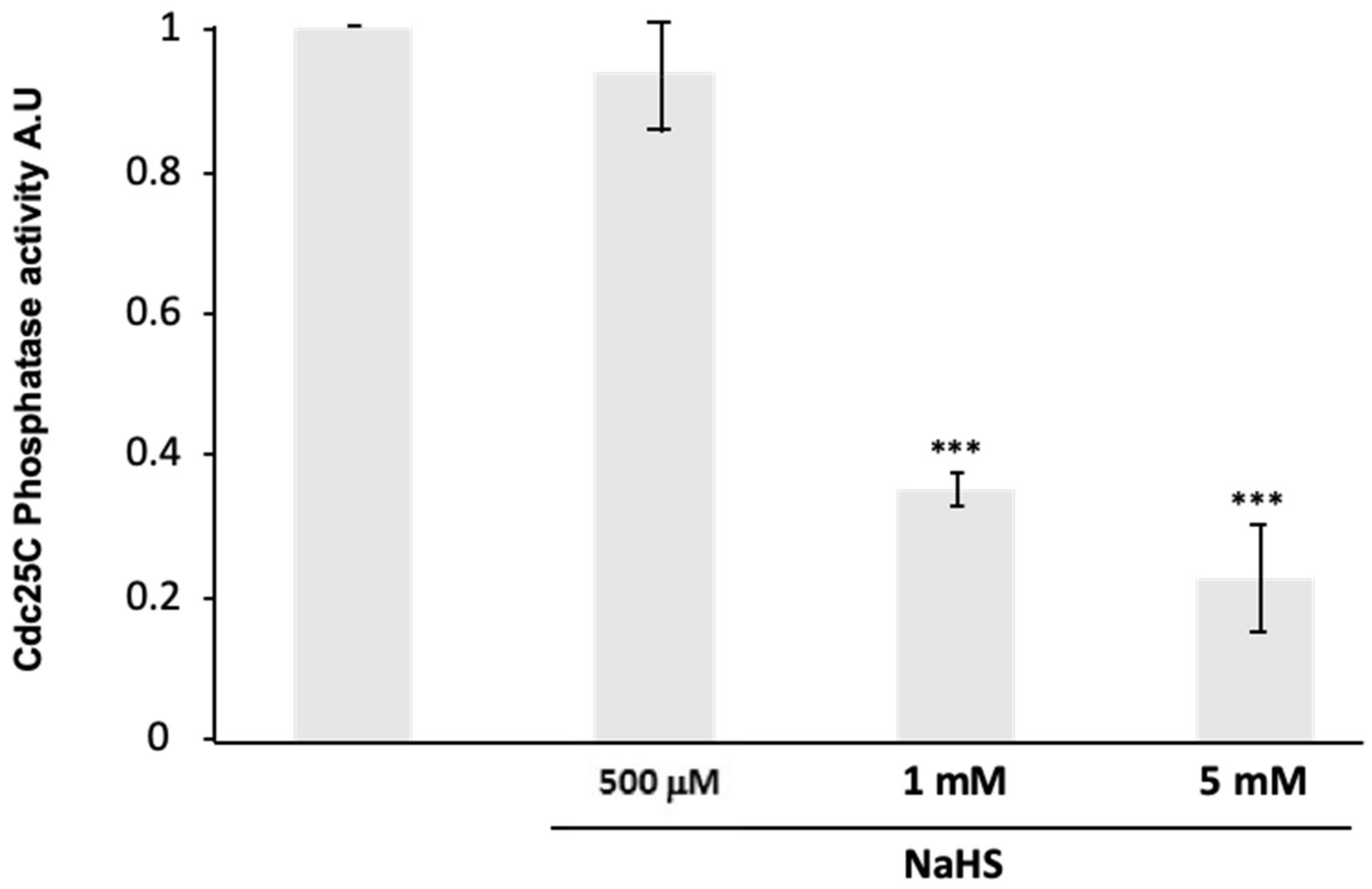
3.7. NaHS Impairs Human Cdc25C Activity In Vitro
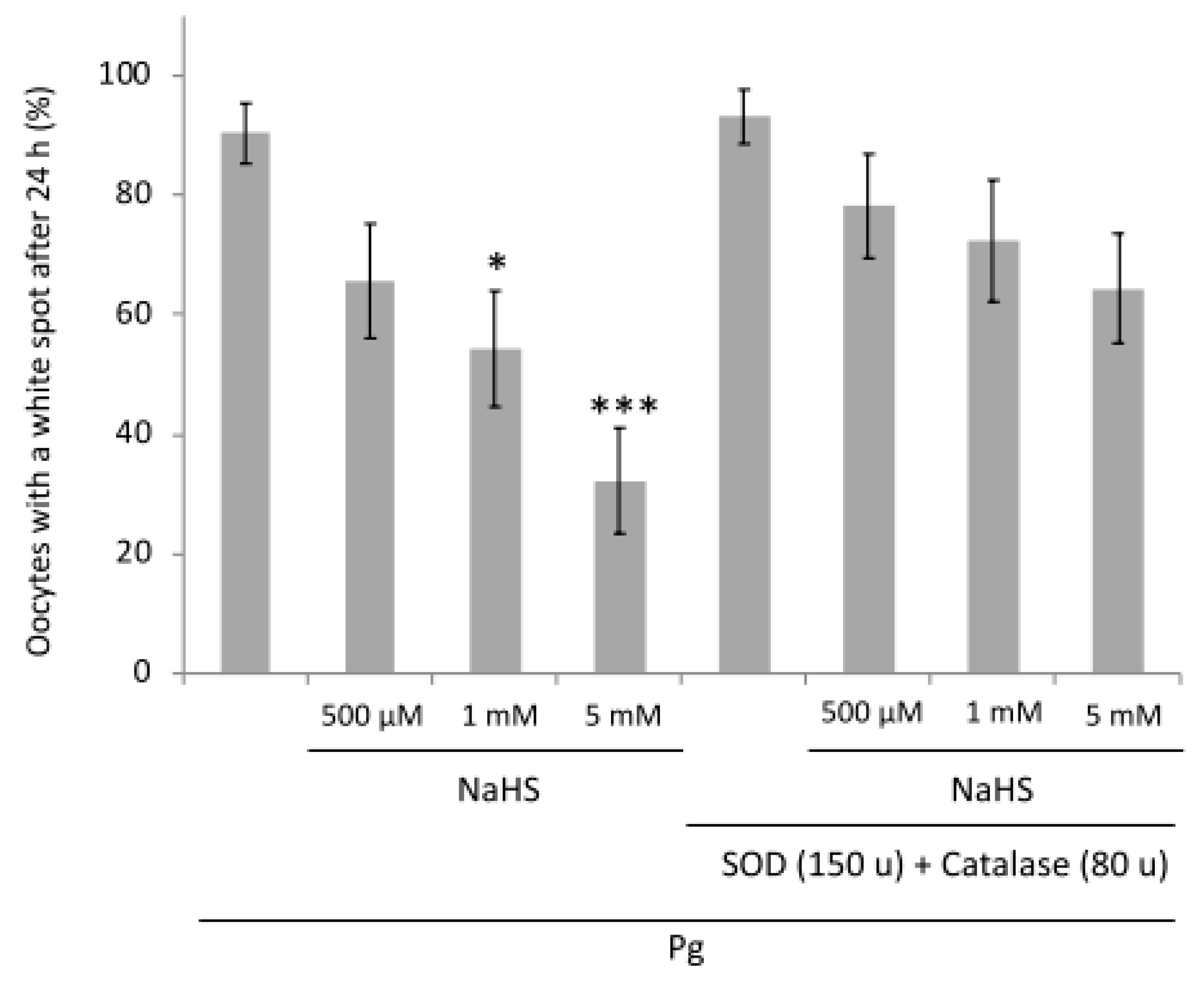
3.8. NaHS Effects Are Partially ROS-Dependent
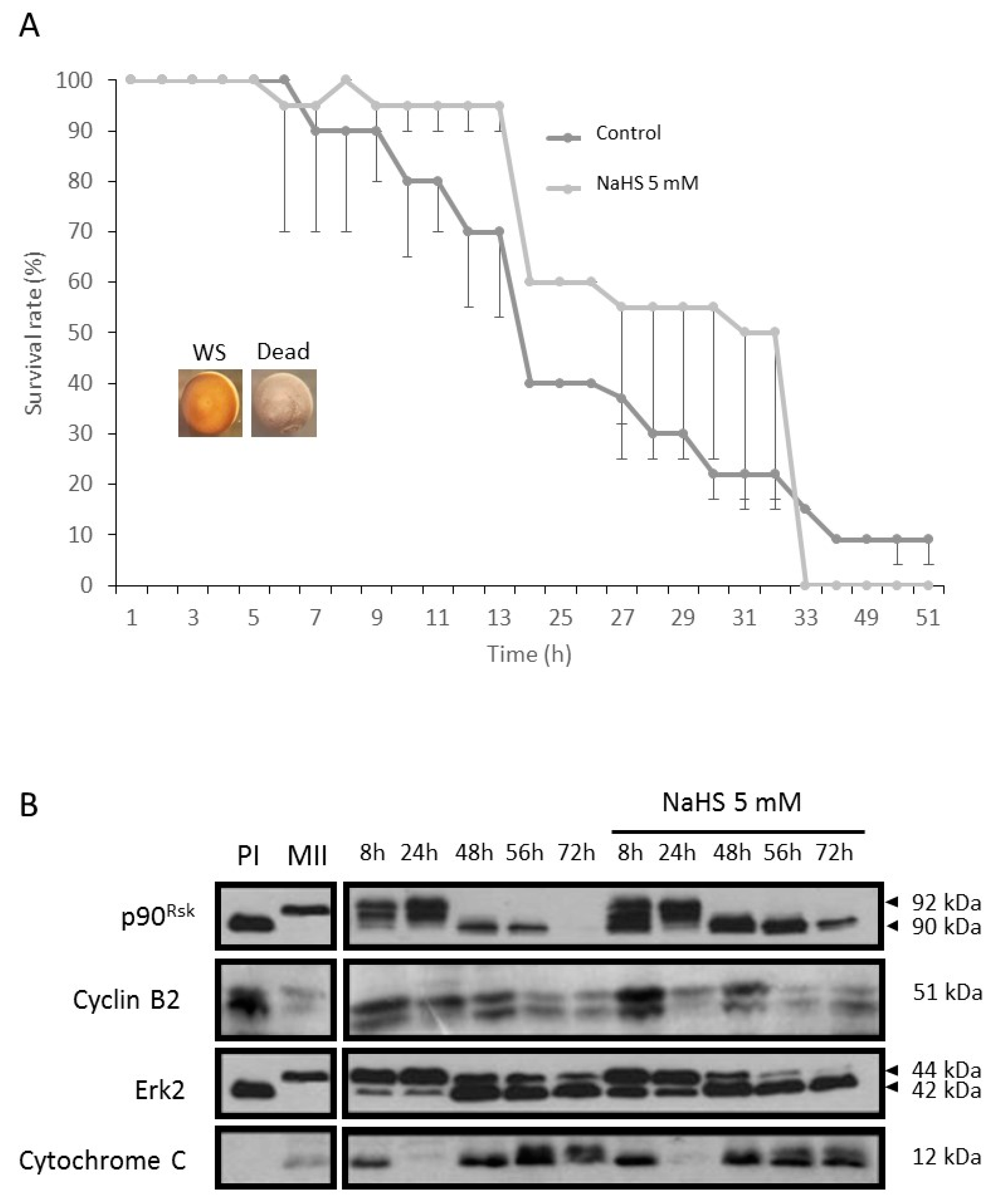
3.9. NaHS Does Not Promote Nor Protect from Apoptosis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Qu, K.; Lee, S.W.; Bian, J.S.; Low, C.-M.; Wong, P.T.-H. Hydrogen sulfide: Neurochemistry and neurobiology. Neurochem. Int. 2008, 52, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-B.; Du, J.-B.; Cui, H. Signal pathways involved in the biological effects of sulfur dioxide. Eur. J. Pharmacol. 2015, 764, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Two’s company, three’s a crowd: Can H2S be the third endogenous gaseous transmitter? FASEB J. 2002, 16, 1792–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate Sulfurtransferase Produces Hydrogen Sulfide and Bound Sulfane Sulfur in the Brain. Antioxid. Redox Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Chan, S.-J.; Ng, Y.-K.; Wong, P.T.-H. Brain 3-Mercaptopyruvate Sulfurtransferase (3MST): Cellular Localization and Downregulation after Acute Stroke. PLoS ONE 2013, 8, e67322. [Google Scholar] [CrossRef] [Green Version]
- Liang, R.; Yu, W.; Du, J.; Yang, L.; Shang, M.; Guo, J. Localization of cystathionine beta synthase in mice ovaries and its expression profile during follicular development. Chin. Med. J. (Engl.) 2006, 119, 1877–1883. [Google Scholar] [CrossRef]
- Sugiura, Y.; Kashiba, M.; Maruyama, K.; Hoshikawa, K.; Sasaki, R.; Saito, K.; Kimura, H.; Goda, N.; Suematsu, M. Cadmium exposure alters metabolomics of sulfur-containing amino acids in rat testes. Antioxid. Redox Signal. 2005, 7, 781–787. [Google Scholar] [CrossRef]
- Patel, P.; Vatish, M.; Heptinstall, J.; Wang, R.; Carson, R.J. The endogenous production of hydrogen sulphide in intrauterine tissues. Reprod. Biol. Endocrinol. 2009, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Srilatha, B.; Hu, L.; Adaikan, G.P.; Moore, P.K. Initial characterization of hydrogen sulfide effects in female sexual function. J. Sex. Med. 2009, 6, 1875–1884. [Google Scholar] [CrossRef]
- Guzmán, M.A.; Navarro, M.A.; Carnicer, R.; Sarría, A.J.; Acín, S.; Arnal, C.; Muniesa, P.; Surra, J.C.; Arbonés-Mainar, J.M.; Maeda, N.; et al. Cystathionine beta-synthase is essential for female reproductive function. Hum. Mol. Genet. 2006, 15, 3168–3176. [Google Scholar] [CrossRef] [Green Version]
- Oi, Y.; Imafuku, M.; Shishido, C.; Kominato, Y.; Nishimura, S.; Iwai, K. Garlic supplementation increases testicular testosterone and decreases plasma corticosterone in rats fed a high protein diet. J. Nutr. 2001, 131, 2150–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srilatha, B.; Adaikan, P.G.; Moore, P.K. Possible role for the novel gasotransmitter hydrogen sulphide in erectile dysfunction—A pilot study. Eur. J. Pharmacol. 2006, 535, 280–282. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, M.; Dehpour, A.R.; Moore, K.P.; Mani, A.R. Role of endogenous hydrogen sulfide in neurogenic relaxation of rat corpus cavernosum. Biochem. Pharmacol. 2012, 83, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Srilatha, B.; Adaikan, P.G.; Li, L.; Moore, P.K. Hydrogen sulphide: A novel endogenous gasotransmitter facilitates erectile function. J. Sex. Med. 2007, 4, 1304–1311. [Google Scholar] [CrossRef]
- D’Emmanuele di Villa Bianca, R.; Sorrentino, R.; Maffia, P.; Mirone, V.; Imbimbo, C.; Fusco, F.; De Palma, R.; Ignarro, L.J.; Cirino, G. Hydrogen sulfide as a mediator of human corpus cavernosum smooth-muscle relaxation. Proc. Natl. Acad. Sci. USA 2009, 106, 4513–4518. [Google Scholar] [CrossRef] [Green Version]
- Hemminki, K.; Niemi, M.L. Community study of spontaneous abortions : Relation to occupation and air pollution by sulfur dioxid, hydrogen sulfide, and carbon disulfide. Int. Arch. Occup. Environ. Health. 1982, 51, 55–63. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Gu, H.; Ni, X. Hydrogen sulfide in the endocrine and reproductive systems. Expert Rev. Clin. Pharmacol. 2011, 4, 75–82. [Google Scholar] [CrossRef]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Liang, R.; Yu, W.-D.; Du, J.-B.; Yang, L.-J.; Yang, J.-J.; Xu, J.; Shang, M.; Guo, J.-Z. Cystathionine beta synthase participates in murine oocyte maturation mediated by homocysteine. Reprod. Toxicol. 2007, 24, 89–96. [Google Scholar] [CrossRef]
- Krejcova, T.; Smelcova, M.; Petr, J.; Bodart, J.-F.; Sedmikova, M.; Nevoral, J.; Dvorakova, M.; Vyskocilova, A.; Weingartova, I.; Kucerova-Chrpova, V.; et al. Hydrogen sulfide donor protects porcine oocytes against aging and improves the developmental potential of aged porcine oocytes. PLoS ONE 2015, 10, e0116964. [Google Scholar] [CrossRef] [Green Version]
- Nevoral, J.; Žalmanová, T.; Zámostná, K.; Kott, T.; Kučerová-Chrpová, V.; Bodart, J.-F.; Gelaude, A.; Procházka, R.; Orsák, M.; Šulc, M.; et al. Endogenously produced hydrogen sulfide is involved in porcine oocyte maturation in vitro. Nitric Oxide 2015, 51, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Nevoral, J.; Petr, J.; Gelaude, A.; Bodart, J.F.; Kucerova-Chrpova, V.; Sedmikova, M.; Krejcova, T.; Kolbabova, T.; Dvorakova, M.; Vyskocilova, A.; et al. Dual effects of hydrogen sulfide donor on meiosis and cumulus expansion of porcine cumulus-oocyte complexes. PLoS ONE 2014, 1, 9. [Google Scholar] [CrossRef] [PubMed]
- Masui, Y.; Markert, C.L. Cytoplasmic control of nuclear behavior during meiotic maturation of frog oocytes. J. Exp. Zool. 1971, 177, 129–145. [Google Scholar] [CrossRef]
- Rime, H.; Huchon, D.; De Smedt, V.; Thibier, C.; Galaktionov, K.; Jessus, C.; Ozon, R. Microinjection of Cdc25 protein phosphatase into Xenopus prophase oocyte activates MPF and arrests meiosis at metaphase I. Biol. Cell. 1994, 82, 11–22. [Google Scholar] [CrossRef]
- Ferrell, J.E.; Wu, M.; Gerhart, J.C.; Martin, G.S. Cell cycle tyrosine phosphorylation of p34cdc2 and a microtubule-associated protein kinase homolog in Xenopus oocytes and eggs. Mol. Cell. Biol. 1991, 11, 1965–1971. [Google Scholar] [CrossRef] [PubMed]
- Baert, F.; Bodart, J.-F.; Bocquet-Muchembled, B.; Lescuyer-Rousseau, A.; Vilain, J.-P. Xp42(Mpk1) activation is not required for germinal vesicle breakdown but for Raf complete phosphorylation in insulin-stimulated Xenopus oocytes. J. Biol. Chem. 2003, 278, 49714–49720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupré, A.; Jessus, C.; Ozon, R.; Haccard, O. Mos is not required for the initiation of meiotic maturation in Xenopus oocytes. EMBO J. 2002, 21, 4026–4036. [Google Scholar] [CrossRef] [Green Version]
- Furuno, N.; Kawasaki, A.; Sagata, N. Expression of cell-cycle regulators during Xenopus oogenesis. Gene Expr. Patterns 2003, 3, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Ferrell, J.E., Jr.; Machleder, E.M. The biochemical basis of an all-or-none cell fate switch in Xenopus oocytes. Science 1998, 280, 895–898. [Google Scholar] [CrossRef] [Green Version]
- Bagowski, C.P.; Besser, J.; Frey, C.R.; Ferrell, J.E., Jr. The JNK cascade as a biochemical switch in mammalian cells: Ultrasensitive and all-or-none responses. Curr. Biol. 2003, 13, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, R.R.; Ferrell, J.E., Jr. The protein kinase p90 rsk as an essential mediator of cytostatic factor activity. Science 1999, 286, 1362–1365. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, R.R.; Ferrell, J.E., Jr. Cloning and characterization of Xenopus Rsk2, the predominant p90 Rsk isozyme in oocytes and eggs. J. Biol. Chem. 2000, 275, 32983–32990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, C. Hydrogen sulphide and its therapeutic potential. Nat. Rev. Drug Discov. 2007, 6, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Gaffré, M.; Martoriati, A.; Belhachemi, N.; Chambon, J.P.; Houliston, E.; Jessus, C.; Karaiskou, A. A critical balance between Cyclin B synthesis and Myt1 activity controls meiosis entry in Xenopus oocytes. Development 2011, 138, 3735–3744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, M.; Vanderstraete, M.; Cailliau, K.; Hahnel, S.; Grevelding, C.G.; Dissous, C. SmShb, the SH2-Containing Adaptor Protein B of Schistosoma mansoni Regulates Venus Kinase Receptor Signaling Pathways. PLoS ONE 2016, 11, e0163283. [Google Scholar] [CrossRef] [PubMed]
- Tokmakov, A.A.; Iguchi, S.; Iwasaki, T.; Fukami, Y. Unfertilized frog eggs die by apoptosis following meiotic exit. BMC Cell Biol. 2011, 12, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigel, E. Use of Xenopus oocytes for the functional expression of plasma membrane proteins. J. Membr. Biol. 1990, 117, 201–221. [Google Scholar] [CrossRef]
- Cailliau, K.; Browaeys-Poly, E. A microinjectable biological system, the Xenopus oocyte, as an approach to understanding signal transduction protein function. Methods Mol. Biol. 2009, 518, 43–55. [Google Scholar]
- Truong, D.H.; Eghbal, M.A.; Hindmarsh, W.; Roth, S.H.; O’Brien, P.J. Molecular mechanisms of hydrogen sulfide toxicity. Drug Metab. Rev. 2006, 38, 733–744. [Google Scholar] [CrossRef]
- Eghbal, M.A.; Pennefather, P.S.; O’Brien, P.J. H2S cytotoxicity mechanism involves reactive oxygen species formation and mitochondrial depolarisation. Toxicology 2004, 203, 69–76. [Google Scholar] [CrossRef]
- Dehennaut, V.; Lefebvre, T.; Sellier, C.; Leroy, Y.; Gross, B.; Walker, S.; Cacan, R.; Michalski, J.-C.; Vilain, J.-P.; Bodar, J.-F. O-linked N-acetylglucosaminyltransferase inhibition prevents G2/M transition in Xenopus laevis oocytes. J. Biol. Chem. 2007, 282, 12527–12536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, O.; Morales, A.; Fernández-Checa, J.C.; Garcia-Ruiz, C. Neutral sphingomyelinase-induced ceramide triggers germinal vesicle breakdown and oxidant-dependent apoptosis in Xenopus laevis oocytes. J. Lipid Res. 2007, 48, 1924–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Ma, C.; Karmouch, J.; Katbi, H.A.; Liu, X.J. Antiapoptotic role for ornithine decarboxylase during oocyte maturation. Mol. Cell. Biol. 2009, 29, 1786–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Pasquier, D.; Dupré, A.; Jessus, C. Unfertilized Xenopus eggs die by Bad-dependent apoptosis under the control of Cdk1 and JNK. PLoS ONE 2011, 6, e23672. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, M.; Wong, F.L.; Fu, D.; Lau, H.Y.; Moochhala, S.M.; Moore, P.K. Role of hydrogen sulfide in acute pancreatitis and associated lung injury. FASEB J. 2005, 19, 623–625. [Google Scholar] [CrossRef]
- Soleymanlou, N.; Jurisica, I.; Nevo, O.; Ietta, F.; Zhang, X.; Zamudio, S.; Post, M.; Caniggia, I. Molecular evidence of placental hypoxia in preeclampsia. J. Clin. Endocrinol. Metab. 2005, 90, 4299–4308. [Google Scholar] [CrossRef] [Green Version]
- Reynhout, J.K.; Smith, L.D. Studies on the appearance and nature of a maturation-inducing factor in the cytoplasm of amphibian oocytes exposed to progesterone. Dev. Biol. 1974, 38, 394–400. [Google Scholar] [CrossRef]
- Drury, K.C.; Schorderet-Slatkine, S. Effects of cycloheximide on the “autocatalytic” nature of the maturation promoting factor (MPF) in oocytes of Xenopus laevis. Cell 1975, 4, 269–274. [Google Scholar] [CrossRef]
- Wasserman, W.J.; Masui, Y. Effects of cyclohexamide on a cytoplasmic factor initiating meiotic naturation in Xenopus oocytes. Exp. Cell Res. 1975, 91, 381–388. [Google Scholar] [CrossRef]
- Karaïskou, A.; Jessus, C.; Brassac, T.; Ozon, R. Phosphatase 2A and polo kinase, two antagonistic regulators of cdc25 activation and MPF auto-amplification. J. Cell Sci. 1999, 112, 3747–3756. [Google Scholar]
- Karaiskou, A.; Leprêtre, A.-C.; Pahlavan, G.; Du Pasquier, D.; Ozon, R.; Jessus, C. Polo-like kinase confers MPF autoamplification competence to growing Xenopus oocytes. Development 2004, 131, 1543–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrieu, A.; Brassac, T.; Galas, S.; Fisher, D.; Labbé, J.C.; Dorée, M. The Polo-like kinase Plx1 is a component of the MPF amplification loop at the G2/M-phase transition of the cell cycle in Xenopus eggs. J. Cell Sci. 1998, 111, 1751–1757. [Google Scholar] [PubMed]
- Hochegger, H.; Klotzbücher, A.; Kirk, J.; Howell, M.; le Guellec, K.; Fletcher, K.; Duncan, T.; Sohail, M.; Hunt, T. New B-type cyclin synthesis is required between meiosis I and II during Xenopus oocyte maturation. Development 2001, 128, 3795–3807. [Google Scholar] [PubMed]
- Dorée, M.; Hunt, T. From Cdc2 to Cdk1: When did the cell cycle kinase join its cyclin partner? J. Cell Sci. 2002, 115, 2461–2464. [Google Scholar]
- Kimura, H. Hydrogen sulfide induces cyclic AMP and modulates the NMDA receptor. Biochem. Biophys. Res. Commun. 2000, 267, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Iciek, M.; Kowalczyk-Pachel, D.; Bilska-Wilkosz, A.; Kwiecień, I.; Górny, M.; Włodek, L. S-sulfhydration as a cellular redox regulation. Biosci. Rep. 2015, 36, e00304. [Google Scholar] [CrossRef]
- Kimura, H. Physiological role of hydrogen sulfide and polysulfide in the central nervous system. Neurochem. Int. 2013, 63, 492–497. [Google Scholar] [CrossRef]
- Krishnan, N.; Fu, C.; Pappin, D.J.; Tonks, N.K. H2S-Induced sulfhydration of the phosphatase PTP1B and its role in the endoplasmic reticulum stress response. Sci. Signal. 2011, 4, ra86. [Google Scholar] [CrossRef] [Green Version]
- Viry, E.; Anwar, A.; Kirsch, G.; Jacob, C.; Diederich, M.; Bagrel, D. Antiproliferative effect of natural tetrasulfides in human breast cancer cells is mediated through the inhibition of the cell division cycle 25 phosphatases. Int. J. Oncol. 2011, 38, 1103–1111. [Google Scholar]
- Tomko, R.J.; Lazo, J.S. Multimodal control of Cdc25A by nitrosative stress. Cancer Res. 2008, 68, 7457–7465. [Google Scholar] [CrossRef] [Green Version]
- Foster, M.W.; Forrester, M.T.; Stamler, J.S. A protein microarray-based analysis of S-nitrosylation. Proc. Natl. Acad. Sci. USA 2009, 106, 18948–18953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumdar, U.; Biswas, P.; Subhra Sarkar, T.; Maiti, D.; Ghosh, S. Regulation of cell cycle and stress responses under nitrosative stress in Schizosaccharomyces pombe. Free Radic. Biol. Med. 2012, 52, 2186–2200. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, J. Redox regulation of the Cdc25 phosphatases. Antioxid. Redox Signal. 2005, 7, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, L.; Su, J.; Li, B.; Zhang, X.; Chen, T. Proteomic analysis of G2/M arrest triggered by natural borneol/curcumin in HepG2 cells, the importance of ROS-p53 pathway. J. Agric. Food Chem. 2015, 63, 6440–6449. [Google Scholar] [CrossRef]
- Peng, A.; Lewellyn, A.L.; Maller, J.L. DNA damage signaling in early Xenopus embryos. Cell Cycle 2008, 7, 3–6. [Google Scholar] [CrossRef]
- Berger, L.; Wilde, A. Glycolytic metabolites are critical modulators of oocyte maturation and viability. PLoS ONE 2013, 8, e77612. [Google Scholar] [CrossRef] [Green Version]
- Mishanina, T.V.; Libiad, M.; Banerjee, R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Olson, K.R.; Gao, Y.; DeLeon, E.R.; Arif, M.; Arif, F.; Arora, N.; Straub, K.D. Catalase as a sulfide-sulfur oxido-reductase: An ancient (and modern?) regulator of reactive sulfur species (RSS). Redox Biol. 2017, 12, 325–339. [Google Scholar] [CrossRef]
- Jeseta, M.J.; Bodart, J.-F.L. Aneuploidy in Health and Disease; Storchova, Z., Ed.; InTech Open: Rijeka, Croatia, 2012; Volume 10, pp. 193–216. [Google Scholar]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gelaude, A.; Slaby, S.; Cailliau, K.; Marin, M.; Lescuyer-Rousseau, A.; Molinaro, C.; Nevoral, J.; Kučerová-Chrpová, V.; Sedmikova, M.; Petr, J.; et al. Hydrogen Sulfide Impairs Meiosis Resumption in Xenopus laevis Oocytes. Cells 2020, 9, 237. https://doi.org/10.3390/cells9010237
Gelaude A, Slaby S, Cailliau K, Marin M, Lescuyer-Rousseau A, Molinaro C, Nevoral J, Kučerová-Chrpová V, Sedmikova M, Petr J, et al. Hydrogen Sulfide Impairs Meiosis Resumption in Xenopus laevis Oocytes. Cells. 2020; 9(1):237. https://doi.org/10.3390/cells9010237
Chicago/Turabian StyleGelaude, Armance, Sylvain Slaby, Katia Cailliau, Matthieu Marin, Arlette Lescuyer-Rousseau, Caroline Molinaro, Jan Nevoral, Veronica Kučerová-Chrpová, Marketa Sedmikova, Jaroslav Petr, and et al. 2020. "Hydrogen Sulfide Impairs Meiosis Resumption in Xenopus laevis Oocytes" Cells 9, no. 1: 237. https://doi.org/10.3390/cells9010237
APA StyleGelaude, A., Slaby, S., Cailliau, K., Marin, M., Lescuyer-Rousseau, A., Molinaro, C., Nevoral, J., Kučerová-Chrpová, V., Sedmikova, M., Petr, J., Martoriati, A., & Bodart, J. -F. (2020). Hydrogen Sulfide Impairs Meiosis Resumption in Xenopus laevis Oocytes. Cells, 9(1), 237. https://doi.org/10.3390/cells9010237