IL-12 and IL-23—Close Relatives with Structural Homologies but Distinct Immunological Functions

Abstract
:1. Introduction
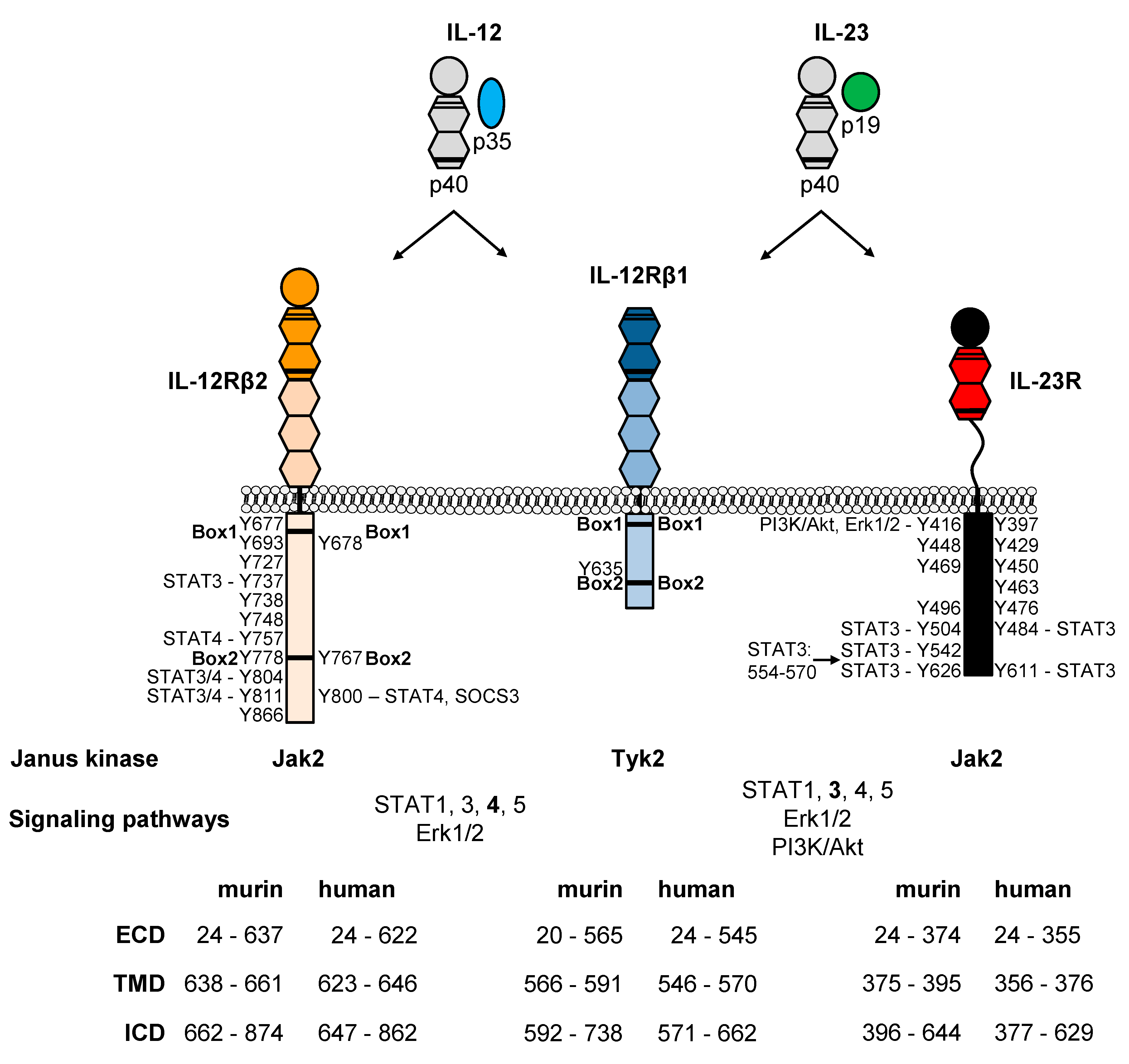
2. Structural Features of IL-12, IL-23 and Their Receptors
3. Insights into IL-12 and IL-23 Signal Transduction
4. Disease and Functional Consequences of Single Nucleotide Polymorphisms in the IL-23R
4.1. Internal SNP Number 03; rs1884444; Q3H; 43 Publications
4.2. Internal SNP Number 58; rs76575803; R86Q; 1 Publication
4.3. Internal SNP Number 93; rs76418789; G149R; 8 Publications
4.4. Internal SNP Number 109; rs371531867; Y173H; 1 Publication
4.5. Internal SNP Number 111; rs11465797; T175N; 1 Publication
4.6. Internal SNP Number 198; rs7530511; L310P; 39 Publications
4.7. Internal SNP Number 231; rs41313262; V362I; 4 Publications
4.8. Internal SNP Number 241; rs11209026; R381Q; 210 Publications
5. Conclusions and Perspectives for New Targeting Strategies
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Spangler, J.B.; Moraga, I.; Mendoza, J.L.; Garcia, K.C. Insights into cytokine-receptor interactions from cytokine engineering. Annu. Rev. Immunol. 2015, 33, 139–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose-John, S. Interleukin-6 family cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floss, D.M.; Scheller, J. Naturally occurring and synthetic constitutive-active cytokine receptors in disease and therapy. Cytokine Growth Factor Rev. 2019, 47, 1–20. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.; Gadina, M.; Siegel, R.M. Cytokines and cytokine receptors. In Clinical Immunology, Priciples and Practice, 5th ed.; Rich, R.R., Fleisher, T.A., Shearer, W.T., Schroeder, H.W., Frew, A.J., Weyand, C.M., Eds.; Elsevier: London, UK, 2018; pp. 127–155. [Google Scholar]
- Murakami, M.; Kamimura, D.; Hirano, T. Pleiotropy and specificity: Insights from the Interleukin 6 family of cytokines. Immunity 2019, 50, 812–831. [Google Scholar] [CrossRef] [Green Version]
- Tait Wojno, E.D.; Hunter, C.A.; Stumhofer, J.S. The Immunobiology of the Interleukin-12 family: Room for discovery. Immunity 2019, 50, 851–870. [Google Scholar] [CrossRef]
- Wang, R.X.; Yu, C.R.; Mahdi, R.M.; Egwuagu, C.E. Novel IL27p28/IL12p40 cytokine suppressed experimental autoimmune uveitis by inhibiting autoreactive Th1/Th17 cells and promoting expansion of regulatory T cells. J. Biol. Chem. 2012, 287, 36012–36021. [Google Scholar] [CrossRef] [Green Version]
- Flores, R.R.; Kim, E.; Zhou, L.; Yang, C.; Zhao, J.; Gambotto, A.; Robbins, P.D. IL-Y, a synthetic member of the IL-12 cytokine family, suppresses the development of type 1 diabetes in NOD mice. Eur. J. Immunol. 2015, 45, 3114–3125. [Google Scholar] [CrossRef]
- Oppmann, B.; Lesley, R.; Blom, B.; Timans, J.C.; Xu, Y.; Hunte, B.; Vega, F.; Yu, N.; Wang, J.; Singh, K.; et al. Novel p19 protein engages IL-12p40 to form a cytokine, IL-23, with biological activities similar as well as distinct from IL-12. Immunity 2000, 13, 715–725. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Fitz, L.; Ryan, M.; Hewick, R.M.; Clark, S.C.; Chan, S.; Loudon, R.; Sherman, F.; Perussia, B.; Trinchieri, G. Identification and purification of natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologic effects on human lymphocytes. J. Exp. Med. 1989, 170, 827–845. [Google Scholar] [CrossRef]
- Wolf, S.F.; Temple, P.A.; Kobayashi, M.; Young, D.; Dicig, M.; Lowe, L.; Dzialo, R.; Fitz, L.; Ferenz, C.; Hewick, R.M.; et al. Cloning of cDNA for natural killer cell stimulatory factor, a heterodimeric cytokine with multiple biologic effects on T and natural killer cells. J. Immunol. 1991, 146, 3074–3081. [Google Scholar]
- Stern, A.S.; Podlaski, F.J.; Hulmes, J.D.; Pan, Y.C.; Quinn, P.M.; Wolitzky, A.G.; Familletti, P.C.; Stremlo, D.L.; Truitt, T.; Chizzonite, R.; et al. Purification to homogeneity and partial characterization of cytotoxic lymphocyte maturation factor from human B-lymphoblastoid cells. Proc. Natl. Acad. Sci. USA 1990, 87, 6808–6812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubler, U.; Chua, A.O.; Schoenhaut, D.S.; Dwyer, C.M.; McComas, W.; Motyka, R.; Nabavi, N.; Wolitzky, A.G.; Quinn, P.M.; Familletti, P.C.; et al. Coexpression of two distinct genes is required to generate secreted bioactive cytotoxic lymphocyte maturation factor. Proc. Natl. Acad. Sci. USA 1991, 88, 4143–4147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenhaut, D.S.; Chua, A.O.; Wolitzky, A.G.; Quinn, P.M.; Dwyer, C.M.; McComas, W.; Familletti, P.C.; Gately, M.K.; Gubler, U. Cloning and expression of murine IL-12. J. Immunol. 1992, 148, 3433–3440. [Google Scholar] [PubMed]
- Ramnath, D.; Tunny, K.; Hohenhaus, D.M.; Pitts, C.M.; Bergot, A.S.; Hogarth, P.M.; Hamilton, J.A.; Kapetanovic, R.; Sturm, R.A.; Scholz, G.M.; et al. TLR3 drives IRF6-dependent IL-23p19 expression and p19/EBI3 heterodimer formation in keratinocytes. Immunol Cell. Biol. 2015, 93, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wei, Y.; Xiao, H.; Liu, X.; Zhang, Y.; Han, G.; Chen, G.; Hou, C.; Ma, N.; Shen, B.; et al. A novel IL-23p19/Ebi3 (IL-39) cytokine mediates inflammation in Lupus-like mice. Eur. J. Immunol. 2016, 46, 1343–1350. [Google Scholar] [CrossRef] [Green Version]
- Bridgewood, C.; Alase, A.; Watad, A.; Wittmann, M.; Cuthbert, R.; McGonagle, D. The IL-23p19/EBI3 heterodimeric cytokine termed IL-39 remains a theoretical cytokine in man. Inflamm. Res. 2019, 68, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Pflanz, S.; Timans, J.C.; Cheung, J.; Rosales, R.; Kanzler, H.; Gilbert, J.; Hibbert, L.; Churakova, T.; Travis, M.; Vaisberg, E.; et al. IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein, induces proliferation of naive CD4+ T cells. Immunity 2002, 16, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Devergne, O.; Birkenbach, M.; Kieff, E. Epstein-Barr virus-induced gene 3 and the p35 subunit of interleukin 12 form a novel heterodimeric hematopoietin. Proc. Natl. Acad. Sci. USA. 1997, 94, 12041–12046. [Google Scholar] [CrossRef] [Green Version]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef]
- Ling, P.; Gately, M.K.; Gubler, U.; Stern, A.S.; Lin, P.; Hollfelder, K.; Su, C.; Pan, Y.C.; Hakimi, J. Human IL-12 p40 homodimer binds to the IL-12 receptor but does not mediate biologic activity. J. Immunol. 1995, 154, 116–127. [Google Scholar]
- Crabe, S.; Guay-Giroux, A.; Tormo, A.J.; Duluc, D.; Lissilaa, R.; Guilhot, F.; Mavoungou-Bigouagou, U.; Lefouili, F.; Cognet, I.; Ferlin, W.; et al. The IL-27 p28 subunit binds cytokine-like factor 1 to form a cytokine regulating NK and T cell activities requiring IL-6R for signaling. J. Immunol. 2009, 183, 7692–7702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdi, K.; Singh, N.J.; Spooner, E.; Kessler, B.M.; Radaev, S.; Lantz, L.; Xiao, T.S.; Matzinger, P.; Sun, P.D.; Ploegh, H.L. Free IL-12p40 monomer is a polyfunctional adaptor for generating novel IL-12-like heterodimers extracellularly. J. Immunol. 2014, 192, 6028–6036. [Google Scholar] [CrossRef] [PubMed]
- Detry, S.; Skladanowska, K.; Vuylsteke, M.; Savvides, S.N.; Bloch, Y. Revisiting the combinatorial potential of cytokine subunits in the IL-12 family. Biochem. Pharmacol. 2019, 165, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Chehboun, S.; Labrecque-Carbonneau, J.; Pasquin, S.; Meliani, Y.; Meddah, B.; Ferlin, W.; Sharma, M.; Tormo, A.; Masson, J.F.; Gauchat, J.F. Epstein-Barr virus-induced gene 3 (EBI3) can mediate IL-6 trans-signaling. J. Biol. Chem. 2017, 292, 6644–6656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbers, C.; Spudy, B.; Aparicio-Siegmund, S.; Waetzig, G.H.; Sommer, J.; Holscher, C.; Rose-John, S.; Grotzinger, J.; Lorenzen, I.; Scheller, J. An interleukin-6 receptor-dependent molecular switch mediates signal transduction of the IL-27 cytokine subunit p28 (IL-30) via a gp130 protein receptor homodimer. J. Biol. Chem. 2013, 288, 4346–4354. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, H.; Mizoguchi, I.; Chiba, Y.; Ohashi, M.; Xu, M.; Yoshimoto, T. Expanding diversity in molecular structures and functions of the IL-6/IL-12 heterodimeric cytokine family. Front. Immunol. 2016, 7, 479. [Google Scholar] [CrossRef] [Green Version]
- Floss, D.M.; Schonberg, M.; Franke, M.; Horstmeier, F.C.; Engelowski, E.; Schneider, A.; Rosenfeldt, E.M.; Scheller, J. IL-6/IL-12 cytokine receptor shuffling of extra- and intracellular domains reveals canonical STAT activation via synthetic IL-35 and IL-39 signaling. Sci. Rep. 2017, 7, 15172. [Google Scholar] [CrossRef]
- Rose-John, S.; Scheller, J.; Schaper, F. “Family reunion”--A structured view on the composition of the receptor complexes of interleukin-6-type and interleukin-12-type cytokines. Cytokine Growth Factor Rev. 2015, 26, 471–474. [Google Scholar] [CrossRef]
- Cua, D.J.; Sherlock, J.; Chen, Y.; Murphy, C.A.; Joyce, B.; Seymour, B.; Lucian, L.; To, W.; Kwan, S.; Churakova, T.; et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 2003, 421, 744–748. [Google Scholar] [CrossRef]
- Zhang, G.X.; Gran, B.; Yu, S.; Li, J.; Siglienti, I.; Chen, X.; Kamoun, M.; Rostami, A. Induction of experimental autoimmune encephalomyelitis in IL-12 receptor-beta 2-deficient mice: IL-12 responsiveness is not required in the pathogenesis of inflammatory demyelination in the central nervous system. J. Immunol. 2003, 170, 2153–2160. [Google Scholar] [CrossRef]
- Zhang, G.X.; Yu, S.; Gran, B.; Li, J.; Siglienti, I.; Chen, X.; Calida, D.; Ventura, E.; Kamoun, M.; Rostami, A. Role of IL-12 receptor beta 1 in regulation of T cell response by APC in experimental autoimmune encephalomyelitis. J. Immunol. 2003, 171, 4485–4492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khader, S.A.; Thirunavukkarasu, S. The tale of IL-12 and IL-23: A paradigm shift. J. Immunol. 2019, 202, 629–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croxford, A.L.; Kulig, P.; Becher, B. IL-12-and IL-23 in health and disease. Cytokine Growth Factor Rev. 2014, 25, 415–421. [Google Scholar] [CrossRef]
- Moschen, A.R.; Tilg, H.; Raine, T. IL-12, IL-23 and IL-17 in IBD: Immunobiology and therapeutic targeting. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Wang, Y.; Wang, Z.; Liu, L.; Yang, Z.; Wang, M.; Xu, Y.; Ye, D.; Zhang, J.; Lin, Y.; et al. Roles and mechanisms of Interleukin-12 family members in cardiovascular diseases: Opportunities and challenges. Front. Pharmacol. 2020, 11, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Smyth, M.J.; Teng, M.W.L. Interleukin (IL)-12 and IL-23 and their conflicting roles in cancer. Cold Spring Harb. Perspect. Biol. 2018, 10, a028530. [Google Scholar] [CrossRef]
- Guo, Y.; Cao, W.; Zhu, Y. Immunoregulatory functions of the IL-12 family of cytokines in antiviral systems. Viruses 2019, 11, 772. [Google Scholar] [CrossRef] [Green Version]
- Sieper, J.; Poddubnyy, D.; Miossec, P. The IL-23-IL-17 pathway as a therapeutic target in axial spondyloarthritis. Nat. Rev. Rheumatol. 2019, 15, 747–757. [Google Scholar] [CrossRef]
- Neurath, M.F. IL-23 in inflammatory bowel diseases and colon cancer. Cytokine Growth Factor Rev. 2019, 45, 1–8. [Google Scholar] [CrossRef]
- Zundler, S.; Neurath, M.F. Interleukin-12: Functional activities and implications for disease. Cytokine Growth Factor Rev. 2015, 26, 559–568. [Google Scholar] [CrossRef]
- Chyuan, I.T.; Lai, J.H. New insights into the IL-12 and IL-23: From a molecular basis to clinical application in immune-mediated inflammation and cancers. Biochem. Pharmacol. 2020, 175, 113928. [Google Scholar] [CrossRef] [PubMed]
- Wong, U.; Cross, R.K. Expert opinion on interleukin-12/23 and interleukin-23 antagonists as potential therapeutic options for the treatment of inflammatory bowel disease. Expert Opin. Investig. Drugs 2019, 28, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Zabotti, A.; Goletti, D.; Lubrano, E.; Cantini, F. The impact of the interleukin 12/23 inhibitor ustekinumab on the risk of infections in patients with psoriatic arthritis. Expert Opin. Drug Saf. 2020, 19, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Jeon, C.; Sekhon, S.; Yan, D.; Afifi, L.; Nakamura, M.; Bhutani, T. Monoclonal antibodies inhibiting IL-12, -23, and -17 for the treatment of psoriasis. Hum. Vaccin. Immunother. 2017, 13, 2247–2259. [Google Scholar] [CrossRef] [Green Version]
- Yuan, N.; Yu, G.; Liu, D.; Wang, X.; Zhao, L. An emerging role of interleukin-23 in rheumatoid arthritis. Immunopharmacol. Immunotoxicol. 2019, 41, 185–191. [Google Scholar] [CrossRef]
- Sospedra, M.; Martin, R. Immunology of Multiple Sclerosis. Semin. Neurol. 2016, 36, 115–127. [Google Scholar] [CrossRef]
- Tugues, S.; Burkhard, S.H.; Ohs, I.; Vrohlings, M.; Nussbaum, K.; Vom Berg, J.; Kulig, P.; Becher, B. New insights into IL-12-mediated tumor suppression. Cell Death Differ. 2015, 22, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Egwuagu, C.E.; Yu, C.R.; Sun, L.; Wang, R. Interleukin 35: Critical regulator of immunity and lymphocyte-mediated diseases. Cytokine Growth Factor Rev. 2015, 26, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Parham, C.; Chirica, M.; Timans, J.; Vaisberg, E.; Travis, M.; Cheung, J.; Pflanz, S.; Zhang, R.; Singh, K.P.; Vega, F.; et al. A receptor for the heterodimeric cytokine IL-23 is composed of IL-12Rbeta1 and a novel cytokine receptor subunit, IL-23R. J. Immunol. 2002, 168, 5699–5708. [Google Scholar] [CrossRef] [Green Version]
- Yoon, C.; Johnston, S.C.; Tang, J.; Stahl, M.; Tobin, J.F.; Somers, W.S. Charged residues dominate a unique interlocking topography in the heterodimeric cytokine interleukin-12. EMBO J. 2000, 19, 3530–3541. [Google Scholar] [CrossRef]
- Heinzel, F.P.; Hujer, A.M.; Ahmed, F.N.; Rerko, R.M. In vivo production and function of IL-12 p40 homodimers. J. Immunol. 1997, 158, 4381–4388. [Google Scholar] [PubMed]
- Jalah, R.; Rosati, M.; Ganneru, B.; Pilkington, G.R.; Valentin, A.; Kulkarni, V.; Bergamaschi, C.; Chowdhury, B.; Zhang, G.M.; Beach, R.K.; et al. The p40 subunit of interleukin (IL)-12 promotes stabilization and export of the p35 subunit: Implications for improved IL-12 cytokine production. J. Biol. Chem. 2013, 288, 6763–6776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Andrea, A.; Rengaraju, M.; Valiante, N.M.; Chehimi, J.; Kubin, M.; Aste, M.; Chan, S.H.; Kobayashi, M.; Young, D.; Nickbarg, E.; et al. Production of natural killer cell stimulatory factor (interleukin 12) by peripheral blood mononuclear cells. J. Exp. Med. 1992, 176, 1387–1398. [Google Scholar] [CrossRef]
- Reitberger, S.; Haimerl, P.; Aschenbrenner, I.; Esser-von Bieren, J.; Feige, M.J. Assembly-induced folding regulates interleukin 12 biogenesis and secretion. J. Biol. Chem. 2017, 292, 8073–8081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparicio-Siegmund, S.; Moll, J.M.; Lokau, J.; Grusdat, M.; Schroder, J.; Plohn, S.; Rose-John, S.; Grotzinger, J.; Lang, P.A.; Scheller, J.; et al. Recombinant p35 from bacteria can form Interleukin (IL-)12, but Not IL-35. PLoS ONE 2014, 9, e107990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.L.; Chaturvedi, V.; Uyttenhove, C.; Van Snick, J.; Vignali, D.A. Distinct subunit pairing criteria within the heterodimeric IL-12 cytokine family. Mol. Immunol. 2012, 51, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.M.; Khader, S.A. IL-12p40: An inherently agonistic cytokine. Trends. Immunol. 2007, 28, 33–38. [Google Scholar] [CrossRef]
- Mattner, F.; Fischer, S.; Guckes, S.; Jin, S.; Kaulen, H.; Schmitt, E.; Rude, E.; Germann, T. The interleukin-12 subunit p40 specifically inhibits effects of the interleukin-12 heterodimer. Eur. J. Immunol. 1993, 23, 2202–2208. [Google Scholar] [CrossRef] [PubMed]
- Gillessen, S.; Carvajal, D.; Ling, P.; Podlaski, F.J.; Stremlo, D.L.; Familletti, P.C.; Gubler, U.; Presky, D.H.; Stern, A.S.; Gately, M.K. Mouse interleukin-12 (IL-12) p40 homodimer: A potent IL-12 antagonist. Eur. J. Immunol. 1995, 25, 200–206. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jung, Y.O.; Kim, D.J.; Kang, C.M.; Moon, Y.M.; Heo, Y.J.; Oh, H.J.; Park, S.J.; Yang, S.H.; Kwok, S.K.; et al. IL-12p40 homodimer ameliorates experimental autoimmune arthritis. J. Immunol. 2015, 195, 3001–3010. [Google Scholar] [CrossRef] [Green Version]
- Shimozato, O.; Ugai, S.; Chiyo, M.; Takenobu, H.; Nagakawa, H.; Wada, A.; Kawamura, K.; Yamamoto, H.; Tagawa, M. The secreted form of the p40 subunit of interleukin (IL)-12 inhibits IL-23 functions and abrogates IL-23-mediated antitumour effects. J. Immunol. 2006, 117, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Russell, T.D.; Yan, Q.; Fan, G.; Khalifah, A.P.; Bishop, D.K.; Brody, S.L.; Walter, M.J. IL-12 p40 homodimer-dependent macrophage chemotaxis and respiratory viral inflammation are mediated through IL-12 receptor beta 1. J. Immunol. 2003, 171, 6866–6874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, S.J.; Lee, C.H.; Lee, S.B.; Kim, C.M.; Jang, K.L.; Shin, H.S.; Sung, Y.C. A novel function of IL-12p40 as a chemotactic molecule for macrophages. J. Immunol. 1999, 163, 2902–2908. [Google Scholar] [PubMed]
- Jana, M.; Dasgupta, S.; Saha, R.N.; Liu, X.; Pahan, K. Induction of tumor necrosis factor-alpha (TNF-alpha) by interleukin-12 p40 monomer and homodimer in microglia and macrophages. J. Neurochem. 2003, 86, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jana, M.; Pahan, K. Induction of lymphotoxin-alpha by interleukin-12 p40 homodimer, the so-called biologically inactive molecule, but not IL-12 p70. Immunology 2009, 127, 312–325. [Google Scholar] [CrossRef]
- Jana, M.; Dasgupta, S.; Pal, U.; Pahan, K. IL-12 p40 homodimer, the so-called biologically inactive molecule, induces nitric oxide synthase in microglia via IL-12R beta 1. Glia 2009, 57, 1553–1565. [Google Scholar] [CrossRef] [Green Version]
- Schroder, J.; Moll, J.M.; Baran, P.; Grotzinger, J.; Scheller, J.; Floss, D.M. Non-canonical interleukin 23 receptor complex assembly: p40 protein recruits interleukin 12 receptor beta1 via site II and induces p19/interleukin 23 receptor interaction via site III. J. Biol. Chem. 2015, 290, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Beyer, B.M.; Ingram, R.; Ramanathan, L.; Reichert, P.; Le, H.V.; Madison, V.; Orth, P. Crystal structures of the pro-inflammatory cytokine interleukin-23 and its complex with a high-affinity neutralizing antibody. J. Mol. Biol. 2008, 382, 942–955. [Google Scholar] [CrossRef]
- Lupardus, P.J.; Garcia, K.C. The structure of interleukin-23 reveals the molecular basis of p40 subunit sharing with interleukin-12. J. Mol. Biol. 2008, 382, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Meier, S.; Bohnacker, S.; Klose, C.J.; Lopez, A.; Choe, C.A.; Schmid, P.W.N.; Bloemeke, N.; Ruhrnossl, F.; Haslbeck, M.; Bieren, J.E.; et al. The molecular basis of chaperone-mediated interleukin 23 assembly control. Nat. Commun. 2019, 10, 4121. [Google Scholar] [CrossRef] [Green Version]
- Espigol-Frigole, G.; Planas-Rigol, E.; Ohnuki, H.; Salvucci, O.; Kwak, H.; Ravichandran, S.; Luke, B.; Cid, M.C.; Tosato, G. Identification of IL-23p19 as an endothelial proinflammatory peptide that promotes gp130-STAT3 signaling. Sci. Signal. 2016, 9, ra28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, M.J.; Chow, D.C.; Brevnova, E.E.; Garcia, K.C. Hexameric structure and assembly of the interleukin-6/IL-6 alpha-receptor/gp130 complex. Science 2003, 300, 2101–2104. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.L.; Vignali, D.A. Molecular interactions within the IL-6/IL-12 cytokine/receptor superfamily. Immunol. Res. 2011, 51, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignali, D.A.; Kuchroo, V.K. IL-12 family cytokines: Immunological playmakers. Nat. Immunol. 2012, 13, 722–728. [Google Scholar] [CrossRef] [Green Version]
- Collison, L.W.; Delgoffe, G.M.; Guy, C.S.; Vignali, K.M.; Chaturvedi, V.; Fairweather, D.; Satoskar, A.R.; Garcia, K.C.; Hunter, C.A.; Drake, C.G.; et al. The composition and signaling of the IL-35 receptor are unconventional. Nat. Immunol. 2012, 13, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, B.C.; Ultsch, M.; De Vos, A.M.; Mulkerrin, M.G.; Clauser, K.R.; Wells, J.A. Dimerization of the extracellular domain of the human growth hormone receptor by a single hormone molecule. Science 1991, 254, 821–825. [Google Scholar] [CrossRef]
- de Vos, A.M.; Ultsch, M.; Kossiakoff, A.A. Human growth hormone and extracellular domain of its receptor: Crystal structure of the complex. Science 1992, 255, 306–312. [Google Scholar] [CrossRef]
- Hummel, T.M.; Ackfeld, T.; Schonberg, M.; Ciupka, G.; Schulz, F.; Oberdoerster, A.; Grotzinger, J.; Scheller, J.; Floss, D.M. Synthetic deletion of the Interleukin 23 receptor (IL-23R) stalk region led to autonomous IL-23R homodimerization and activation. Mol. Cell. Biol. 2017, 37, e00014–e00017. [Google Scholar] [CrossRef] [Green Version]
- Siupka, P.; Hamming, O.T.; Kang, L.; Gad, H.H.; Hartmann, R. A conserved sugar bridge connected to the WSXWS motif has an important role for transport of IL-21R to the plasma membrane. Genes Immun. 2015, 16, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Chua, A.O.; Chizzonite, R.; Desai, B.B.; Truitt, T.P.; Nunes, P.; Minetti, L.J.; Warrier, R.R.; Presky, D.H.; Levine, J.F.; Gately, M.K.; et al. Expression cloning of a human IL-12 receptor component. A new member of the cytokine receptor superfamily with strong homology to gp130. J. Immunol. 1994, 153, 128–136. [Google Scholar]
- Chua, A.O.; Wilkinson, V.L.; Presky, D.H.; Gubler, U. Cloning and characterization of a mouse IL-12 receptor-beta component. J. Immunol. 1995, 155, 4286–4294. [Google Scholar] [PubMed]
- Presky, D.H.; Yang, H.; Minetti, L.J.; Chua, A.O.; Nabavi, N.; Wu, C.Y.; Gately, M.K.; Gubler, U. A functional interleukin 12 receptor complex is composed of two beta-type cytokine receptor subunits. Proc. Natl. Acad. Sci. USA 1996, 93, 14002–14007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayago, C.; Gonzalez Valcarcel, I.C.; Qian, Y.; Lee, J.; Alsina-Fernandez, J.; Fite, N.C.; Carrillo, J.J.; Zhang, F.F.; Chalmers, M.J.; Dodge, J.A.; et al. Deciphering binding interactions of IL-23R with HDX-MS: Mapping protein and macrocyclic dodecapeptide ligands. ACS Med. Chem. Lett. 2018, 9, 912–916. [Google Scholar] [CrossRef] [PubMed]
- Paonessa, G.; Graziani, R.; De Serio, A.; Savino, R.; Ciapponi, L.; Lahm, A.; Salvati, A.L.; Toniatti, C.; Ciliberto, G. Two distinct and independent sites on IL-6 trigger gp 130 dimer formation and signalling. EMBO J. 1995, 14, 1942–1951. [Google Scholar] [CrossRef] [PubMed]
- Plun-Favreau, H.; Perret, D.; Diveu, C.; Froger, J.; Chevalier, S.; Lelievre, E.; Gascan, H.; Chabbert, M. Leukemia inhibitory factor (LIF), cardiotrophin-1, and oncostatin M share structural binding determinants in the immunoglobulin-like domain of LIF receptor. J. Biol. Chem. 2003, 278, 27169–27179. [Google Scholar] [CrossRef] [Green Version]
- Barton, V.A.; Hudson, K.R.; Heath, J.K. Identification of three distinct receptor binding sites of murine interleukin-11. J. Biol. Chem. 1999, 274, 5755–5761. [Google Scholar] [CrossRef] [Green Version]
- Ciapponi, L.; Graziani, R.; Paonessa, G.; Lahm, A.; Ciliberto, G.; Savino, R. Definition of a composite binding site for gp130 in human interleukin-6. J Biol. Chem. 1995, 270, 31249–31254. [Google Scholar] [CrossRef] [Green Version]
- Bloch, Y.; Bouchareychas, L.; Merceron, R.; Składanowska, K.; Van den Bossche, L.; Detry, S.; Govindarajan, S.; Elewaut, D.; Haerynck, F.; Dullaers, M.; et al. Structural activation of pro-inflammatory human cytokine IL-23 by cognate IL-23 receptor enables recruitment of the shared receptor IL-12Rβ1. Immunity 2018, 48, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Esch, A.; Masiarz, A.; Mossner, S.; Moll, J.M.; Grotzinger, J.; Schroder, J.; Scheller, J.; Floss, D.M. Deciphering site 3 interactions of interleukin 12 and interleukin 23 with their cognate murine and human receptors. J. Biol. Chem. 2020, 295, 10478–10492. [Google Scholar] [CrossRef]
- Wang, X.; Wilkinson, V.L.; Podlaski, F.J.; Wu, C.; Stern, A.S.; Presky, D.H.; Magram, J. Characterization of mouse interleukin-12 p40 homodimer binding to the interleukin-12 receptor subunits. Eur. J. Immunol. 1999, 29, 2007–2013. [Google Scholar] [CrossRef]
- Chognard, G.; Bellemare, L.; Pelletier, A.N.; Dominguez-Punaro, M.C.; Beauchamp, C.; Guyon, M.J.; Charron, G.; Morin, N.; Sivanesan, D.; Kuchroo, V.; et al. The dichotomous pattern of IL-12r and IL-23R expression elucidates the role of IL-12 and IL-23 in inflammation. PLoS ONE 2014, 9, e89092. [Google Scholar] [CrossRef] [PubMed]
- Welsby, I.; Goriely, S. Regulation of Interleukin-23 expression in health and disease. Adv. Exp. Med. Biol. 2016, 941, 167–189. [Google Scholar] [PubMed]
- Burkett, P.R.; Meyer zu Horste, G.; Kuchroo, V.K. Pouring fuel on the fire: Th17 cells, the environment, and autoimmunity. J. Clin. Investig. 2015, 125, 2211–2219. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. T helper cell differentiation, heterogeneity, and plasticity. Cold Spring Harb. Perspect. Biol. 2018, 10, a030338. [Google Scholar] [CrossRef] [Green Version]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Ban, Y.; Wei, F.; Ma, X. Regulation of Interleukin-12 production in antigen-presenting cells. Adv. Exp. Med. Biol. 2016, 941, 117–138. [Google Scholar]
- Bacon, C.M.; Petricoin, E.F., 3rd; Ortaldo, J.R.; Rees, R.C.; Larner, A.C.; Johnston, J.A.; O’Shea, J.J. Interleukin 12 induces tyrosine phosphorylation and activation of STAT4 in human lymphocytes. Proc. Natl. Acad. Sci. USA 1995, 92, 7307–7311. [Google Scholar] [CrossRef] [Green Version]
- Villarino, A.V.; Kanno, Y.; O’Shea, J.J. Mechanisms and consequences of Jak-STAT signaling in the immune system. Nat. Immunol. 2017, 18, 374–384. [Google Scholar] [CrossRef]
- Oyamada, A.; Ikebe, H.; Itsumi, M.; Saiwai, H.; Okada, S.; Shimoda, K.; Iwakura, Y.; Nakayama, K.I.; Iwamoto, Y.; Yoshikai, Y.; et al. Tyrosine kinase 2 plays critical roles in the pathogenic CD4 T cell responses for the development of experimental autoimmune encephalomyelitis. J. Immunol. 2009, 183, 7539–7546. [Google Scholar] [CrossRef] [Green Version]
- Ishizaki, M.; Akimoto, T.; Muromoto, R.; Yokoyama, M.; Ohshiro, Y.; Sekine, Y.; Maeda, H.; Shimoda, K.; Oritani, K.; Matsuda, T. Involvement of tyrosine kinase-2 in both the IL-12/Th1 and IL-23/Th17 axes in vivo. J. Immunol. 2011, 187, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorman, J.A.; Hundhausen, C.; Kinsman, M.; Arkatkar, T.; Allenspach, E.J.; Clough, C.; West, S.E.; Thomas, K.; Eken, A.; Khim, S.; et al. The TYK2-P1104A autoimmune protective variant limits coordinate signals required to generate specialized T cell subsets. Front. Immunol. 2019, 10, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Presky, D.H.; Wu, C.Y.; Gubler, U. Differential associations between the cytoplasmic regions of the interleukin-12 receptor subunits beta1 and beta2 and JAK kinases. J. Biol. Chem. 1997, 272, 6073–6077. [Google Scholar] [CrossRef] [Green Version]
- Floss, D.M.; Klocker, T.; Schroder, J.; Lamertz, L.; Mrotzek, S.; Strobl, B.; Hermanns, H.; Scheller, J. Defining the functional binding sites of interleukin 12 receptor beta1 and interleukin 23 receptor to Janus kinases. Mol. Biol. Cell 2016, 27, 2301–2316. [Google Scholar] [CrossRef]
- Yamamoto, K.; Shibata, F.; Miura, O.; Kamiyama, R.; Hirosawa, S.; Miyasaka, N. Physical interaction between interleukin-12 receptor beta 2 subunit and Jak2 tyrosine kinase: Jak2 associates with cytoplasmic membrane-proximal region of interleukin-12 receptor beta 2 via amino-terminus. Biochem. Biophys. Res. Commun. 1999, 257, 400–404. [Google Scholar] [CrossRef]
- Neubauer, H.; Cumano, A.; Muller, M.; Wu, H.; Huffstadt, U.; Pfeffer, K. Jak2 deficiency defines an essential developmental checkpoint in definitive hematopoiesis. Cell 1998, 93, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, A.; Ito, M.; Chikuma, S.; Akanuma, T.; Nakatsukasa, H. Negative regulation of cytokine signaling in immunity. Cold Spring Harb. Perspect. Biol. 2018, 10, a028571. [Google Scholar] [CrossRef] [PubMed]
- Babon, J.J.; Varghese, L.N.; Nicola, N.A. Inhibition of IL-6 family cytokines by SOCS3. Semin. Immunol. 2014, 26, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The molecular details of cytokine signaling via the JAK/STAT pathway. Protein Sci. 2018, 27, 1984–2009. [Google Scholar] [CrossRef] [Green Version]
- Anhuf, D.; Weissenbach, M.; Schmitz, J.; Sobota, R.; Hermanns, H.M.; Radtke, S.; Linnemann, S.; Behrmann, I.; Heinrich, P.C.; Schaper, F. Signal transduction of IL-6, leukemia-inhibitory factor, and oncostatin M: Structural receptor requirements for signal attenuation. J. Immunol. 2000, 165, 2535–2543. [Google Scholar] [CrossRef] [Green Version]
- Floss, D.M.; Mrotzek, S.; Klocker, T.; Schroder, J.; Grotzinger, J.; Rose-John, S.; Scheller, J. Identification of canonical tyrosine-dependent and non-canonical tyrosine-independent STAT3 activation sites in the intracellular domain of the interleukin 23 receptor. J. Biol. Chem. 2013, 288, 19386–19400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lochmatter, C.; Fischer, R.; Charles, P.D.; Yu, Z.; Powrie, F.; Kessler, B.M. Integrative phosphoproteomics links IL-23R signaling with metabolic adaptation in lymphocytes. Sci. Rep. 2016, 6, 24491. [Google Scholar] [CrossRef]
- Alvarez-Salamero, C.; Castillo-Gonzalez, R.; Pastor-Fernandez, G.; Mariblanca, I.R.; Pino, J.; Cibrian, D.; Navarro, M.N. IL-23 signaling regulation of pro-inflammatory T-cell migration uncovered by phosphoproteomics. PLoS Biol. 2020, 18, e3000646. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Fernandez, G.; Mariblanca, I.R.; Navarro, M.N. Decoding IL-23 signaling cascades for new Therapeutic opportunities. Cells 2020, 9, 2044. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.L.; Kang, J.W.; Moon, Y.M.; Nam, H.J.; Jhun, J.Y.; Heo, S.B.; Jin, H.T.; Min, S.Y.; Ju, J.H.; Park, K.S.; et al. STAT3 and NF-kappaB signal pathway is required for IL-23-mediated IL-17 production in spontaneous arthritis animal model IL-1 receptor antagonist-deficient mice. J. Immunol. 2006, 176, 5652–5661. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Wang, X.; Gadina, M.; O’Shea, J.J.; Presky, D.H.; Magram, J. IL-12 receptor beta 2 (IL-12R beta 2)-deficient mice are defective in IL-12-mediated signaling despite the presence of high affinity IL-12 binding sites. J. Immunol. 2000, 165, 6221–6228. [Google Scholar] [CrossRef] [Green Version]
- Nishikomori, R.; Usui, T.; Wu, C.Y.; Morinobu, A.; O’Shea, J.J.; Strober, W. Activated STAT4 has an essential role in Th1 differentiation and proliferation that is independent of its role in the maintenance of IL-12R beta 2 chain expression and signaling. J. Immunol. 2002, 169, 4388–4398. [Google Scholar] [CrossRef]
- Naeger, L.K.; McKinney, J.; Salvekar, A.; Hoey, T. Identification of a STAT4 binding site in the interleukin-12 receptor required for signaling. J. Biol. Chem. 1999, 274, 1875–1878. [Google Scholar] [CrossRef] [Green Version]
- Yao, B.B.; Niu, P.; Surowy, C.S.; Faltynek, C.R. Direct interaction of STAT4 with the IL-12 receptor. Arch. Biochem. Biophys. 1999, 368, 147–155. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yamaguchi, M.; Miyasaka, N.; Miura, O. SOCS-3 inhibits IL-12-induced STAT4 activation by binding through its SH2 domain to the STAT4 docking site in the IL-12 receptor beta2 subunit. Biochem. Biophys. Res. Commun. 2003, 310, 1188–1193. [Google Scholar] [CrossRef]
- Eyles, J.L.; Metcalf, D.; Grusby, M.J.; Hilton, D.J.; Starr, R. Negative regulation of interleukin-12 signaling by suppressor of cytokine signaling-1. J. Biol. Chem. 2002, 277, 43735–43740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Laurence, A.; Kanno, Y.; Pacher-Zavisin, M.; Zhu, B.M.; Tato, C.; Yoshimura, A.; Hennighausen, L.; O’Shea, J.J. Selective regulatory function of Socs3 in the formation of IL-17-secreting T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 8137–8142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelowski, E.; Schneider, A.; Franke, M.; Xu, H.; Clemen, R.; Lang, A.; Baran, P.; Binsch, C.; Knebel, B.; Al-Hasani, H.; et al. Synthetic cytokine receptors transmit biological signals using artificial ligands. Nat. Commun. 2018, 9, 2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, A.; Lambert, Q.T.; Reuther, G.W. Transformation of hematopoietic cells and activation of JAK2-V617F by IL-27R, a component of a heterodimeric type I cytokine receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 18502–18507. [Google Scholar] [CrossRef] [Green Version]
- de Paus, R.; van de Wetering, D.; van Dissel, J.; van de Vosse, E. IL-23 and IL-12 responses in activated human T cells retrovirally transduced with IL-23 receptor variants. Mol. Immunol. 2008, 45, 3889–3895. [Google Scholar] [CrossRef]
- Kleinsteuber, K.; Heesch, K.; Schattling, S.; Sander-Juelch, C.; Mock, U.; Riecken, K.; Fehse, B.; Fleischer, B.; Jacobsen, M. SOCS3 promotes interleukin-17 expression of human T cells. Blood 2012, 120, 4374–4382. [Google Scholar] [CrossRef]
- Wu, C.; Yosef, N.; Thalhamer, T.; Zhu, C.; Xiao, S.; Kishi, Y.; Regev, A.; Kuchroo, V.K. Induction of pathogenic TH17 cells by inducible salt-sensing kinase SGK1. Nature 2013, 496, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Engelowski, E.; Modares, N.F.; Gorressen, S.; Bouvain, P.; Semmler, D.; Alter, C.; Ding, Z.; Flogel, U.; Schrader, J.; Xu, H.; et al. IL-23R signaling plays no role in myocardial infarction. Sci. Rep. 2018, 8, 17078. [Google Scholar] [CrossRef]
- Kumar, P.; Henikoff, S.; Ng, P. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protoc. 2009, 4, 1073–1081. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Ogola, G.; Ouma, C.; Jura, W.; Muok, E.; Colebunders, R.; Mwinzi, P. A non-synonymous polymorphism in IL-23R Gene (rs1884444) is associated with reduced risk to schistosomiasis-associated Immune Reconstitution Inflammatory Syndrome in a Kenyan population. BMC Infect. Dis. 2014, 14, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Lu, Y.; Zhang, H.; Ding, Y.; Ren, C.; Hua, Z.; Zhou, Y.; Deng, B.; Jin, G.; Hu, Z.; et al. A nonsynonymous polymorphism in IL23R gene is associated with risk of gastric cancer in a Chinese population. Mol. Carcinog. 2010, 49, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Cao, W.; Chen, W.; Pan, S.; Xiao, Y.; Liu, Y.; Gu, H.; Guo, W.; Xu, L.; Hu, Z.; et al. Potentially functional polymorphisms in IL-23 receptor and risk of esophageal cancer in a Chinese population. Int. J. Cancer 2012, 130, 1093–1097. [Google Scholar] [CrossRef]
- Safrany, E.; Szabo, M.; Szell, M.; Kemeny, L.; Sumegi, K.; Melegh, B.; Magyari, L.; Matyas, P.; Figler, M.; Weber, A.; et al. Difference of interleukin-23 receptor gene haplotype variants in ulcerative colitis compared to Crohn’s disease and psoriasis. Inflamm. Res. 2013, 62, 195–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faragó, B.; Magyari, L.; Sáfrány, E.; Csöngei, V.; Járomi, L.; Horvatovich, K.; Sipeky, C.; Maász, A.; Radics, J.; Gyetvai, A.; et al. Functional variants of interleukin-23 receptor gene confer risk for rheumatoid arthritis but not for systemic sclerosis. Ann. Rheum. Dis. 2007, 67, 248–250. [Google Scholar] [CrossRef]
- Momozawa, Y.; Mni, M.; Nakamura, K.; Coppieters, W.; Almer, S.; Amininejad, L.; Cleynen, I.; Colombel, J.; de Rijk, P.; Dewit, O.; et al. Resequencing of positional candidates identifies low frequency IL23R coding variants protecting against inflammatory bowel disease. Nat. Med. 2011, 43, 43–47. [Google Scholar] [CrossRef]
- Onodera, K.; Arimura, Y.; Isshiki, H.; Kawakami, K.; Nagaishi, K.; Yamashita, K.; Yamamoto, E.; Niinuma, T.; Naishiro, Y.; Suzuki, H.; et al. Low-frequency IL23R coding variant associated with Crohn’s disease susceptibility in Japanese subjects identified by personal genomics analysis. PLoS ONE 2015, 10, e0137801. [Google Scholar] [CrossRef]
- Hong, J.; Yang, H.; Moon, J.; Chang, J.; Ko, J. Association of IL23R variants with Crohn’s disease in Korean children. Front. Pediatr. 2019, 7, 472. [Google Scholar] [CrossRef] [Green Version]
- Sivanesan, D.; Beauchamp, C.; Quinou, C.; Lee, J.; Lesage, S.; Chemtob, S.; Rioux, J.; Michnick, S. IL23R (Interleukin 23 receptor) variants protective against inflammatory bowel diseases (IBD) display loss of function due to impaired protein stability and intracellular trafficking. J. Biol. Chem. 2016, 291, 8673–8685. [Google Scholar] [CrossRef] [Green Version]
- Takaku, T.; Calado, R.; Kajigaya, S.; Young, N. Interleukin-23 receptor (IL-23R) gene polymorphisms in acquired aplastic anemia. Ann. Hematol. 2009, 88, 653–657. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Kim, S.; Park, H.; Chung, J. Association of IL23R polymorphism (rs7530511) with intracerebral hemorrhage in Korean population. Neurol. Sci. 2016, 37, 983–985. [Google Scholar] [CrossRef] [PubMed]
- Duerr, R.; Taylor, K.; Brant, S.; Rioux, J.; Silverberg, M.; Daly, M.; Steinhart, A.; Abraham, C.; Regueiro, M.; Griffiths, A.; et al. A genome-wide association study identifies IL23R as an inflammatory bowel disease gene. Science 2006, 314, 1461–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cargill, M.; Schrodi, S.; Chang, M.; Garcia, V.; Brandon, R.; Callis, K.; Matsunami, N.; Ardlie, K.; Civello, D.; Catanese, J.; et al. A large-scale genetic association study confirms IL12B and leads to the identification of IL23R as psoriasis-risk genes. Am. J. Hum. Genet. 2007, 80, 273–290. [Google Scholar] [CrossRef] [Green Version]
- Huber, A.; Jacobson, E.; Jazdzewski, K.; Concepcion, E.; Tomer, Y. Interleukin (IL)-23 receptor is a major susceptibility gene for Graves’ ophthalmopathy: The IL-23/T-helper 17 axis extends to thyroid autoimmunity. J. Clin. Endocrinol. Metab. 2008, 93, 1077–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Jiang, H.; Chen, Z.; Lu, B.; Li, J.; Shen, X. Genetic association between IL23R rs11209026 and rs10889677 polymorphisms and risk of Crohn’s disease and ulcerative colitis: Evidence from 41 studies. Inflamm. Res. 2020, 69, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, H.; Kugathasan, S.; Annese, V.; Bradfield, J.; Russell, R.; Sleiman, P.; Imielinski, M.; Glessner, J.; Hou, C.; et al. Diverse genome-wide association studies associate the IL12/IL23 pathway with Crohn Disease. Am. J. Hum. Genet. 2009, 84, 399–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, L.; Wang, W.; Song, H. Complex role of IL-23R polymorphisms on ankylosing spondylitis: A meta-analysis. Expert Rev. Clin. Immunol. 2018, 14, 635–643. [Google Scholar] [CrossRef]
- Du, J.; Wang, X.; Tan, G.; Liang, Z.; Zhang, Z.; Yu, H. The association between genetic polymorphisms of interleukin 23 receptor gene and the risk of rheumatoid arthritis: An updated meta-analysis. Clin. Immunol. 2020, 210, 108250. [Google Scholar] [CrossRef]
- Pidasheva, S.; Trifari, S.; Phillips, A.; Hackney, J.; Ma, Y.; Smith, A.; Sohn, S.; Spits, H.; Little, R.; Behrens, T.; et al. Functional studies on the IBD susceptibility gene IL23R implicate reduced receptor function in the protective genetic variant R381Q. PLoS ONE 2011, 6, e25038. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, R.; Thirthar Palanivelu, V. Tildrakizumab for the treatment of psoriasis. Expert Rev. Clin. Immunol. 2019, 15, 5–12. [Google Scholar] [CrossRef]
- Soendergaard, C.; Bergenheim, F.H.; Bjerrum, J.T.; Nielsen, O.H. Targeting JAK-STAT signal transduction in IBD. Pharmacol. Ther. 2018, 192, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Gracey, E.; Hromadova, D.; Lim, M.; Qaiyum, Z.; Zeng, M.; Yao, Y.; Srinath, A.; Baglaenko, Y.; Yeremenko, N.; Westlin, W.; et al. TYK2 inhibition reduces type 3 immunity and modifies disease progression in murine spondyloarthritis. J. Clin. Investig. 2020, 130, 1863–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmet, J.; Verstraete, K.; Bloch, Y.; Lorent, E.; Wen, Y.; Devreese, B.; Vandenbroucke, K.; Loverix, S.; Hettmann, T.; Deroo, S.; et al. Structural basis of IL-23 antagonism by an Alphabody protein scaffold. Nat. Commun. 2014, 5, 5237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiniou, C.; Dominguez-Punaro, M.; Cloutier, F.; Erfani, A.; Ennaciri, J.; Sivanesan, D.; Sanchez, M.; Chognard, G.; Hou, X.; Rivera, J.C.; et al. Specific targeting of the IL-23 receptor, using a novel small peptide noncompetitive antagonist, decreases the inflammatory response. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R1216–R1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Taranath, R.; Mattheakis, L.; Bhandari, A.; Liu, D. The biomarker profile of PTG-200, an oral peptide antagonist of IL-23 receptor, tracks with efficacy in a preclinical model of IBD. Gastroenterology 2017, 152, S31. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α Subunits | ||||
|---|---|---|---|---|
| p19 | p28 | p35 | ||
| β subunits | p40 | IL-23 [9] | IL-Y [7,8] | IL-12 [10,11,12,13,14] |
| EBI3 | IL-39 [15,16,17] | IL-27 [18] | IL-35 [19,20] | |
| IL-12 | IL-23 | Ref. | |
|---|---|---|---|
| transcription regulators | T-bet, STAT4 | RORγT, STAT3 | [6] |
| T cell differentiation | TH1 cells | TH17 cells | [6] |
| cytokine induction | IFN-γ, TNFα | IL-17A, IL-17F, IL-22, TNFα | [35] |
| inflammation | pro-inflammatory cytokines, significant role of IL-12p40 | [43] | |
| IBD (UC and CD) | inflammation of gastrointestinal mucosa | [43] | |
| autoimmunity | important role of IL-12p40 | [44] | |
| psoriasis | IL-12p40 as therapeutic target | [45] | |
| RA | promotion of joint autoimmune inflammation | attenuation of joint autoimmune inflammation | [46] |
| multiple sclerosis | importance of multifunctional CD4+ T cells secreting IFN-γ, IL-17 and GM-CSF | [47] | |
| cancer | antitumor activity | tumor-promoting and tumor suppressing effects | [37,48] |
| infection | important role in cell-mediated immunity against bacteria, fungi and intracellular protozoa | role in immunity to extracellular bacterial infections and fungal infections | [44] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floss, D.M.; Moll, J.M.; Scheller, J. IL-12 and IL-23—Close Relatives with Structural Homologies but Distinct Immunological Functions. Cells 2020, 9, 2184. https://doi.org/10.3390/cells9102184
Floss DM, Moll JM, Scheller J. IL-12 and IL-23—Close Relatives with Structural Homologies but Distinct Immunological Functions. Cells. 2020; 9(10):2184. https://doi.org/10.3390/cells9102184
Chicago/Turabian StyleFloss, Doreen M., Jens M. Moll, and Jürgen Scheller. 2020. "IL-12 and IL-23—Close Relatives with Structural Homologies but Distinct Immunological Functions" Cells 9, no. 10: 2184. https://doi.org/10.3390/cells9102184
APA StyleFloss, D. M., Moll, J. M., & Scheller, J. (2020). IL-12 and IL-23—Close Relatives with Structural Homologies but Distinct Immunological Functions. Cells, 9(10), 2184. https://doi.org/10.3390/cells9102184