The Collagen-Based Medical Device MD-Tissue Acts as a Mechanical Scaffold Influencing Morpho-Functional Properties of Cultured Human Tenocytes

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
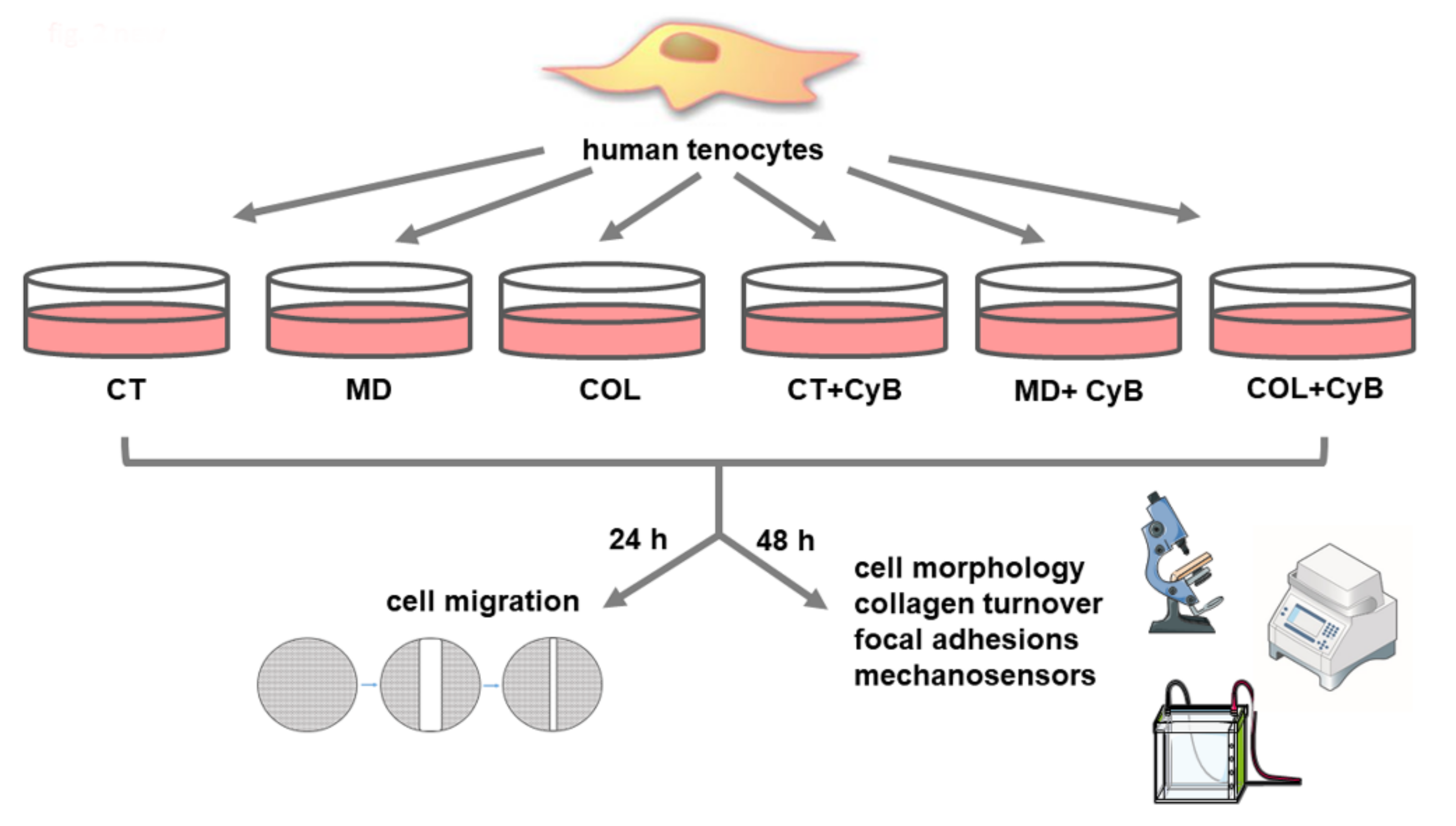
2. Materials and Methods
2.1. Samples
2.2. Cell Cultures
2.3. Coating with MD-Tissue or Collagen
2.4. Cytochalasin Administration
2.5. Scanning Electron Microscopy
2.6. Raman Spectroscopy
2.7. Immunofluorescence Analysis
2.8. Real-Time PCR
2.9. Slot Blot
2.10. Western Blot
2.11. SDS-Zymography
2.12. Wound Healing Assay
2.13. Statistical Analysis
3. Results
3.1. Analysis and Characterization of the Coating
3.2. Cell Morphology
3.3. Expression of Genes and Proteins Related to Collagen Turnover
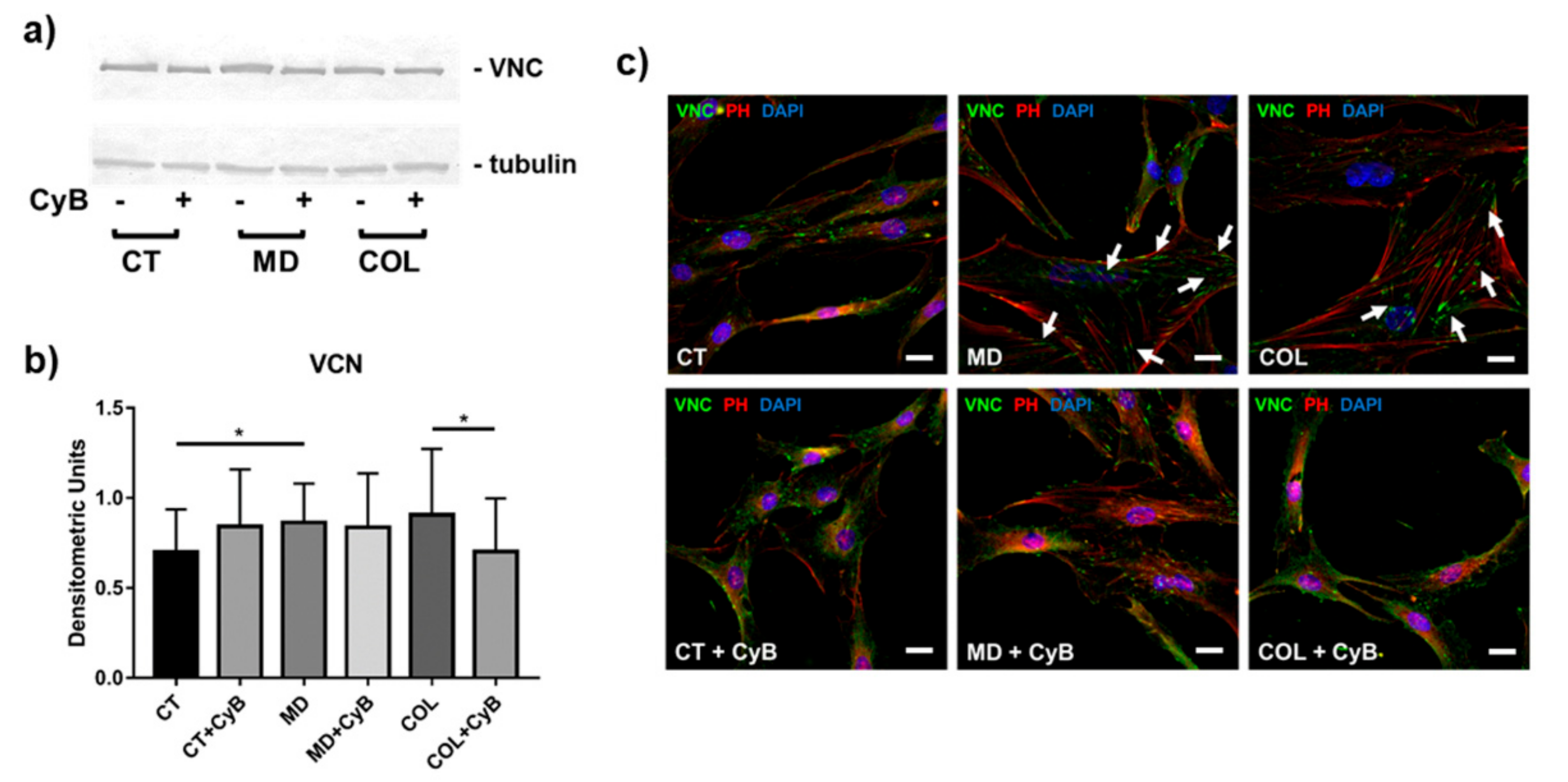
3.4. Cytoskeleton Arrangement and Vinculin Expression in Focal Adhesions
3.5. Wound Healing Assay
3.6. Expression of FAK, PAX and YAP/TAZ as Mechanosensors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Via, A.G.; Papa, G.; Oliva, F.; Maffulli, N. Tendinopathy. Curr. Phys. Med. Rehabil. Rep. 2016, 4, 50–55. [Google Scholar] [CrossRef]
- Steinmann, S.; Pfeifer, C.G.; Brochhausen, C.; Docheva, D. Spectrum of tendon pathologies: Triggers, trails and end-state. Int. J. Mol. Sci. 2020, 21, 844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, D. The management of greater trochanteric pain syndrome: A systematic literature review. J. Orthop. 2016, 13, 15–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frizziero, A.; Vittadini, F.; Pignataro, A.; Gasparre, G.; Biz, C.; Ruggieri, P.; Masiero, S. Conservative management of tendinopathies around hip. Muscles Ligaments Tendons J. 2016, 6, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.H.; Lee, J.Y.; Yoon, K.; Do, J.G.; Park, H.J.; Lee, S.Y.; Park, Y.S.; Lee, Y.T. Long-term outcome of low-energy extracorporeal shockwave therapy on gluteal tendinopathy documented by magnetic resonance imaging. PLoS ONE 2018, 13, e019746. [Google Scholar] [CrossRef] [PubMed]
- Patruno, M.; Martinello, T. Treatments of the injured tendon in Veterinary Medicine: From scaffolds to adult stem cells. Histol. Histopathol. 2014, 29, 417–422. [Google Scholar] [CrossRef]
- Haltmayer, E.; Ribitsch, I.; Gabner, S.; Rosser, J.; Gueltekin, S.; Peham, J.; Giese, U.; Dolezal, M.; Egerbacher, M.; Jenner, F. Co-culture of osteochondral explants and synovial membrane as in vitro model for osteoarthritis. PLoS ONE 2019, 14, e0214709. [Google Scholar] [CrossRef]
- O’Brien, C.; Marr, N.; Thorpe, C. Microdamage in the equine superficial digital flexor tendon. Equine Vet. J. 2020, 1–14. [Google Scholar] [CrossRef]
- Ferrario, C.; Rusconi, F.; Pulaj, A.; Macchi, R.; Landini, P.; Paroni, M.; Colombo, G.; Martinello, T.; Melotti, L.; Gomiero, C.; et al. from food waste to innovative biomaterial: Sea urchin-derived collagen for applications in skin regenerative medicine. Mar. Drugs 2020, 18, 414. [Google Scholar] [CrossRef]
- Martin, L.S.; Massafra, U.; Bizzi, E.; Migliore, A. A double blind randomized active-controlled clinical trial on the intra-articular use of Md-Knee versus sodium hyaluronate in patients with knee osteoarthritis (“Joint”). BMC Musculoskelet. Disord. 2016, 17, 94. [Google Scholar] [CrossRef] [Green Version]
- Pavelka, K.; Jarosova, H.; Sleglova, O.; Svobodova, R.; Votavova, M.; Milani, L.; Prochazka, Z.; Kotlarova, L.; Kostiuk, P.; Sliva, J.; et al. Chronic low back pain: Current pharmacotherapeutic therapies and a new biological approach. Curr. Med. Chem. 2018, 25, 1–8. [Google Scholar] [CrossRef]
- Kannus, P. Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef]
- Kjaer, M. Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol. Rev. 2004, 84, 649–698. [Google Scholar] [CrossRef]
- Riley, G.P.; Harrall, R.L.; Constant, C.R.; Chard, M.D.; Cawston, T.E.; Hazleman, B.L. Glycosaminoglycans of human rotator cuff tendons: Changes with age and in chronic rotator cuff tendinitis. Ann. Rheum. Dis. 1994, 53, 367–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banes, A.J.; Horesovsky, G.; Larson, C.; Tsuzaki, M.; Judex, S.; Archambault, J.; Zernicke, R.; Herzog, W.; Kelley, S.; Miller, L. Mechanical load stimulates expression of novel genes in vivo and in vitro in avian flexor tendon cells. Osteoarthr. Cartil. 1999, 7, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamin, M.; Ralphs, J.R. The cell and developmental biology of tendons and ligaments. Int. Rev. Cytol. 2000, 196, 85–130. [Google Scholar] [CrossRef]
- Jansen, K.A.; Donato, D.M.; Balcioglu, H.E.; Schmidt, T.; Danen, E.H.; Koenderink, G.H. A guide to mechanobiology: Where biology and physics meet. Biochim. Biophys. Acta 2015, 1853, 3043–3052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavagnino, M.; Wall, M.E.; Little, D.; Banes, A.J.; Guilak, F.; Arnoczky, S.P. Tendon mechanobiology: Current knowledge and future research opportunities. J. Orthop. Res. 2015, 33, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Randelli, F.; Menon, A.; Via, A.G.; Mazzoleni, M.G.; Sciancalepore, F.; Brioschi, M.; Gagliano, N. Effect of a collagen-based compound on morpho-functional properties of cultured human tenocytes. Cells 2018, 7, 246. [Google Scholar] [CrossRef] [Green Version]
- Canty, E.G.; Starborg, T.; Lu, Y.; Humphries, S.M.; Holmes, D.F.; Meadows, R.S.; Huffman, A.; O’Toole, E.T.; Kadler, K.E. Actin filaments are required for fibripositor-mediated collagen fibril alignment in tendon. J. Biol. Chem. 2006, 281, 38592–38598. [Google Scholar] [CrossRef] [Green Version]
- Carlomagno, C.; Banfi, P.I.; Gualerzi, A.; Volpato, E.; Meloni, M.; Lax, A.; Colombo, E.; Ticozzi, N.; Verde, F.; Silani, V.; et al. Human salivary Raman fingerprint as biomarker for the diagnosis of Amyotrophic Lateral Sclerosis. Sci. Rep. 2020, 10, 10175. [Google Scholar] [CrossRef] [PubMed]
- Menon, A.; Pettinari, L.; Martinelli, C.; Colombo, G.; Portinaro, N.; Dalle-Donne, I.; d’Agostino, M.C.; Gagliano, N. New insights in extracellular matrix remodeling and collagen turnover related pathways in cultured human tenocytes after ciprofloxacin administration. Muscle Ligaments Tendons J. 2013, 113, 122–131. [Google Scholar]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualerzi, A.; Niada, S.; Giannasi, C.; Picciolini, S.; Morasso, C.; Vanna, R.; Rossella, V.; Masserini, M.; Bedoni, M.; Ciceri, F.; et al. Raman spectroscopy uncovers biochemical tissue-related features of extracellular vesicles from mesenchymal stromal cells. Sci. Rep. 2017, 7, 9820. [Google Scholar] [CrossRef]
- Cullum, B.M.; Boca, S.; Rugina, D.; Pintea, A.; Leopold, N.; Astilean, S. Designing gold nanoparticle-ensembles as surface enhanced raman scattering tags inside human retinal cells. J. Nanotechnol. 2012, 961216. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.R.; Schultz, Z.D. SERS speciation of the electrochemical oxidation–reduction of riboflavin. Analyst 2016, 141, 5078–5087. [Google Scholar] [CrossRef] [Green Version]
- Cárcamo, J.J.; Aliaga, A.E.; Clavijo, R.E.; Brañes, M.R.; Campos-Vallette, M.M. Raman study of the shockwave effect on collagens. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2012, 86, 360–365. [Google Scholar] [CrossRef]
- Hannafin, J.A.; Arnoczky, S.P.; Hoonjan, A.; Torzilli, P.A. Effect of stress deprivation and cyclic tensile loading on the material and morphologic properties of canine flexor digitorum profundus tendon: An in vitro study. J. Orthop. Res. 1995, 13, 907–914. [Google Scholar] [CrossRef]
- Galloway, M.T.; Lalley, A.L.; Shearn, J.T. The role of mechanical loading in tendon development, maintenance, injury, and repair. J. Bone Jt. Surg. Am. 2013, 95, 1620–1628. [Google Scholar] [CrossRef]
- Magnusson, S.P.; Langberg, H.; Kjaer, M. The pathogenesis of tendinopathy: Balancing the response to loading. Nat. Rev. Rheumatol. 2010, 6, 262–268. [Google Scholar] [CrossRef]
- Heinemeier, K.M.; Kjaer, M. In vivo investigation of tendon responses to mechanical loading. J. Musculoskelet. Neuronal Interact. 2011, 11, 115–123. [Google Scholar] [PubMed]
- Banes, A.J.; Tsuzaki, M.; Yamamoto, J.; Brigman, B.; Fischer, T.; Brown, T.; Miller, L. Mechanoreception at the cellular level: The detection, interpretation and diversity of response to mechanical signals. Biochem. Cell Biol. 1995, 73, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Lavagnino, M.; Arnoczky, S.P.; Tian, T.; Vaupel, Z. Effect of amplitude and frequency of cyclic tensile strain on the inhibition of MMP-1 mRNA expression in tendon cells: An in vitro study. Connect. Tissue Res. 2003, 44, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Screen, H.R.; Shelton, J.C.; Bader, D.L.; Lee, D.A. Cyclic tensile strain upregulates collagen synthesis in isolated tendon fascicles. Biochem. Biophys. Res. Commun. 2005, 336, 424–429. [Google Scholar] [CrossRef]
- Langberg, H.; Rosendal, L.; Kjaer, M. Training-induced changes in peritendinous type I collagen turnover determined by microdialysis in humans. J. Physiol. 2001, 534, 297–302. [Google Scholar] [CrossRef]
- Arnoczky, S.P.; Tian, T.; Lavagnino, M.; Gardner, K. Ex vivo static tensile loading inhibits MMP-1 expression in rat tail tendon cells through a cytoskeletally based mechanotransduction mechanism. J. Orthop. Res. 2004, 22, 328–333. [Google Scholar] [CrossRef]
- Discher, D.E. Tissue cells feel and respon to the stiffness of their substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Martino, F.; Perestrelo, A.R.; Vinarský, V.; Pagliari, S.; Forte, G. Cellular mechanotransduction: From tension to function. Front. Physiol. 2018, 9, 824. [Google Scholar] [CrossRef]
- Silver, F.H.; Christiansen, D.; Snowhill, P.B.; Chen, Y.; Landis, W.J. The role of mineral in the storage of elastic energy in turkey tendons. Biomacromolecules 2000, 1, 180–185. [Google Scholar] [CrossRef]
- Walker, L.C.; Overstreet, M.A.; Yeowell, H.N. Tissue-specific expression and regulation of the alternatively-spliced forms of lysyl hydroxylase 2 (LH2) in human kidney cells and skin fibroblasts. Matrix. Biol. 2005, 23, 515–523. [Google Scholar] [CrossRef]
- Sakai, T.; Gross, J. Some properties of the products of reaction of tadpole collagenase with collagen. Biochemistry 1967, 6, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Woessner, F.J. Matrix metalloproteinases and their inhibitors in connective tissue remodelling. FASEB J. 1991, 5, 2145–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brew, K.; Dinakarpandian, D.; Nagase, H. Tissue inhibitors of metalloproteinases: Evolution, structure and function. Biochim. Biophys. Acta 2001, 1477, 267–283. [Google Scholar] [CrossRef]
- Murphy, G.; Willenbrock, F.; Crabbe, T.; O’Shea, M.; Ward, R.; Atkinson, S.; O’Connell, J.; Docherty, A. Regulation of matrix metalloproteinase activity. Ann. N. Y. Acad. Sci. 1994, 732, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Burridge, K.; Guilluy, C. Focal adhesions, stress fibers and mechanical tension. Exp. Cell Res. 2016, 343, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.H.; Guo, Q.; Li, B. Tendon biomechanics and mechanobiology-a minireview of basic concepts and recent advancements. J. Hand Ther. 2012, 25, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.E.; Mudera, V.; Brown, R.A.; Cambrey, A.D.; Grobbelaar, A.O.; McGrouther, D.A. The early surface cell response to flexor tendon injury. J. Hand Surg. Am. 2003, 28, 221–230. [Google Scholar] [CrossRef]
- Reed, M.J.; Ferara, N.S.; Vernon, R.B. Impaired migration, integrin function, and actin cytoskeletal organization in dermal fibroblasts from a subset of aged human donors. Mech. Ageing Dev. 2001, 122, 1203–1220. [Google Scholar] [CrossRef]
- Pesqueira, T.; Costa-Almeida, R.; Gomes, M.E. Uncovering the effect of low-frequency static magnetic field on tendon-derived cells: From mechanosensing to tenogenesis. Sci. Rep. 2017, 7, 10948. [Google Scholar] [CrossRef] [Green Version]
- Spanjaard, E.; de Rooij, J. Mechanotransduction: Vinculin provides stability when tension rises. Curr. Biol. 2013, 23, R159–R161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totaro, A.; Panciera, T.; Piccolo, S. YAP/TAZ upstream signals and downstream responses. Nat. Cell Biol. 2018, 20, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Low, B.C.; Qiurong Pan, C.; Shivashankar, G.V.; Bershadsky, A.; Sudol, M.; Sheetz, M. YAP/TAZ as mechanosensors and mechanotransducers in regulating organ size and tumor growth. FEBS Lett. 2014, 588, 2663–2670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Halder, G.; Dupont, S.; Piccolo, S. Transduction of mechanical and cytoskeletal cues by YAP and TAZ. Nat. Rev. Mol. Cell Biol. 2012, 13, 591–600. [Google Scholar] [CrossRef]
- Schroeder, M.C.; Halder, G. Regulation of the Hippo pathway by cell architecture and mechanical signals. Semin. Cell Dev. Biol. 2012, 23, 803–811. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raman Shift (cm−1) | Attribution |
|---|---|
| 475 | Skeletal deformations |
| 508 | Skeletal deformations |
| 800 | Skeletal C-C vibrations |
| 920 | C-COO− vibrations |
| 992 | Phenylalanine |
| 1015 | Vibration of Proline C-N |
| 1055 | Distortion of Proline N-C-H |
| 1235 | Amide III |
| 1403 | Deformation of CH3 |
| 1443 | Deformation of CH3 |
| 1544 | Deformation of NH3+ |
| 1665 | Amide I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Randelli, F.; Sartori, P.; Carlomagno, C.; Bedoni, M.; Menon, A.; Vezzoli, E.; Sommariva, M.; Gagliano, N. The Collagen-Based Medical Device MD-Tissue Acts as a Mechanical Scaffold Influencing Morpho-Functional Properties of Cultured Human Tenocytes. Cells 2020, 9, 2641. https://doi.org/10.3390/cells9122641
Randelli F, Sartori P, Carlomagno C, Bedoni M, Menon A, Vezzoli E, Sommariva M, Gagliano N. The Collagen-Based Medical Device MD-Tissue Acts as a Mechanical Scaffold Influencing Morpho-Functional Properties of Cultured Human Tenocytes. Cells. 2020; 9(12):2641. https://doi.org/10.3390/cells9122641
Chicago/Turabian StyleRandelli, Filippo, Patrizia Sartori, Cristiano Carlomagno, Marzia Bedoni, Alessandra Menon, Elena Vezzoli, Michele Sommariva, and Nicoletta Gagliano. 2020. "The Collagen-Based Medical Device MD-Tissue Acts as a Mechanical Scaffold Influencing Morpho-Functional Properties of Cultured Human Tenocytes" Cells 9, no. 12: 2641. https://doi.org/10.3390/cells9122641
APA StyleRandelli, F., Sartori, P., Carlomagno, C., Bedoni, M., Menon, A., Vezzoli, E., Sommariva, M., & Gagliano, N. (2020). The Collagen-Based Medical Device MD-Tissue Acts as a Mechanical Scaffold Influencing Morpho-Functional Properties of Cultured Human Tenocytes. Cells, 9(12), 2641. https://doi.org/10.3390/cells9122641