Regulation of Synaptic Development by Astrocyte Signaling Factors and Their Emerging Roles in Substance Abuse

{kind=link}
Abstract
:1. Introduction
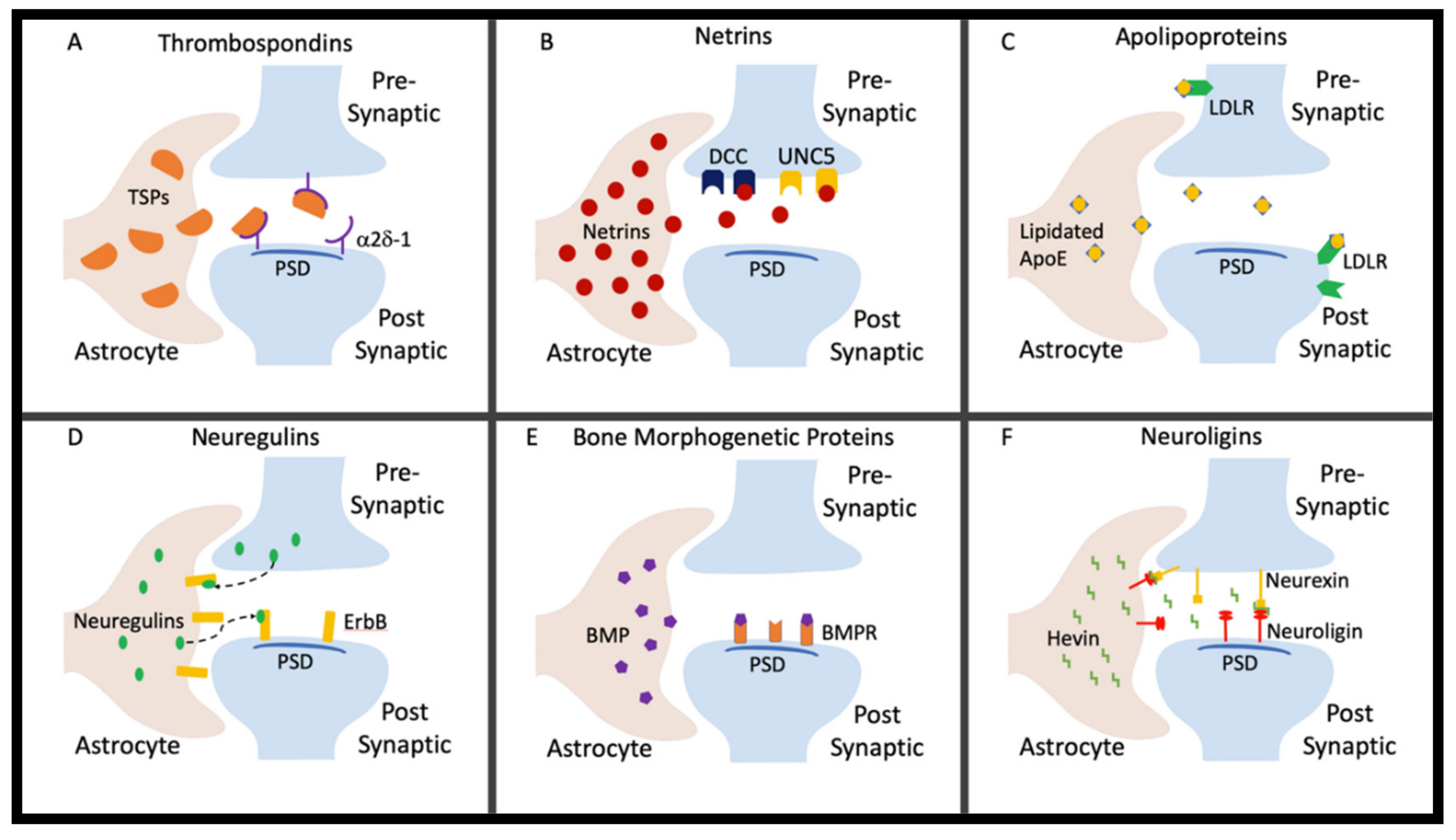
2. Thrombospondins
3. Netrins
4. Apolipoproteins
5. Neuregulins
6. Bone Morphogenetic Proteins
7. Neuroligins
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kubotera, H.; Ikeshima-Kataoka, H.; Hatashita, Y.; Allegra Mascaro, A.L.; Pavone, F.S.; Inoue, T. Astrocytic endfeet re-cover blood vessels after removal by laser ablation. Sci. Rep. 2019, 9, 1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, R.; Harris, K.M. Three-dimensional relationships between hippocampal synapses and astrocytes. J. Neurosci. 1999, 19, 6897–6906. [Google Scholar] [CrossRef] [PubMed]
- Bushong, E.A.; Martone, M.E.; Jones, Y.Z.; Ellisman, M.H. Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains. J. Neurosci. 2002, 22, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Oberheim, N.A.; Takano, T.; Han, X.; He, W.; Lin, J.H.; Wang, F.; Xu, Q.; Wyatt, J.D.; Pilcher, W.; Ojemann, J.G.; et al. Uniquely hominid features of adult human astrocytes. J. Neurosci. 2009, 29, 3276–3287. [Google Scholar] [CrossRef]
- Ullian, E.M.; Sapperstein, S.K.; Christopherson, K.S.; Barres, B.A. Control of synapse number by glia. Science 2001, 291, 657–661. [Google Scholar] [CrossRef]
- Christopherson, K.S.; Ullian, E.M.; Stokes, C.C.; Mullowney, C.E.; Hell, J.W.; Agah, A.; Lawler, J.; Mosher, D.F.; Bornstein, P.; Barres, B.A. Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell 2005, 120, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Farhy-Tselnicker, I.; Allen, N.J. Astrocytes, neurons, synapses: A tripartite view on cortical circuit development. Neural Dev. 2018, 13, 7. [Google Scholar] [CrossRef] [Green Version]
- Wiese, S.; Karus, M.; Faissner, A. Astrocytes as a source for extracellular matrix molecules and cytokines. Front. Pharmacol. 2012, 3, 120. [Google Scholar] [CrossRef] [Green Version]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic analysis of reactive astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.; Dorovini-Zis, K.; Vincent, S.R. Cytokines, nitric oxide, and cGMP modulate the permeability of an in vitro model of the human blood-brain barrier. Exp. Neurol. 2004, 190, 446–455. [Google Scholar] [CrossRef]
- Fan, Y.; Shen, F.; Chen, Y.; Hao, Q.; Liu, W.; Su, H.; Young, W.L.; Yang, G.Y. Overexpression of netrin-1 induces neovascularization in the adult mouse brain. J. Cereb. Blood Flow Metab. 2008, 28, 1543–1551. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Stevens, C.F.; Gage, F.H. Astroglia induce neurogenesis from adult neural stem cells. Nature 2002, 417, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Dickson, B.J. Molecular mechanisms of axon guidance. Science 2002, 298, 1959–1964. [Google Scholar] [CrossRef] [Green Version]
- Khakh, B.S.; Sofroniew, M.V. Diversity of astrocyte functions and phenotypes in neural circuits. Nat. Neurosci. 2015, 18, 942–952. [Google Scholar] [CrossRef]
- Matias, I.; Morgado, J.; Gomes, F.C.A. Astrocyte Heterogeneity: Impact to Brain Aging and Disease. Front. Aging Neurosci. 2019, 11, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, L.E.; Liddelow, S.A.; Chakraborty, C.; Munch, A.E.; Heiman, M.; Barres, B.A. Normal aging induces A1-like astrocyte reactivity. Proc. Natl. Acad. Sci. USA 2018, 115, E1896–E1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karve, I.P.; Taylor, J.M.; Crack, P.J. The contribution of astrocytes and microglia to traumatic brain injury. Br. J. Pharmacol 2016, 173, 692–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Pekny, M.; Wilhelmsson, U.; Pekna, M. The dual role of astrocyte activation and reactive gliosis. Neurosci. Lett. 2014, 565, 30–38. [Google Scholar] [CrossRef]
- Zakharova, M.; Ziegler, H.K. Paradoxical anti-inflammatory actions of TNF-alpha: Inhibition of IL-12 and IL-23 via TNF receptor 1 in macrophages and dendritic cells. J. Immunol. 2005, 175, 5024–5033. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.T.; Boeshore, K.L.; Wu, C.; Becker, K.G.; Errico, S.L.; Mash, D.C.; Freed, W.J. Cocaine promotes primary human astrocyte proliferation via JNK-dependent up-regulation of cyclin A2. Restor. Neurol. Neurosci. 2016, 34, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Lasic, E.; Lisjak, M.; Horvat, A.; Bozic, M.; Sakanovic, A.; Anderluh, G.; Verkhratsky, A.; Vardjan, N.; Jorgacevski, J.; Stenovec, M.; et al. Astrocyte Specific Remodeling of Plasmalemmal Cholesterol Composition by Ketamine Indicates a New Mechanism of Antidepressant Action. Sci. Rep. 2019, 9, 10957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stansfield, W.E.; Ranek, M.; Pendse, A.; Schisler, J.C.; Wang, S.; Pulinilkunnil, T.; Willis, M.S. The Pathophysiology of Cardiac Hypertrophy and Heart Failure. In Cellular and Molecular Pathobiology of Cardiovascular Disease; Willis, M.S., Homeister, J.W., Stone, J.R., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2014; pp. 51–78. [Google Scholar]
- Risher, W.C.; Eroglu, C. Thrombospondins as key regulators of synaptogenesis in the central nervous system. Matrix Biol. 2012, 31, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eroglu, C.; Allen, N.J.; Susman, M.W.; O’Rourke, N.A.; Park, C.Y.; Ozkan, E.; Chakraborty, C.; Mulinyawe, S.B.; Annis, D.S.; Huberman, A.D.; et al. Gabapentin receptor alpha2delta-1 is a neuronal thrombospondin receptor responsible for excitatory CNS synaptogenesis. Cell 2009, 139, 380–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risher, W.C.; Kim, N.; Koh, S.; Choi, J.E.; Mitev, P.; Spence, E.F.; Pilaz, L.J.; Wang, D.; Feng, G.; Silver, D.L.; et al. Thrombospondin receptor alpha2delta-1 promotes synaptogenesis and spinogenesis via postsynaptic Rac1. J. Cell Biol. 2018, 217, 3747–3765. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Kipnis, J. Thrombospondin 1--a key astrocyte-derived neurogenic factor. FASEB J. 2010, 24, 1925–1934. [Google Scholar] [CrossRef] [Green Version]
- Spencer, S.; Brown, R.M.; Quintero, G.C.; Kupchik, Y.M.; Thomas, C.A.; Reissner, K.J.; Kalivas, P.W. alpha2delta-1 signaling in nucleus accumbens is necessary for cocaine-induced relapse. J. Neurosci. 2014, 34, 8605–8611. [Google Scholar] [CrossRef] [Green Version]
- Risher, M.L.; Sexton, H.G.; Risher, W.C.; Wilson, W.A.; Fleming, R.L.; Madison, R.D.; Moore, S.D.; Eroglu, C.; Swartzwelder, H.S. Adolescent Intermittent Alcohol Exposure: Dysregulation of Thrombospondins and Synapse Formation are Associated with Decreased Neuronal Density in the Adult Hippocampus. Alcohol Clin. Exp. Res. 2015, 39, 2403–2413. [Google Scholar] [CrossRef]
- Hakanen, J.; Duprat, S.; Salminen, M. Netrin1 is required for neural and glial precursor migrations into the olfactory bulb. Dev. Biol. 2011, 355, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Yung, A.R.; Nishitani, A.M.; Goodrich, L.V. Phenotypic analysis of mice completely lacking netrin 1. Development 2015, 142, 3686–3691. [Google Scholar] [CrossRef] [Green Version]
- Lai Wing Sun, K.; Correia, J.P.; Kennedy, T.E. Netrins: Versatile extracellular cues with diverse functions. Development 2011, 138, 2153–2169. [Google Scholar] [PubMed] [Green Version]
- Harter, P.N.; Bunz, B.; Dietz, K.; Hoffmann, K.; Meyermann, R.; Mittelbronn, M. Spatio-temporal deleted in colorectal cancer (DCC) and netrin-1 expression in human foetal brain development. Neuropathol. Appl. Neurobiol. 2010, 36, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Chen, R.; Chen, H.; Zhang, Y.; Chen, J.; Lin, P.; Lan, Q.; Yuan, Q.; Lai, Y.; Jiang, X.; et al. Netrin-1 Promotes Synaptic Formation and Axonal Regeneration via JNK1/c-Jun Pathway after the Middle Cerebral Artery Occlusion. Front. Cell Neurosci. 2018, 12, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, J.S.; Ashour, M.A.; Magdesian, M.H.; Tritsch, N.X.; Harris, S.N.; Christofi, N.; Chemali, R.; Stern, Y.E.; Thompson-Steckel, G.; Gris, P.; et al. Netrin-1 promotes excitatory synaptogenesis between cortical neurons by initiating synapse assembly. J. Neurosci. 2013, 33, 17278–17289. [Google Scholar] [CrossRef]
- Matsukawa, H.; Akiyoshi-Nishimura, S.; Zhang, Q.; Lujan, R.; Yamaguchi, K.; Goto, H.; Yaguchi, K.; Hashikawa, T.; Sano, C.; Shigemoto, R.; et al. Netrin-G/NGL complexes encode functional synaptic diversification. J. Neurosci. 2014, 34, 15779–15792. [Google Scholar] [CrossRef] [PubMed]
- Kelai, S.; Ramoz, N.; Moalic, J.M.; Noble, F.; Mechawar, N.; Imbeaud, S.; Turecki, G.; Simonneau, M.; Gorwood, P.; Maussion, G. Netrin G1: Its downregulation in the nucleus accumbens of cocaine-conditioned mice and genetic association in human cocaine dependence. Addict. Biol. 2018, 23, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Serafini, T.; Colamarino, S.A.; Leonardo, E.D.; Wang, H.; Beddington, R.; Skarnes, W.C.; Tessier-Lavigne, M. Netrin-1 is required for commissural axon guidance in the developing vertebrate nervous system. Cell 1996, 87, 1001–1014. [Google Scholar] [CrossRef] [Green Version]
- Elliott, D.A.; Weickert, C.S.; Garner, B. Apolipoproteins in the brain: Implications for neurological and psychiatric disorders. Clin. Lipidol. 2010, 51, 555–573. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, Q. Cholesterol metabolism and homeostasis in the brain. Protein Cell 2015, 6, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Liu, C.C.; Kanekiyo, T.; Xu, H.; Bu, G. Apolipoprotein E and Alzheimer disease: Risk, mechanisms and therapy. Nat. Rev. Neurol. 2013, 9, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Harris, F.M.; Tesseur, I.; Brecht, W.J.; Xu, Q.; Mullendorff, K.; Chang, S.; Wyss-Coray, T.; Mahley, R.W.; Huang, Y. Astroglial regulation of apolipoprotein E expression in neuronal cells. Implications for Alzheimer’s disease. J. Biol. Chem. 2004, 279, 3862–3868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spagnuolo, M.S.; Donizetti, A.; Iannotta, L.; Aliperti, V.; Cupidi, C.; Bruni, A.C.; Cigliano, L. Brain-derived neurotrophic factor modulates cholesterol homeostasis and Apolipoprotein E synthesis in human cell models of astrocytes and neurons. J. Cell Physiol. 2018, 233, 6925–6943. [Google Scholar] [CrossRef] [PubMed]
- Koistinaho, M.; Lin, S.; Wu, X.; Esterman, M.; Koger, D.; Hanson, J.; Higgs, R.; Liu, F.; Malkani, S.; Bales, K.R.; et al. Apolipoprotein E promotes astrocyte colocalization and degradation of deposited amyloid-beta peptides. Nat. Med. 2004, 10, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Cartocci, V.; Servadio, M.; Trezza, V.; Pallottini, V. Can Cholesterol Metabolism Modulation Affect Brain Function and Behavior? J. Cell Physiol. 2017, 232, 281–286. [Google Scholar] [CrossRef]
- Fernandez, C.G.; Hamby, M.E.; McReynolds, M.L.; Ray, W.J. The Role of APOE4 in Disrupting the Homeostatic Functions of Astrocytes and Microglia in Aging and Alzheimer’s Disease. Front. Aging Neurosci. 2019, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Farrer, L.A.; Cupples, L.A.; Haines, J.L.; Hyman, B.; Kukull, W.A.; Mayeux, R.; Myers, R.H.; Pericak-Vance, M.A.; Risch, N.; van Duijn, C.M. Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. APOE and Alzheimer Disease Meta Analysis Consortium. JAMA 1997, 278, 1349–1356. [Google Scholar] [CrossRef]
- Li, L.; Liu, M.S.; Li, G.Q.; Tang, J.; Liao, Y.; Zheng, Y.; Guo, T.L.; Kang, X.; Yuan, M.T. Relationship between Apolipoprotein Superfamily and Parkinson’s Disease. Chin. Med. J. (Engl) 2017, 130, 2616–2623. [Google Scholar] [CrossRef]
- Lewohl, J.M.; Wang, L.; Miles, M.F.; Zhang, L.; Dodd, P.R.; Harris, R.A. Gene expression in human alcoholism: Microarray analysis of frontal cortex. Alcohol Clin. Exp. Res. 2000, 24, 1873–1882. [Google Scholar] [CrossRef]
- Bechtholt, A.J.; Smith, R.; Raber, J.; Cunningham, C.L. Enhanced ethanol-, but not cocaine-induced, conditioned place preference in Apoe(-/-) mice. Pharmacol. Biochem. Behav. 2004, 77, 783–792. [Google Scholar] [CrossRef]
- Djousse, L.; Himali, J.J.; Beiser, A.; Kelly-Hayes, M.; Wolf, P.A. Apolipoprotein e, alcohol consumption, and risk of ischemic stroke: The Framingham Heart Study revisited. J. Stroke Cerebrovasc. Dis. 2009, 18, 384–388. [Google Scholar] [CrossRef] [Green Version]
- Harwood, D.G.; Kalechstein, A.; Barker, W.W.; Strauman, S.; St George-Hyslop, P.; Iglesias, C.; Loewenstein, D.; Duara, R. The effect of alcohol and tobacco consumption, and apolipoprotein E genotype, on the age of onset in Alzheimer’s disease. Int. J. Geriatr. Psychiatry 2010, 25, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Falls, D.L. Neuregulins and the neuromuscular system: 10 years of answers and questions. J. Neurocytol. 2003, 32, 619–647. [Google Scholar] [CrossRef]
- Yang, X.; Kuo, Y.; Devay, P.; Yu, C.; Role, L. A cysteine-rich isoform of neuregulin controls the level of expression of neuronal nicotinic receptor channels during synaptogenesis. Neuron 1998, 20, 255–270. [Google Scholar] [CrossRef] [Green Version]
- Talmage, D.A. Mechanisms of neuregulin action. Novartis Found. Symp. 2008, 289, 74–84. [Google Scholar] [PubMed]
- Deakin, I.H.; Godlewska, B.R.; Walker, M.A.; Huang, G.J.; Schwab, M.H.; Nave, K.A.; Law, A.J.; Harrison, P.J. Altered hippocampal gene expression and structure in transgenic mice overexpressing neuregulin 1 (Nrg1) type I. Transl. Psychiatry 2018, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Deakin, I.H.; Nissen, W.; Law, A.J.; Lane, T.; Kanso, R.; Schwab, M.H.; Nave, K.A.; Lamsa, K.P.; Paulsen, O.; Bannerman, D.M.; et al. Transgenic overexpression of the type I isoform of neuregulin 1 affects working memory and hippocampal oscillations but not long-term potentiation. Cereb. Cortex 2012, 22, 1520–1529. [Google Scholar] [CrossRef] [Green Version]
- Vaht, M.; Laas, K.; Kiive, E.; Parik, J.; Veidebaum, T.; Harro, J. A functional neuregulin-1 gene variant and stressful life events: Effect on drug use in a longitudinal population-representative cohort study. J. Psychopharmacol 2017, 31, 54–61. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef]
- Jha, M.K.; Kim, J.H.; Song, G.J.; Lee, W.H.; Lee, I.K.; Lee, H.W.; An, S.S.A.; Kim, S.; Suk, K. Functional dissection of astrocyte-secreted proteins: Implications in brain health and diseases. Prog. Neurobiol. 2018, 162, 37–69. [Google Scholar] [CrossRef]
- Mabie, P.C.; Mehler, M.F.; Marmur, R.; Papavasiliou, A.; Song, Q.; Kessler, J.A. Bone morphogenetic proteins induce astroglial differentiation of oligodendroglial-astroglial progenitor cells. J. Neurosci. 1997, 17, 4112–4120. [Google Scholar] [CrossRef]
- Cole, A.E.; Murray, S.S.; Xiao, J. Bone Morphogenetic Protein 4 Signalling in Neural Stem and Progenitor Cells during Development and after Injury. Stem Cells Int. 2016, 2016, 9260592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykaras, N.; Opperman, L.A. Bone morphogenetic proteins (BMPs): How do they function and what can they offer the clinician? J. Oral Sci. 2003, 45, 57–73. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, J.S.; Yetz-Aldape, J.; Wang, E.A. Bone morphogenetic proteins induce differentiation in astrocyte lineage cells. Growth Factors 1994, 11, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Wilkemeyer, M.F.; Pajerski, M.; Charness, M.E. Alcohol inhibition of cell adhesion in BMP-treated NG108-15 cells. Alcohol Clin. Exp. Res. 1999, 23, 1711–1720. [Google Scholar] [CrossRef]
- Skowronska, K.; Obara-Michlewska, M.; Zielinska, M.; Albrecht, J. NMDA Receptors in Astrocytes: In Search for Roles in Neurotransmission and Astrocytic Homeostasis. Int. J. Mol. Sci. 2019, 20, 309. [Google Scholar] [CrossRef] [Green Version]
- Dorit, R.; Jun, W. The NMDA Receptor and Alcohol Addiction. In Biology of the NMDA Receptor; Van Dongen, A.M., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2009. [Google Scholar]
- Newcomer, J.W.; Farber, N.B.; Olney, J.W. NMDA receptor function, memory, and brain aging. Dialogues Clin. Neurosci. 2000, 2, 219–232. [Google Scholar]
- Dalvi, P.; O’Brien-Ladner, A.; Dhillon, N.K. Downregulation of bone morphogenetic protein receptor axis during HIV-1 and cocaine-mediated pulmonary smooth muscle hyperplasia: Implications for HIV-related pulmonary arterial hypertension. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2585–2595. [Google Scholar] [CrossRef] [Green Version]
- Varoqueaux, F.; Jamain, S.; Brose, N. Neuroligin 2 is exclusively localized to inhibitory synapses. Eur. J. Cell Biol. 2004, 83, 449–456. [Google Scholar] [CrossRef]
- Stogsdill, J.A.; Ramirez, J.; Liu, D.; Kim, Y.H.; Baldwin, K.T.; Enustun, E.; Ejikeme, T.; Ji, R.-R.; Eroglu, C. Astrocytic neuroligins control astrocyte morphogenesis and synaptogenesis. Nature 2017, 551, 192–197. [Google Scholar] [CrossRef]
- Song, J.Y.; Ichtchenko, K.; Sudhof, T.C.; Brose, N. Neuroligin 1 is a postsynaptic cell-adhesion molecule of excitatory synapses. Proc. Natl. Acad. Sci. USA 1999, 96, 1100–1105. [Google Scholar] [CrossRef] [Green Version]
- Budreck, E.C.; Scheiffele, P. Neuroligin-3 is a neuronal adhesion protein at GABAergic and glutamatergic synapses. Eur. J. Neurosci. 2007, 26, 1738–1748. [Google Scholar] [CrossRef] [PubMed]
- Octeau, J.C.; Chai, H.; Jiang, R.; Bonanno, S.L.; Martin, K.C.; Khakh, B.S. An Optical Neuron-Astrocyte Proximity Assay at Synaptic Distance Scales. Neuron 2018, 98, 49–66.e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.K.; Stogsdill, J.A.; Pulimood, N.S.; Dingsdale, H.; Kim, Y.H.; Pilaz, L.J.; Kim, I.H.; Manhaes, A.C.; Rodrigues, W.S.; Pamukcu, A.; et al. Astrocytes Assemble Thalamocortical Synapses by Bridging NRX1alpha and NL1 via Hevin. Cell 2016, 164, 183–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhof, T.C. Neuroligins and neurexins link synaptic function to cognitive disease. Nature 2008, 455, 903–911. [Google Scholar] [CrossRef] [Green Version]
- Hishimoto, A.; Liu, Q.R.; Drgon, T.; Pletnikova, O.; Walther, D.; Zhu, X.G.; Troncoso, J.C.; Uhl, G.R. Neurexin 3 polymorphisms are associated with alcohol dependence and altered expression of specific isoforms. Hum. Mol. Genet. 2007, 16, 2880–2891. [Google Scholar] [CrossRef]
- Herman, M.A.; Jahr, C.E. Extracellular glutamate concentration in hippocampal slice. J. Neurosci. 2007, 27, 9736–9741. [Google Scholar] [CrossRef]
- Jackson, J.G.; O’Donnell, J.C.; Takano, H.; Coulter, D.A.; Robinson, M.B. Neuronal activity and glutamate uptake decrease mitochondrial mobility in astrocytes and position mitochondria near glutamate transporters. J. Neurosci. 2014, 34, 1613–1624. [Google Scholar] [CrossRef] [Green Version]
- Bolewska, P.; Martin, B.I.; Orlando, K.A.; Rhoads, D.E. Sequential Changes in Brain Glutamate and Adenosine A1 Receptors May Explain Severity of Adolescent Alcohol Withdrawal after Consumption of High Levels of Alcohol. Neurosci. J. 2019, 2019, 5950818. [Google Scholar] [CrossRef] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, C.D.; Risher, W.C.; Risher, M.-L. Regulation of Synaptic Development by Astrocyte Signaling Factors and Their Emerging Roles in Substance Abuse. Cells 2020, 9, 297. https://doi.org/10.3390/cells9020297
Walker CD, Risher WC, Risher M-L. Regulation of Synaptic Development by Astrocyte Signaling Factors and Their Emerging Roles in Substance Abuse. Cells. 2020; 9(2):297. https://doi.org/10.3390/cells9020297
Chicago/Turabian StyleWalker, Christopher D., W. Christopher Risher, and Mary-Louise Risher. 2020. "Regulation of Synaptic Development by Astrocyte Signaling Factors and Their Emerging Roles in Substance Abuse" Cells 9, no. 2: 297. https://doi.org/10.3390/cells9020297
APA StyleWalker, C. D., Risher, W. C., & Risher, M. -L. (2020). Regulation of Synaptic Development by Astrocyte Signaling Factors and Their Emerging Roles in Substance Abuse. Cells, 9(2), 297. https://doi.org/10.3390/cells9020297