Analysis of Killer Immunoglobulin-Like Receptor Genes in Colorectal Cancer

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Genotyping and Imputation
2.3. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet (Lond. Engl.) 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Schmit, S.L.; Edlund, C.K.; Schumacher, F.R.; Gong, J.; Harrison, T.A.; Huyghe, J.R.; Qu, C.; Melas, M.; Van Den Berg, D.J.; Wang, H.; et al. Novel Common Genetic Susceptibility Loci for Colorectal Cancer. J. Natl. Cancer Inst. 2019, 111, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Law, P.J.; Timofeeva, M.; Fernandez-Rozadilla, C.; Broderick, P.; Studd, J.; Fernandez-Tajes, J.; Farrington, S.; Svinti, V.; Palles, C.; Orlando, G.; et al. Association analyses identify 31 new risk loci for colorectal cancer susceptibility. Nat. Commun. 2019, 10, 2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillerey, C.; Huntington, N.D.; Smyth, M.J. Targeting natural killer cells in cancer immunotherapy. Nat. Immunol. 2016, 17, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Parham, P.; Guethlein, L.A. Genetics of Natural Killer Cells in Human Health, Disease, and Survival. Annu. Rev. Immunol. 2018, 36. [Google Scholar] [CrossRef]
- Lanier, L.L.; Cortiss, B.C.; Wu, J.; Leong, C.; Phillips, J.H. Immunoreceptor DAP12 bearing a tyrosine-based activation motif is involved in activating NK cells. Nature 1998, 391, 703–707. [Google Scholar] [CrossRef]
- Olcese, L.; Cambiaggi, A.; Semenzato, G.; Bottino, C.; Moretta, A.; Vivier, E. Human killer cell activatory receptors for MHC class I molecules are included in a multimeric complex expressed by natural killer cells. J. Immunol. 1997, 158, 5083–5086. [Google Scholar]
- Shahjahan Miah, S.M.; Hughes, T.L.; Campbell, K.S. KIR2DL4 Differentially Signals Downstream Functions in Human NK Cells through Distinct Structural Modules. J. Immunol. 2008, 180, 2922–2932. [Google Scholar] [CrossRef] [Green Version]
- Winter, C.C.; Gumperz, J.E.; Parham, P.; Long, E.O.; Wagtmann, N. Direct binding and functional transfer of NK cell inhibitory receptors reveal novel patterns of HLA-C allotype recognition. J. Immunol. 1998, 161, 571–577. [Google Scholar]
- Cella, M.; Longo, A.; Ferrara, G.B.; Strominger, J.L.; Colonna, M. NK3-specific natural killer cells are selectively inhibited by Bw4-positive HLA alleles with isoleucine 80. J. Exp. Med. 1994, 180, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Hansasuta, P.; Dong, T.; Thananchai, H.; Weekes, M.; Willberg, C.; Aldemir, H.; Rowland-Jones, S.; Braud, V.M. Recognition of HLA-A3 and HLA-A11 by KIR3DL2 is peptide-specific. Eur. J. Immunol. 2004. [Google Scholar] [CrossRef] [PubMed]
- Augusto, D.G. The impact of KIR polymorphism on the risk of developing cancer: Not as strong as imagined? Front. Genet. 2016, 7, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, A.G.; Morreau, H.; Tollenaar, R.A.E.M.; Alphenaar, E.; Van Puijenbroek, M.; Putter, H.; Janssen-Van Rhijn, C.M.; Van De Velde, C.J.H.; Fleuren, G.J.; Kuppen, P.J.K. Down-regulation of HLA-A expression correlates with a better prognosis in colorectal cancer patients. Lab. Investig. A J. Tech. Methods Pathol. 2002, 82, 1725–1733. [Google Scholar] [CrossRef] [Green Version]
- Watson, N.F.S.; Ramage, J.M.; Madjd, Z.; Spendlove, I.; Ellis, I.O.; Scholefield, J.H.; Durrant, L.G. Immunosurveillance is active in colorectal cancer as downregulation but not complete loss of MHC class I expression correlates with a poor prognosis. Int. J. Cancer 2006, 118, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Shen, J.; Cox, C.; Wakefield, J.C.; Ehm, M.G.; Nelson, M.R.; Weir, B.S. HIBAG—HLA genotype imputation with attribute bagging. Pharm. J. 2014, 14, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, J.; Gurdasani, D.; Delaneau, O.; Pirastu, N.; Ulivi, S.; Cocca, M.; Traglia, M.; Huang, J.; Huffman, J.E.; Rudan, I.; et al. A general approach for haplotype phasing across the full spectrum of relatedness. PLoS Genet. 2014, 10, e1004234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vukcevic, D.; Traherne, J.A.; Næss, S.; Ellinghaus, E.; Kamatani, Y.; Dilthey, A.; Lathrop, M.; Karlsen, T.H.; Franke, A.; Moffatt, M.; et al. Imputation of KIR Types from SNP Variation Data. Am. J. Hum. Genet. 2015, 97, 593–607. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Song, Y.-Q. PyHLA: Tests for the association between HLA alleles and diseases. BMC Bioinform. 2017, 18, 90. [Google Scholar] [CrossRef] [Green Version]
- Middleton, D.; Gonzelez, F. The extensive polymorphism of KIR genes. Immunology 2010, 129, 8–19. [Google Scholar] [CrossRef]
- Middleton, D.; Vilchez, J.R.; Cabrera, T.; Meenagh, A.; Williams, F.; Halfpenny, I.; Maleno, I.; Ruiz-Cabello, F.; Lopez-Nevot, M.A.; Garrido, F. Analysis of KIR gene frequencies in HLA class I characterised bladder, colorectal and laryngeal tumours. Tissue Antigens 2007, 69, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Al Omar, S.; Middleton, D.; Marshall, E.; Porter, D.; Xinarianos, G.; Raji, O.; Field, J.K.; Christmas, S.E. Associations between genes for killer immunoglobulin-like receptors and their ligands in patients with solid tumors. Hum. Immunol. 2010, 71, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Choi, H.B.; Jang, J.P.; Baek, I.C.; Choi, E.J.; Park, M.; Kim, T.G.; Oh, S.T. HLA-Cw polypmorphism and killer cell immunoglobulin-like receptor (KIR) gene analysis in Korean colorectal cancer patients. Int. J. Surg. 2014, 12, 815–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Omar, S.Y.; Mansour, L.; Dar, J.A.; Alwasel, S.; Alkhuriji, A.; Arafah, M.; Al Obeed, O.; Christmas, S. The Relationship between Killer Cell Immunoglobulin-Like Receptors and HLA-C Polymorphisms in Colorectal Cancer in a Saudi Population. Genet. Test. Mol. Biomark. 2015, 19, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Ghanadi, K.; Shayanrad, B.; Ahmadi, S.A.Y.; Shahsavar, F.; Eliasy, H. Colorectal cancer and the KIR genes in the human genome: A meta-analysis. Genom. Data 2016, 10, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portela, P.; Merzoni, J.; Lindenau, J.D.; Damin, D.C.; Wilson, T.J.; Roesler, R.; Schwartsmann, G.; Jobim, L.F.; Jobim, M. KIR genes and HLA class I ligands in a Caucasian Brazilian population with colorectal cancer. Hum. Immunol. 2017, 78, 263–268. [Google Scholar] [CrossRef]
- Barani, S.; Hosseini, S.V.; Ghaderi, A. Activating and inhibitory killer cell immunoglobulin like receptors (KIR) genes are involved in an increased susceptibility to colorectal adenocarcinoma and protection against invasion and metastasis. Immunobiology 2019, 224, 681–686. [Google Scholar] [CrossRef]
- Beksac, K.; Beksac, M.; Dalva, K.; Karaagaoglu, E.; Tirnaksiz, M.B. Impact of “Killer Immunoglobulin-Like Receptor /Ligand” Genotypes on Outcome following Surgery among Patients with Colorectal Cancer: Activating KIRs Are Associated with Long-Term Disease Free Survival. PLoS One 2015, 10, e0132526. [Google Scholar] [CrossRef]
- Dring, M.M.; Morrison, M.H.; McSharry, B.P.; Guinan, K.J.; Hagan, R.; O’Farrelly, C.; Gardiner, C.M. Innate immune genes synergize to predict increased risk of chronic disease in hepatitis C virus infection. Proc. Natl. Acad. Sci. United States Am. 2011, 108, 5736–5741. [Google Scholar] [CrossRef] [Green Version]
- Wauquier, N.; Padilla, C.; Becquart, P.; Leroy, E.; Vieillard, V. Association of KIR2DS1 and KIR2DS3 with fatal outcome in Ebola virus infection. Immunogenetics 2010, 62, 767–771. [Google Scholar] [CrossRef] [Green Version]
- VandenBussche, C.J.; Mulrooney, T.J.; Frazier, W.R.; Dakshanamurthy, S.; Hurley, C.K. Dramatically reduced surface expression of NK cell receptor KIR2DS3 is attributed to multiple residues throughout the molecule. Genes Immun. 2009, 10, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Sconocchia, G.; Eppenberger-Castori, S.; Zlobec, I.; Karamitopoulou, E.; Arriga, R.; Coppola, A.; Caratelli, S.; Spagnoli, G.C.; Lauro, D.; Lugli, A.; et al. HLA class II antigen expression in colorectal carcinoma tumors as a favorable prognostic marker. Neoplasia (United States) 2014, 16, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrity-Park, M.M.; Loftus, E.V.; Sandborn, W.J.; Bryant, S.C.; Smyrk, T.C. MHC class II alleles in ulcerative colitis-associated colorectal cancer. Gut 2009, 58, 1226–1233. [Google Scholar] [CrossRef] [PubMed]
- Aureli, A.; Canossi, A.; Del Beato, T.; Franceschilli, L.; Buonomo, O.; Papola, F.; De Sanctis, F.; Lanzilli, G.; Sileri, P.; Coppola, A.; et al. HLA-DRB1∗13:01 allele in the genetic susceptibility to colorectal carcinoma. Int. J. Cancer 2015, 136, 2464–2468. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, S.; Cariappa, A.; Tang, M.; Bell, D.; Haber, D.A.; Isselbacher, K.J.; Finkelstein, D.; Forcione, D.; Pillai, S. Genetic susceptibility to breast cancer: HLA DQB*03032 and HLA DRB1*11 may represent protective alleles. Proc. Natl. Acad. Sci. United States Am. 2000, 97, 11451–11454. [Google Scholar] [CrossRef] [Green Version]
- Arons, E.; Adams, S.; Venzon, D.J.; Pastan, I.; Kreitman, R.J. Class II human leucocyte antigen DRB1*11 in hairy cell leukaemia patients with and without haemolytic uraemic syndrome. Br. J. Haematol. 2014, 166, 729–738. [Google Scholar] [CrossRef]
- Meyer, D.; Vitor, V.R.; Bitarello, B.D.; Débora, D.Y.; Nunes, K. A genomic perspective on HLA evolution. Immunogenetics 2018, 70, 5–27. [Google Scholar] [CrossRef]



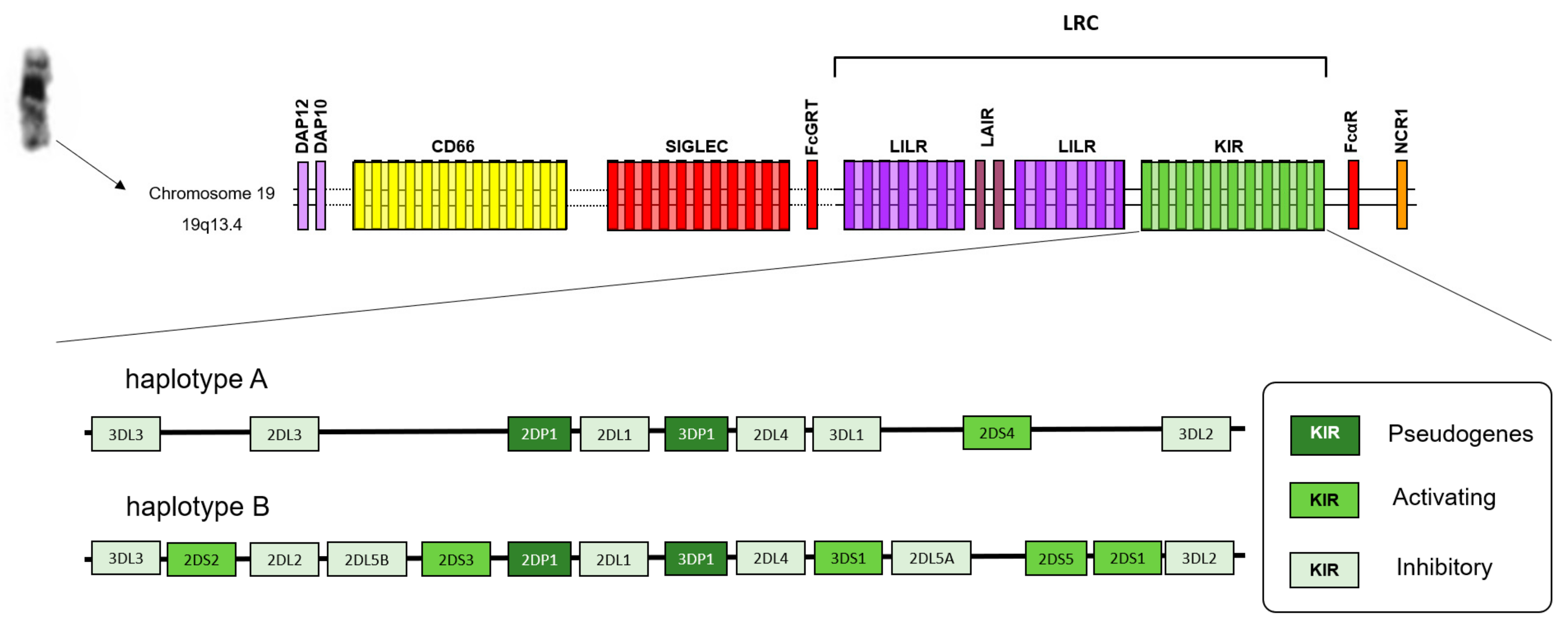{kind=link}
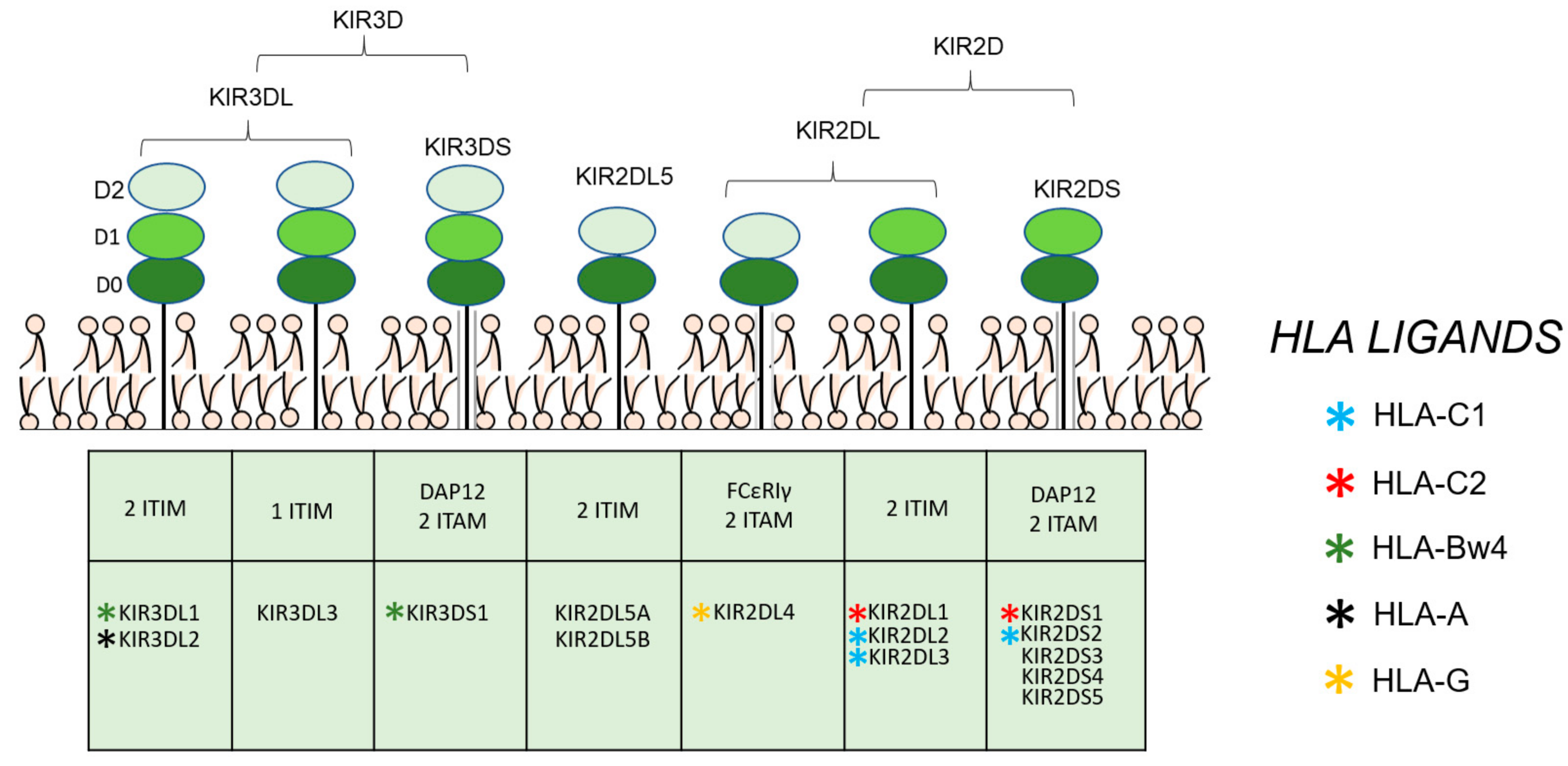{kind=link}
| CRC Patients (n = 943) | Healthy Controls (n = 1076) | |
|---|---|---|
| Age (median, range) | 69 (23–91) | 64.15 (24–92) |
| Gender Female Male | 338 (35.8%) 605 (64.2%) | 527 (48.98%) 549 (51.02%) |
| Smoke | ||
| Current smoker Former smoker Never smoker Unknown | 119 (12.6%) 386 (40.9%) 413 (43.8%) 26 (2.8%) | 180 (16.73%) 335 (31.13%) 553 (51.39%) 8 (0.74%) |
| TNM stage at diagnosis | ||
| I–II III IV Unknown | 418 (44.3%) 342 (36.3%) 131 (13.9%) 52 (5.5%) | |
| Tumor location | ||
| Rectum Sigma Cecum Ascending colon Sigmoid colon Descending colon Others* Unknown | 311 (33%) 168 (17.8%) 98 (10.4%) 81 (8.6%) 70 (7.4%) 52 (5.5%) 152 (16.1%) 11 (1.2%) | |
| Chr | SNP | Position | Associated Gene | A1 | MAF CRC Cases | MAF Healthy Controls | p Value | OR |
|---|---|---|---|---|---|---|---|---|
| 6 | rs16896742 | 29922740 | Intergenic | G | 0.31 | 0.3669 | 3.97 × 10−4 | 0.79 |
| 6 | rs28367832 | 31305731 | Intergenic | A | 0.42 | 0.4845 | 6.57 × 10−5 | 0.77 |
| 6 | rs9277952 | 33204274 | Intergenic | A | 0.15 | 0.1097 | 4.01 × 10−5 | 1.47 |
| HLA Allele | AF CRC Cases | AF Healthy Controls | P value | OR | 95% CI | Padj |
|---|---|---|---|---|---|---|
| DRB1*11:01 | 0.1002 | 0.0785 | 0.0137 | 1.32 | 1.06–1.65 | 0.0961 |
| C*07:01 | 0.1304 | 0.1543 | 0.0321 | 0.82 | 0.69–0.98 | 0.1774 |
| C*12:03 | 0.0689 | 0.0534 | 0.0443 | 1.30 | 1.01–1.67 | 0.1774 |
| A*02:01 | 0.2264 | 0.2519 | 0.0576 | 0.87 | 0.75–1.00 | 0.3453 |
| A*29:02 | 0.0848 | 0.0730 | 0.1554 | 1.18 | 0.94–1.50 | 0.4662 |
| DRB1*04:01 | 0.0467 | 0.0562 | 0.1751 | 0.82 | 0.62–1.09 | 0.6128 |
| B*35:01 | 0.0583 | 0.0511 | 0.3096 | 1.15 | 0.88–1.52 | 0.8689 |
| C*16:01 | 0.0843 | 0.0757 | 0.3136 | 1.13 | 0.89–1.42 | 0.6060 |
| B*44:03 | 0.1071 | 0.0976 | 0.3240 | 1.11 | 0.90–1.35 | 0.8689 |
| C*07:02 | 0.0817 | 0.0897 | 0.3641 | 0.90 | 0.72–1.13 | 0.6060 |
| Healthy Controls | CRC Cases | ||||||
|---|---|---|---|---|---|---|---|
| KIR Gene | n | % | n | % | p-value * | Pbonf | OR (95% CI) |
| KIR2DL1 | 1075 | 99.91 | 942 | 99.89 | NS | NS | |
| KIR2DL2 | 490 | 45.54 | 418 | 44.33 | NS | NS | |
| KIR2DL3 | 925 | 85.97 | 818 | 86.74 | NS | NS | |
| KIR2DL4 | 1076 | 100 | 943 | 100 | NS | NS | |
| KIR2DL5 | 500 | 46.47 | 450 | 47.72 | NS | NS | |
| KIR3DL1ex4 | 1030 | 95.72 | 911 | 96.61 | NS | NS | |
| KIR3DL1ex9 | 1032 | 95.91 | 911 | 96.61 | NS | NS | |
| KIR3DL2 | 1076 | 100 | 943 | 100 | NS | NS | |
| KIR2DS1 | 386 | 35.87 | 346 | 36.69 | NS | NS | |
| KIR2DS2 | 530 | 49.26 | 480 | 50.9 | NS | NS | |
| KIR2DS3 | 350 | 32.53 | 370 | 39.24 | 0.002 | 0.036 | 1.34 (1.12–1.61) |
| KIR2DS4total | 1030 | 95.72 | 909 | 96.39 | NS | NS | |
| KIR2DS4wt | 366 | 34.01 | 306 | 32.45 | NS | NS | |
| KIR2DS4del | 905 | 84.11 | 801 | 84.94 | NS | NS | |
| KIR2DS5 | 286 | 26.58 | 257 | 27.25 | NS | NS | |
| KIR3DS1 | 337 | 31.32 | 319 | 33.83 | NS | NS | |
| KIR2DP1 | 1074 | 99.81 | 943 | 100 | NS | NS | |
| KIR3DP1 | 1076 | 100 | 943 | 100 | NS | NS | |
| KIR Genotype | n | % | n | % | p-value * | OR (95% CI) | |
| AA | 280 | 26.02 | 243 | 26.77 | NS | ||
| Bx | 796 | 73.98 | 700 | 74.23 | NS | ||
| Healthy Controls | CRC Cases | |||||
|---|---|---|---|---|---|---|
| HLA Ligands | N | % | n | % | p-Value | OR (95% CI) |
| Bw4 | 726 | 67.47 | 650 | 68.93 | NS | |
| Bw6 | 885 | 82.25 | 766 | 81.23 | NS | |
| Bw4/Bw4 | 191 | 17.75 | 177 | 18.77 | NS | |
| Bw4/Bw6 | 535 | 49.72 | 473 | 50.16 | NS | |
| Bw6/Bw6 | 350 | 32.53 | 293 | 31.07 | NS | |
| Bw4I80 | 416 | 38.66 | 361 | 38.28 | NS | |
| Bw4T80 | 402 | 37.36 | 373 | 39.55 | NS | |
| HLA-C1 | 890 | 82.71 | 752 | 79.75 | NS | |
| HLA-C2 | 725 | 67.38 | 640 | 67.87 | NS | |
| HLA-C1C1 | 351 | 32.62 | 303 | 32.13 | NS | |
| HLA-C1C2 | 539 | 50.09 | 449 | 47.61 | NS | |
| HLA-C2C2 | 186 | 17.29 | 191 | 20.25 | NS | |
| KIR genotypes | N | % | n | % | p-value | OR (95% CI) |
| KIR3DL1/KIR3DL1 | 738 | 68.59 | 624 | 66.17 | NS | |
| KIR3DL1/KIR3DS1 | 294 | 27.32 | 287 | 30.43 | NS | |
| KIR3DS1/KIR3DS1 | 43 | 4.00 | 32 | 3.39 | NS | |
| KIR2DL2/KIR2DL2 | 149 | 13.85 | 125 | 13.26 | NS | |
| KIR2DL2/KIR2DL3 | 341 | 31.69 | 293 | 31.07 | NS | |
| KIR2DL3/KIR2DL3 | 584 | 54.28 | 525 | 55.67 | NS | |
| NS | ||||||
| KIR ligand associations | N | % | n | % | p-value | OR (95% CI) |
| KIR3DS1-Bw4I80 | 131 | 12.17 | 114 | 12.09 | NS | |
| KIR3DS1-Bw4T80 | 125 | 11.62 | 123 | 13.04 | NS | |
| KIR3DS1-Bw4 | 228 | 21.19 | 206 | 21.85 | NS | |
| KIR3DS1-Bw6 | 285 | 26.49 | 264 | 27.99 | NS | |
| KIR3DL1-Bw4I80 | 402 | 37.36 | 349 | 37.01 | NS | |
| KIR3DL1-Bw4T80 | 386 | 35.87 | 356 | 37.75 | NS | |
| KIR3DL1-Bw4 | 697 | 64.78 | 626 | 66.38 | NS | |
| KIR3DL1-Bw6 | 847 | 78.72 | 742 | 78.69 | NS | |
| KIR2DL1/HLA-C1C1 | 351 | 32.62 | 302 | 32.03 | NS | |
| KIR2DL1/HLA-C1C2 | 538 | 50.00 | 449 | 47.61 | NS | |
| KIR2DL1/HLA-C2C2 | 186 | 17.29 | 191 | 20.25 | NS | |
| KIR2DL2/HLA-C1C1 | 149 | 13.85 | 125 | 13.26 | NS | |
| KIR2DL2/HLA-C1C2 | 189 | 17.57 | 153 | 16.22 | NS | |
| KIR2DL2/HLA-C2C2 | 152 | 14.13 | 140 | 14.85 | NS | |
| KIR2DL3/HLA-C1C1 | 339 | 31.51 | 297 | 31.50 | NS | |
| KIR2DL3/HLA-C1C2 | 403 | 37.45 | 337 | 35.74 | NS | |
| KIR2DL3/HLA-C2C2 | 183 | 17.01 | 184 | 19.51 | NS | |
| KIR2DS1/HLA-C1C1 | 115 | 10.69 | 102 | 10.82 | NS | |
| KIR2DS1/HLA-C1C2 | 194 | 18.03 | 175 | 18.56 | NS | |
| KIR2DS1/HLA-C2C2 | 77 | 7.16 | 69 | 7.32 | NS | |
| Reference | Type of Experiment/Objective | Conclusions |
|---|---|---|
| [19] | 109 CRC patients (70 bladder and 34 laryngeal) and 100 controls. HLA and KIR genotyping. | No differences in KIR/HLA frequencies was observed between patients and controls. |
| [20] | 128 CRC patients and 255 controls. KIR and HLA genotyping. | The data showed no significant differences between KIR gene frequencies in CRC patients versus controls. |
| [21] | 241 CRC patients and 159 controls from Korean populations. KIR and HLA-C genotyping | The activating KIR2DS5 was more frequent in Korean CRC patients, showing a risk for the disease. The frequencies of KIR3DL1, KIR2DS2 and KIR2DS4 were lower in the rectal cancer subgroup, and they could have a protective effect against CRC. Also, the lower frequency of KIR2DS2 in patients with HLA-C1 homozygote, may be a protective effect too. |
| [22] | 52 CRC patients and 70 controls from Saudi population. KIR and HLA-C genotyping. | Activating KIRs (2DS1, 2DS2, 2DS3, 2DS5 and 3DS1) was more frequent in CRC patients, suggesting their presence a risk for disease. |
| [23] | 470 CRC patients and 483 controls. KIR genotyping. | The presence of KIR2DS5 was associated with CRC like as a non-protective gene. This result explains the inflammatory basis of this cancer. |
| [24] | 154 CRC patients and 216 controls from Caucasian Brazilian population. KIR and HLA genotyping. | No associations between KIRs and HLA in CRC was observed. However, the Bx haplotype was more frequent in controls than in patients, being a possible mechanism of protection to CRC. |
| [25] | 165 colorectal adenocarcinoma patients and 165 controls. KIR genotyping. | The presence of activating KIRs (≥ 4) and KIR3DL1, 3DS1, 2DS1 and 2DS4, were associated with protection against metastasis in CRC patients. |
| [26] | 29 CRC recurrent patients (in 5 years) vs. 58 CRC non-recurrent patients (in 5 years) after surgery and 154 controls. KIR and HLA-class I genotyping. | The increment of activating KIRs (in particularly 2DS2 and 2DS3) and the lack of inhibitory KIRs (in particularly 2DL1) was associated with long term disease-free survival and this was independent of tumor localization or stage. Also, HLA-A-Bw4 was associated with recurrent disease. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaz-Peña, R.; Mondelo-Macía, P.; Molina de la Torre, A.J.; Sanz-Pamplona, R.; Moreno, V.; Martín, V. Analysis of Killer Immunoglobulin-Like Receptor Genes in Colorectal Cancer. Cells 2020, 9, 514. https://doi.org/10.3390/cells9020514
Diaz-Peña R, Mondelo-Macía P, Molina de la Torre AJ, Sanz-Pamplona R, Moreno V, Martín V. Analysis of Killer Immunoglobulin-Like Receptor Genes in Colorectal Cancer. Cells. 2020; 9(2):514. https://doi.org/10.3390/cells9020514
Chicago/Turabian StyleDiaz-Peña, Roberto, Patricia Mondelo-Macía, Antonio José Molina de la Torre, Rebeca Sanz-Pamplona, Víctor Moreno, and Vicente Martín. 2020. "Analysis of Killer Immunoglobulin-Like Receptor Genes in Colorectal Cancer" Cells 9, no. 2: 514. https://doi.org/10.3390/cells9020514
APA StyleDiaz-Peña, R., Mondelo-Macía, P., Molina de la Torre, A. J., Sanz-Pamplona, R., Moreno, V., & Martín, V. (2020). Analysis of Killer Immunoglobulin-Like Receptor Genes in Colorectal Cancer. Cells, 9(2), 514. https://doi.org/10.3390/cells9020514