In Vitro Generation of Oocyte Like Cells and Their In Vivo Efficacy: How Far We have been Succeeded


Abstract
:1. Introduction
2. Ovarian Stem Cells (OSCs) Isolation and Their Differentiation Potential
3. Ectopic Expression/Overexpression of Germ Cell-Specific Markers
4. Follicular Fluid: A Magical OLC Differentiation Inducer
5. Skin-Derived Stem Cells
6. Co-Culturing as an Efficient Method
7. Very Small Embryonic-Like Stem Cells (VSELS): New Heroes in Gamete Research
8. Direct Utilization of Undifferentiated Stem Cells in Curing Fertility Disorders
9. In Vivo Efficacy of the In Vitro Differentiated OLCs
10. Selection of Best OLC Source for Fertility Treatment
11. Present Complications and Future Expectations
12. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bukovsky, A.; Svetlikova, M.; Caudle, M.R. Oogenesis in cultures derived from adult human ovaries. Reprod. Biol. Endocrinol. 2005, 1, 17. [Google Scholar] [CrossRef] [Green Version]
- Virant-Klun, I.; Zech, N.; Rozman, P.; Vogler, A.; Cvjeticanin, B.; Klemenc, P.; Malicev, E.; Meden-Vrtovec, H. Putative stem cells with an embryonic character isolated from the ovarian surface epithelium of women with no naturally present follicles and oocytes. Differentiation 2008, 76, 843–856. [Google Scholar] [CrossRef]
- Fowler, P.A.; Flannigan, S.; Mathers, A.; Gillanders, K.; Lea, R.G.; Wood, M.J.; Maheshwari, A.; Bhattacharya, S.; Collie-Duguid, E.S.; Baker, P.J.; et al. Gene expression analysis of human fetal ovarian primordial follicle formation. J. Clin. Endocrinol. Metab. 2009, 94, 1427–1435. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Huan, Y.; Guo, L.; Cheng, W.; Zou, G. CD44+/CD105+ Human Amniotic Fluid Mesenchymal Stem Cells Survive and Proliferate in the Ovary Long-Term in a Mouse Model of Chemotherapy-Induced Premature Ovarian Failure. Int. J. Med. Sci. 2012, 9, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Virant-Klun, I.; Skutella, T.; Hren, M.; Gruden, K.; Cvjeticanin, B.; Vogler, A.; Sinkovec, J. Isolation of small SSEA-4-positive putative stem cells from the ovarian surface epithelium of adult human ovaries by two different methods. BioMed. Res. Int. 2013, 2013, 690415. [Google Scholar] [CrossRef] [Green Version]
- White, Y.A.; Woods, D.C.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J.L. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Qiu, P.; Bai, Y.; Pan, S.; Li, W.; Liu, W.; Hua, J. Gender dependent potentiality of differentiation of human umbilical cord mesenchymal stem cells into oocyte-like cells in vitro. Cell Biochem. Funct. 2013, 31, 365–373. [Google Scholar] [CrossRef]
- Liu, T.; Huang, Y.; Bu, Z.; Zhao, Y.; Zou, G.; Liu, Z. Induction of E-cadherin+ human amniotic fluid cell differentiation into oocyte-like cells via culture in medium supplemented with follicular fluid. Mol. Med. Rep. 2014, 10, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Huang, Y.; Zhang, J.; Qin, W.; Chi, H.; Chen, J.; Yu, Z.; Chen, C. Transplantation of human menstrual blood stem cells to treat premature ovarian failure in mouse model. Stem Cells Dev. 2014, 23, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Latifpour, M.; Shakiba, Y.; Amidi, F.; Mazaheri, Z.; Sobhani, A. Differentiation of human umbilical cord matrix-derived mesenchymal stem cells into germ-like cells. Avicenna J. Med. Biotech. 2014, 6, 218–227. [Google Scholar]
- Yu, X.; Wang, N.; Qiang, R.; Wan, Q.; Qin, M.; Chen, S.; Wang, H. Human amniotic fluid stem cells possess the potential to differentiate into primordial follicle oocytes in vitro. Biol. Reprod. 2014, 90, 73. [Google Scholar] [CrossRef] [Green Version]
- Asgari, H.R.; Akbari, M.; Abbasi, M.; Ai, J.; Korouji, M.; Aliakbari, F.; Babatunde, A.K.; Sargolzaei, A.F.; Joghataei, M.T. Human Wharton’s jelly-derived mesenchymal stem cells express oocyte developmental genes during co-culture with placental cells. Iran. J. Basic Med. Sci. 2015, 18, 22–29. [Google Scholar]
- Ge, W.; Ma, H.G.; Cheng, S.F.; Sun, Y.C.; Sun, L.L.; Sun, X.F.; Li, L.; Dyce, P.; Li, J.; Shi, Q.H.; et al. Differentiation of early germ cells from human skin-derived stem cells without exogenous gene integration. Sci. Rep. 2015, 5, 13822. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Lu, H.; Cao, S.; Deng, Y.L.; Li, Q.J.; Wan, Q.; Yie, S.M. Stem cells derived from human first-trimester umbilical cord have the potential to differentiate into oocyte-like cells in vitro. Int. J. Mol. Med. 2015, 35, 1219–1229. [Google Scholar] [CrossRef] [Green Version]
- Lai, D.; Guo, Y.; Zhang, Q.; Chen, Y.; Xiang, C. Differentiation of human menstrual blood-derived endometrial mesenchymal stem cells into oocyte-like cells. Acta Biochim. Biophys. Sin. 2016, 48, 998–1005. [Google Scholar] [CrossRef] [Green Version]
- Jung, D.; Xiong, J.; Ye, M.; Qin, X.; Li, L.; Cheng, S.; Luo, M.; Peng, J.; Dong, J.; Tang, F.; et al. In vitro differentiation of human embryonic stem cells into ovarian follicle-like cells. Nat. Commun. 2017, 12, 15680. [Google Scholar] [CrossRef]
- Qing, T.; Shi, Y.; Qin, H.; Ye, X.; Wei, W.; Liu, H.; Ding, M.; Deng, H. Induction of oocyte-like cells from mouse embryonic stem cells by co-culture with ovarian granulosa cells. Differentiation 2007, 75, 902–911. [Google Scholar] [CrossRef]
- Dyce, P.W.; Liu, J.; Tayade, C.; Kidder, G.M.; Betts, D.H.; Li, J. In vitro and in vivo germ line potential of stem cells derived from newborn mouse skin. PLoS ONE 2011, 6, e20339. [Google Scholar] [CrossRef] [Green Version]
- Dyce, P.W. Differentiation of newborn mouse skin derived stem cells into germ-like cells in vitro. J. Vis. Exp. 2013, 77, e50486. [Google Scholar] [CrossRef] [Green Version]
- Bahmanpour, S.; Zarei Fard, N.; Talaei-Khozani, T.; Hosseini, A.; Esmaeilpour, T. Effect of BMP4 preceded by retinoic acid and co-culturing ovarian somatic cells on differentiation of mouse embryonic stem cells into oocyte-like cells. Dev. Growth Differ. 2015, 57, 378–388. [Google Scholar] [CrossRef]
- Tan, H.; Wang, J.J.; Cheng, S.F.; Ge, W.; Sun, X.C.; Sun, X.F.; Sun, R.; Li, L.; Li, B.; Shen, W. Retinoic acid promotes the proliferation of primordial germ cell–like cells differentiated from mouse skin-derived stem cells in vitro. Theriogenology. 2016, 85, 408–4018. [Google Scholar] [CrossRef]
- Parvari, S.; Yazdekhasti, H.; Rajabi, Z.; Malek, V.G.; Rastegar, T.; Abbasi, M. Differentiation of mouse ovarian stem cells toward oocyte-like structure by co-culture with granulosa cells. Cell Reprogram. 2016, 18, 419–428. [Google Scholar] [CrossRef]
- Liu, J.; Shang, D.; Xiao, Y.; Zhong, P.; Cheng, H.; Zhou, R. Isolation and characterization of string-forming female germline stem cells from ovaries of neonatal mice. J. Biol. Chem. 2017, 292, 16003–16013. [Google Scholar] [CrossRef] [Green Version]
- Dyce, P.W.; Tenn, N.; Kidder, G.M. Retinoic acid enhances germ cell differentiation of mouse skin-derived stem cells. J. Ovarian Res. 2018, 11, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyce, P.W.; Wen, L.; Li, J. In vitro germline potential of stem cells derived from fetal porcine skin. Nat. Cell Biol. 2006, 8, 384–391. [Google Scholar] [CrossRef]
- Dyce, P.W.; Shen, W.; Huynh, E.; Shao, H.; Villagómez, D.A.; Kidder, G.M.; King, W.A.; Li, J. Analysis of oocyte-like cells differentiated from porcine fetal-skin derived stem cells. Stem Cells Dev. 2011, 20, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Song, S.H.; Kumar, B.M.; Kang, E.J.; Lee, Y.M.; Kim, T.H.; Ock, S.A.; Lee, S.L.; Jeon, B.G.; Rho, G.J. Characterization of porcine multipotent stem/stromal cells derived from skin, adipose, and ovarian tissues and their differentiation in vitro into putative oocyte-like cells. Stem Cells Dev. 2011, 20, 1359–1370. [Google Scholar] [CrossRef]
- Park, B.W.; Shen, W.; Linher-Melville, K.; Li, J. Deleted in azoospermia-like enhances in vitro derived porcine germ cell formation and meiosis. Stem Cells Dev. 2013, 22, 939–950. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.M.; Kim, T.H.; Lee, J.H.; Lee, W.J.; Jeon, R.H.; Jang, S.J.; Ock, S.A.; Lee, S.L.; Park, B.W.; Rho, G.J. Overexpression of Oct4 in porcine ovarian stem/stromal cells enhances differentiation of oocyte-like cells in vitro and ovarian follicular formation in vivo. J. Ovarian Res. 2016, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.J.N.; Souza, G.B.; Bernardo, J.M.P.; Ribeiro, R.P.; Passos, J.R.S.; Bezerra, F.T.G.; Saraiva, M.V.A.; Silva, J.R.V. Expression of markers for germ cells and oocytes in cow dermal fibroblast treated with 5-azacytidine and cultured in differentiation medium containing BMP2, BMP4 and follicular fluid. Zygote 2017, 25, 341–357. [Google Scholar] [CrossRef]
- de Souza, G.B.; Costa, J.; da Cunha, E.V.; Passos, J.; Ribeiro, R.P.; Saraiva, M.; van den Hurk, R.; Silva, J. Bovine ovarian stem cells differentiate into germ cells and oocyte-like structures after culture in vitro. Reprod. Domest. Anim. 2017, 52, 243–250. [Google Scholar] [CrossRef]
- Singhal, D.K.; Singhal, R.; Malik, H.N.; Surender Singh, S.; Kumar, S.; Kaushik, J.K.; Mohanty, A.K.; Malakar, D. Generation of Germ Cell-Like Cells and Oocyte-Like Cells from Goat Induced Pluripotent Stem Cells. J. Stem Cell Res. Ther. 2015, 5, 279. [Google Scholar] [CrossRef]
- Zou, K.; Yuan, Z.; Yang, Z.; Luo, H.; Sun, K.; Zhou, L.; Xiang, J.; Shi, L.; Yu, Q.; Zhang, Y.; et al. Production of offspring from a germline stem cell line derived from neonatal ovaries. Nat. Cell Biol. 2009, 11, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Hou, L.; Sun, K.; Xie, W.; Wu, J. Improved efficiency of female germline stem cell purification using fragilis-based magnetic bead sorting. Stem Cells Dev. 2011, 20, 2197–2204. [Google Scholar] [CrossRef] [PubMed]
- Bui, H.T.; Thuan, N.V.; Kwon, D.N.; Choi, Y.J.; Kang, M.H.; Han, J.W.; Kim, T.; Kim, J.H. Identification and characterization of putative stem cells in the adult pig ovary. Development 2014, 141, 2235–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revelli, A.; Piane, L.D.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Sun, Y.C.; Ge, W.; Tan, H.; Cheng, S.F.; Yin, S.; Sun, X.F.; Li, L.; Dyce, P.; Li, J.; et al. The crucial role of Activin A on the formation of primordial germ cell-like cells from skin-derived stem cells in vitro. Cell Cycle 2015, 14, 3016–3029. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Fong, C.Y.; Biswas, A.; Bongso, A. Comparative characterization of cells from the various compartments of the human umbilical cord shows that the Wharton’s Jelly compartment provides the best source of clinically utilizable mesenchymal stem cells. PLoS ONE 2015, 10, e0127992. [Google Scholar] [CrossRef]
- Ullah, I.; Raghavendra Baregundi Subbarao, R.B.; Kim, E.J.; Bharti, D.; Jang, S.J.; Park, J.S.; Shivakumar, S.B.; Lee, S.L.; Kang, D.; Byun, J.H.; et al. In vitro comparative analysis of human dental stem cells from a single donor and its neuronal differentiation potential evaluated by electrophysiology. Life Sci. 2016, 154, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Bharti, D.; Shivakumar, S.B.; Park, J.K.; Ullah, I.; Subbarao, R.B.; Park, J.S.; Lee, S.L.; Park, B.W.; Rho, G.J. Comparative analysis of human Wharton’s jelly mesenchymal stem cells derived from different parts of the same umbilical cord. Cell Tissue Res. 2018, 372, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Makoolati, Z.; Movahedin, M.; Forouzandeh-Moghadam, M. Bone morphogenetic protein 4 is an efficient inducer for mouse embryonic stem cell differentiation into primordial germ cell. In Vitro Cell Dev. Biol. Anim. 2011, 47, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Ji, P.; Cao, J.; Zhu, S.; Li, Y.; Zheng, L.; Chen, X.; Feng, L. Dazl promotes germ cell differentiation from embryonic stem cells. J. Mol. Cell Biol. 2009, 1, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Ma, Y.; Wang, F.; Hu, L.; Sun, Y. miR-644-5p carried by bone mesenchymal stem cell-derived exosomes targets regulation of p53 to inhibit ovarian granulosa cell apoptosis. Stem Cell Res. Ther. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Sun, J.; Huang, Y.; Bu, S.; Guo, Y.; Gu, T.; Li, B.; Wang, C.; Lai, D. Human amniotic epithelial cell-derived exosomes restore ovarian function by transferring microRNAs against apoptosis. Mol. Ther. Nucleic Acids 2019, 16, 407–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sriraman, K.; Bhartiya, D.; Anand, S.; Bhutda, S. Mouse ovarian very small embryonic-like stem cells resist chemotherapy and retain ability to initiate oocyte-specific differentiation. Reprod. Sci. 2015, 22, 884–903. [Google Scholar] [CrossRef] [Green Version]
- Lai, D.; Wang, F.; Dong, Z.; Zhang, Q. Skin-derived mesenchymal stem cells help restore function to ovaries in a premature ovarian failure mouse model. PLoS ONE 2014, 9, e98749. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, H.; Zhang, Y.; Li, N.; Wen, Y.; Cao, F.; Ai, H.; Xue, X. Homing and restorative effects of bone marrow-derived mesenchymal stem cells on cisplatin injured ovaries in rats. Mol. Cells 2014, 37, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Badawy, A.; Sobh, M.A.; Ahdy, M.; Abdelhafez, M.S. Bone marrow mesenchymal stem cell repair of cyclophosphamide-induced ovarian insufficiency in a mouse model. Int. J. Womens Health 2017, 9, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.H.; Choi, J.H.; Jun, Y.; Lim, S.M.; Park, S.; Paek, J.Y.; Lee, S.H.; Hwang, J.Y.; Kim, G.J. 3D-cultured human placenta derived mesenchymal stem cell spheroids enhance ovary function by inducing folliculogenesis. Sci. Rep. 2018, 8, 15313. [Google Scholar] [CrossRef]
- Li, J.; Yu, Q.; Huang, H.; Deng, W.; Cao, X.; Adu-Frimpong, M.; Yu, J.; Xu, X. Human chorionic plate-derived mesenchymal stem cells transplantation restores ovarian function in a chemotherapy-induced mouse. Stem Cell Res. Ther. 2018, 9, 81. [Google Scholar] [CrossRef]
- Feng, P.; Li, P.; Tan, J. Human menstrual blood-derived stromal cells promote recovery of premature ovarian insufficiency via regulating the ECM-dependent FAK/AKT signaling. Stem Cell Rev. Rep. 2019, 15, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalalie, L.; Rezaie, M.J.; Jalili, A.; Rezaee, M.A.; Vahabzadeh, Z.; Rahmani, M.R.; Karimipoor, M.; Hakhamaneshi, M.S. Distribution of the CM-Dil-labeled human umbilical cord vein mesenchymal stem cells migrated to the cyclophosphamide-injured ovaries in C57BL/6 mice. Iran. Biomed. J. 2019, 23, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Zhang, X.; Fan, Z.; Wang, Y.; Yao, G.; Wan, X.; Liu, Z.; Yang, B.; Yu, L. Human amniotic mesenchymal stem cells improve the follicular microenvironment to recover ovarian function in premature ovarian failure mice. Stem Cell Res. Ther. 2019, 10, 299. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.A.; Shalaby, S.; Brakta, S.; Elam, L.; Elsharoud, A.; Al-Hendy, A. Umbilical cord blood mesenchymal stem cells as an infertility treatment for chemotherapy induced premature ovarian insufficiency. Biomedicines 2019, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Lin, L.; Sha, C.; Li, T.; Zhao, D.; Wei, H.; Chen, Q.; Liu, Y.; Chen, X.; Xu, W.; et al. Bone marrow mesenchymal stem cell-derived exosomal miR-144-5p improves rat ovarian function after chemotherapy-induced ovarian failure by targeting PTEN. Lab. Investig. 2019. [Google Scholar] [CrossRef]
- Yang, Z.; Du, X.; Wang, C.; Zhang, J.; Liu, C.; Li, Y.; Jiang, H. Therapeutic effects of human umbilical cord mesenchymal stem cell-derived micro-vesicles on premature ovarian insufficiency in mice. Stem Cell Res. Ther. 2019, 10, 250. [Google Scholar] [CrossRef] [Green Version]
- Silvestris, E.; Cafforio, P.; D’Oronzo, S.; Felici, C.; Silvestris, F.; Loverro, G. In vitro differentiation of human oocyte-like cells from oogonial stem cells: Single-cell isolation and molecular characterization. Hum. Reprod. 2018, 33, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Clarkson, Y.L.; McLaughlin, M.; Waterfall, M.; Dunlop, C.E.; Skehel, P.A.; Anderson, R.A.; Telfer, E.E. Initial characterization of adult human ovarian cell populations isolated by DDX4 expression and aldehyde dehydrogenase activity. Sci. Rep. 2018, 8, 6953. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, Z.; Yang, Y.; Wang, S.; Shi, L.; Xie, W.; Sun, K.; Zou, K.; Wang, L.; Xiong, J.; et al. Production of transgenic mice by random recombination of targeted genes in female germline stem cells. J. Mol. Cell Biol. 2011, 3, 132–141. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, L.; Kang, J.X.; Xie, W.; Li, X.; Wu, C.; Xu, B.; Wu, J. Production of fat-1 transgenic rats using a post-natal female germline stem cell line. Mol. Hum. Reprod. 2014, 20, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Parvari, S.; Abbasi, M.; Abbasi, N.; Malek, V.G.; Amidi, F.; Aval, F.S.; Roudkenar, M.H.; Izadyar, F. Stem cell isolation by a morphology-based selection method in postnatal mouse ovary. Arch. Med. Sci. 2015, 11, 670–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkson, Y.L.; Weatherall, E.; Waterfall, M.; McLaughlin, M.; Lu, H.; Skehel, P.A.; Anderson, R.A.; Telfer, E.E. Extracellular Localization of the C-Terminus of DDX4 Confirmed by Immunocytochemistry and Fluorescence-Activated Cell Sorting. Cells 2019, 8, 578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Panula, S.; Petropoulos, S.; Edsgärd, D.; Busayavalasa, K.; Liu, L.; Li, X.; Risal, S.; Shen, Y.; Shao, J.; et al. Adult human and mouse ovaries lack DDX4-expressing functional oogonial stem cells. Nat. Med. 2015, 21, 1116–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarate-Garcia, L.; Lane, S.I.; Merriman, J.A.; Jones, K.T. FACS-sorted putative oogonial stem cells from the ovary are neither DDX4-positive nor germ cells. Sci. Rep. 2016, 6, 27991. [Google Scholar] [CrossRef] [Green Version]
- Woods, D.C.; White, Y.A.R.; Tilly, J.L. Purification of oogonial stem cells from adult mouse and human ovaries: An assessment of the literature and a view toward the future. Reprod. Sci. 2013, 20, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, S.F.; Vahidi, N.A.; Park, S.; Weitzel, R.P.; Tisdale, J.; Rueda, B.R.; Wolff, E.F. Characterization of extracellular DDX4-or Ddx4-positive ovarian cells. Nat. Med. 2015, 21, 1114–1116. [Google Scholar] [CrossRef]
- Guan, K.; Wolf, F.; Becker, A.; Engel, W.; Nayernia, K.; Hasenfuss, G. Isolation and cultivation of stem cells from adult mouse testes. Nat. Protoc. 2009, 4, 143–154. [Google Scholar] [CrossRef]
- Yamanaka, S.; Blau, H.M. Nuclear reprogramming to a pluripotent state by three approaches. Nature 2010, 465, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Xie, M.; Laurent, T.; Ding, S. Progress in the reprogramming of somatic cells. Circ. Res. 2013, 112, 562–574. [Google Scholar] [CrossRef] [Green Version]
- Ieda, M.; Fu, J.D.; Delgado-Olguin, P.; Vedantham, V.; Hayashi, Y.; Bruneau, B.G.; Srivastava, D. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell 2010, 142, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Magnúsdóttir, E.; Dietmann, S.; Murakami, K.; Günesdogan, U.; Tang, F.; Bao, S.; Diamanti, E.; Lao, K.; Gottgens, B.; Azim-Surani, M. A tripartite transcription factor network regulates primordial germ cell specification in mice. Nat. Cell. Biol. 2013, 15, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Sterneckert, J.; Höing, S.; Schöler, H.R. Concise Review: Oct4 and More: The Reprogramming Expressway. Stem Cells 2012, 30, 15–21. [Google Scholar] [CrossRef]
- Lin, I.Y.; Chiu, F.L.; Yeang, C.H.; Chen, H.F.; Chuang, C.Y.; Yang, S.Y.; Hou, P.S.; Sintupisut, N.; Ho, H.N.; Kuo, H.C.; et al. Suppression of the SOX2 Neural Effector Gene by PRDM1 Promotes Human Germ Cell Fate in Embryonic Stem Cells. Stem Cell Rep. 2014, 2, 189–204. [Google Scholar] [CrossRef] [Green Version]
- Nakaki, F.; Hayashi, K.; Ohta, H.; Kurimoto, K.; Yabuta, Y.; Saitou, M. Induction of mouse germ-cell fate by transcription factors in vitro. Nature 2013, 501, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Irie, N.; Weinberger, L.; Tang, W.W.; Kobayashi, T.; Viukov, S.; Manor, Y.S.; Dietmann, S.; Hanna, J.H.; Surani, M.A. SOX17 is a critical specifier of human primordial germ cell fate. Cell 2015, 160, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Saitou, M.; Barton, S.C.; Surani, M.A. A molecular programme for the specification of germ cell fate in mice. Nature 2002, 418, 293–300. [Google Scholar] [CrossRef]
- Soyal, S.M.; Amleh, A.; Dean, J. FIGalpha, a germ cell-specific transcription factor required for ovarian follicle formation. Development 2000, 127, 4645–4654. [Google Scholar]
- Zhao, H.; Chen, Z.J.; Qin, Y.; Shi, Y.; Wang, S.; Choi, Y.; Simpson, J.L.; Rajkovic, A. Transcription factor FIGLA is mutated in patients with premature ovarian failure. Am. J. Hum. Genet. 2008, 82, 1342–1348. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Gill, M.E.; Koubova, J.; Page, D.C. Germ cell-intrinsic and -extrinsic factors govern meiotic initiation in mouse embryos. Science 2008, 322, 1685–1687. [Google Scholar] [CrossRef] [Green Version]
- Yen, P.H. Putative biological functions of the DAZ family. Int. J. Androl. 2004, 27, 125–129. [Google Scholar] [CrossRef]
- Lin, Y.; Page, D.C. Dazl deficiency leads to embryonic arrest of germ cell development in XY C57BL/6 mice. Dev. Biol. 2005, 288, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Park, T.S.; Galic, Z.; Conway, A.E.; Lindgren, A.; van Handel, B.J.; Magnusson, M.; Richter, L.; Teitell, M.A.; Mikkola, H.K.; Lowry, W.E.; et al. Derivation of primordial germ cells from human embryonic and induced pluripotent stem cells is significantly improved by co-culture with human fetal gonadal cells. Stem Cells 2009, 27, 783–795. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.H.; Nhu, B.L.Q.; Uyen, N.N.P.; Nguyen, V.T.; Bui, H.T. Isolation of female germline stem cells from porcine ovarian tissue and differentiation into oocyte-like cells. J. Reprod. Dev. 2019, 65, 423–432. [Google Scholar] [CrossRef]
- Bahmanpour, S.; Keshavarz, A.; Zarei Fard, N. Effect of different concentrations of forskolin along with mature granulosa cell co-culturing on mouse embryonic stem cell differentiation into germ-like cells. Iran. Biomed. J. 2020, 24, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T.; Kaga, Y.; Ohta, H.; Odamoto, M.; Sekita, Y.; Li, K.; Yamano, N.; Fujikawa, K.; Isotani, A.; Sasaki, N.; et al. Induction of primordial germ cell-like cells from mouse embryonic stem cells by ERK signal inhibition. Stem Cells 2014, 32, 2668–2678. [Google Scholar] [CrossRef]
- Zou, K.; Wang, J.; Bi, H.; Zhang, Y.; Tian, X.; Tian, N.; Ma, W.; Wu, J. Comparison of different in vitro differentiation conditions for murine female germline stem cells. Cell Prolif. 2019, 52, e12530. [Google Scholar] [CrossRef]
- Gaafar, T.M.; Hanna, M.O.F.; Hammady, M.R.; Amr, H.M.; Osman, O.M.; Nasef, A.; Osman, A.M. Evaluation of cytokines in follicular fluid and their effect on fertilization and pregnancy outcome. Immunol. Investig. 2014, 43, 572–584. [Google Scholar] [CrossRef]
- Shen, X.; Liu, X.; Zhu, P.; Zhang, Y.; Wang, J.; Wang, Y.; Wang, W.; Liu, J.; Li, N.; Liu, F. Proteomic analysis of human follicular fluid associated with successful in vitro fertilization. Reprod. Biol. Endocrinol. 2017, 15, 58. [Google Scholar] [CrossRef] [Green Version]
- Dalman, A.; Totonchi, M.; Rezazadeh, V.M. Human ovarian theca-derived multipotent stem cells have the potential to differentiate into oocyte-like cells in vitro. Cell J. 2019, 20, 527–536. [Google Scholar] [CrossRef]
- Moghadam, A.R.E.; Moghadam, M.T.; Saki, G.; Nikbakht, R. Bone morphogenetic protein 15 induces differentiation of mesenchymal stem cell derived from human follicular fluid to oocyte like cell. Peer J. Preprints. Unpublished work. 2019, 7, e28006v1. [Google Scholar] [CrossRef]
- Chen, M.; Przyborowski, M.; Berthiaume, F. Stem cells for skin tissue engineering and wound healing. Crit. Rev. Biomed. Eng. 2009, 37, 399–421. [Google Scholar] [CrossRef] [Green Version]
- Linher, K.; Dyce, P.; Li, J. Primordial germ cell-like cells differentiated in vitro from skin-derived stem cells. PLoS ONE 2009, 4, e8263. [Google Scholar] [CrossRef] [Green Version]
- Strauss, J.F.; Modi, B.; McAllister, J.M. Defects in Ovarian Steroid Hormone Biosynthesis. In Cellular Endocrinology in Health and Disease; Ulloa-Aguirre, A., Conn, P.M., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 285–309. [Google Scholar] [CrossRef]
- Cheng, X.; Chen, S.; Yu, X.; Zheng, P.; Wang, H. BMP15 gene is activated during human amniotic fluid stem cell differentiation into oocyte-like cells. DNA Cell Biol. 2012, 31, 1198–1204. [Google Scholar] [CrossRef]
- Panula, S.; Reda, A.; Stukenborg, J.B.; Ramathal, C.; Sukhwani, M.; Albalushi, H.; Edsgärd, D.; Nakamura, M.; Söder, O.; Orwig, K.E.; et al. Over expression of NANOS3 and DAZL in human embryonic stem cells. PLoS ONE 2016, 11, e0165268. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Sun, J.; Wang, J.; Wang, L.; Bai, Y.; Yu, M.; Lian, Z.; Zhang, S.; Hua, J. Characterization of female germ-like cells derived from mouse embryonic stem cells through expression of GFP under the control of Figla promoter. J. Cell Biochem. 2012, 113, 1111–1121. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Ratajczak, J.; Kucia, M. Very Small Embryonic-Like Stem Cells (VSELs) An Update and Future Directions. Circ. Res. 2019, 124, 208–210. [Google Scholar] [CrossRef]
- Lahlil, R.; Scrofani, M.; Barbet, R.; Tancredi, C.; Aries, A.; Hénon, P. VSELs maintain their pluripotency and competence to differentiate after enhanced ex vivo expansion. Stem Cell Rev. Rep. 2018, 14, 510–524. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Zuba-Surma, E.; Wojakowski, W.; Suszynska, M.; Mierzejewska, K.; Liu, R.; Ratajczak, J.; Shin, D.M.; Kucia, M. Very small embryonic-like stem cells (VSELs) represent a real challenge in stem cell biology: Recent pros and cons in the midst of a lively debate. Leukemia 2014, 28, 473–484. [Google Scholar] [CrossRef]
- Bhartiya, D.; Anand, S.; Patel, H.; Parte, S. Making gametes from alternative sources of stem cells: Past, present and future. Reprod. Biol. Endocrinol. 2017, 15, 89. [Google Scholar] [CrossRef] [Green Version]
- Bhartiya, D. Ovarian stem cells are always accompanied by very small embryonic-like stem cells in adult mammalian ovary. J. Ovarian Res. 2015, 8, 70. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Guo, X.; Lui, M.; Chu, P.J.; Yoo, J.; Chang, M.; Yen, Y. Identification of a distinct small cell population from human bone marrow reveals its multipotency in vivo and in vitro. PLoS ONE 2014, 9, e85112. [Google Scholar] [CrossRef] [Green Version]
- Nakatsuka, R.; Iwaki, R.; Matsuoka, Y.; Sumide, K.; Kawamura, H.; Fujioka, T.; Sasaki, Y.; Uemura, Y.; Asano, H.; Kwon, A.H.; et al. Identification and characterization of lineage(-)CD45(-)Sca-1(+) VSEL phenotypic cells residing in adult mouse bone tissue. Stem Cells Dev. 2016, 25, 27–42. [Google Scholar] [CrossRef]
- Halasa, M.; Baskiewicz-Masiuk, M.; Dabkowska, E.; Machalinski, B. An efficient two-step method to purify very small embryonic-like (VSEL) stem cells from umbilical cord blood (UCB). Folia Histochem. Cytobiol. 2008, 46, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Sovalat, H.; Scrofani, M.; Eidenschenk, A.; Hénon, P. Human Very Small Embryonic-Like Stem Cells Are Present in Normal Peripheral Blood of Young, Middle-Aged, and Aged Subjects. Stem Cells Int. 2016, 2016, 7651645. [Google Scholar] [CrossRef] [Green Version]
- Bhartiya, D.; Shaikh, A.; Anand, S.; Patel, H.; Kapoor, S.; Sriraman, K.; Parte, S.; Unni, S. Endogenous, very small embryonic-like stem cells: Critical review, therapeutic potential and a look ahead. Hum. Reprod. Update 2016, 23, 41–76. [Google Scholar] [CrossRef] [Green Version]
- Virant-klun, I. Functional testing of primitive oocyte like cell developed in ovarian surface epithelium cell culture from small VSEL-like stem cell: Can they be fertilized one day? Stem Cell Rev. Rep. 2018, 14, 715–721. [Google Scholar] [CrossRef]
- Lee, H.J.; Selesniemi, K.; Niikura, Y.; Niikura, T.; Klein, R.; Dombkowski, D.M.; Tilly, J.L. Bone marrow transplantation generates immature oocytes and rescues long-term fertility in a preclinical mouse model of chemotherapy-induced premature ovarian failure. J. Clin. Oncol. 2007, 25, 3198–3204. [Google Scholar] [CrossRef]
- Abd-Allah, S.H.; Shalaby, S.M.; Pasha, H.F.; El-Shal, A.S.; Raafat, N.; Shabrawy, S.M.; Awad, H.A.; Amer, M.G.; Gharib, M.A.; El Gendy, E.A.; et al. Mechanistic action of mesenchymal stem cell injection in the treatment of chemically induced ovarian failure in rabbits. Cytotherapy 2013, 15, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Lai, D.; Wang, F.; Yao, X.; Zhang, Q.; Wu, X.; Xiang, C. Human endometrial mesenchymal stem cells restore ovarian function through improving the renewal of germline stem cells in a mouse model of premature ovarian failure. J. Transl. Med. 2015, 13, 155. [Google Scholar] [CrossRef] [Green Version]
- Ding, C.; Zou, Q.; Wang, F.; Wu, H.; Chen, R.; Lv, J.; Ling, M.; Sun, J.; Wang, W.; Li, H.; et al. Human amniotic mesenchymal stem cells improve ovarian function in natural aging through secreting hepatocyte growth factor and epidermal growth factor. Stem Cell Res. Ther. 2018, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Zhong, Y.; Qian, C.; Zou, Q.; Ou, J.; Shi, Y.; Gao, L.; Wang, G.; Liu, Z.; Li, H.; et al. Human umbilical cord mesenchymal stem cells therapy in cyclophosphamide-induced premature ovarian failure rat model. Biomed. Res. Int. 2016, 2016, 2517514. [Google Scholar] [CrossRef] [Green Version]
- Takehara, Y.; Yabuuchi, A.; Ezoe, K.; Kuroda, T.; Yamadera, R.; Sano, C.; Murata, N.; Aida, T.; Nakama, K.; Aono, F.; et al. The restorative effects of adipose-derived mesenchymal stem cells on damaged ovarian function. Lab. Investig. 2013, 93, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Silvestris, E.; Cafforio, P.; Felici, C.; Cormio, G.; D’Oronzo, S. Ddx4+ Oogonial Stem Cells in Postmenopausal Women’s Ovaries: A Controversial, Undefined Role. Cells 2019, 8, 650. [Google Scholar] [CrossRef] [Green Version]
- Edessy, M.; Hosni, H.N.; Shady, Y.; Waf, Y.; Bakr, S.; Kamel, M. Autologous stem cells therapy, The first baby of idiopathic premature ovarian failure. Acta Med. Int. 2016, 3, 19–23. [Google Scholar] [CrossRef]
- Hashimoto, K.; Noguchi, M.; Nakatsuji, N. Mouse offspring derived from fetal ovaries or reaggregates which were cultured and transplanted into adult females. Dev. Growth Differ. 1992, 34, 233–238. [Google Scholar] [CrossRef]
- Matoba, S.; Ogura, A. Generation of functional oocytes and spermatids from fetal primordial germ cells after ectopic transplantation in adult mice. Biol. Reprod. 2011, 84, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Ogushi, S.; Kurimoto, K.; Shimamoto, S.; Ohta, H.; Saitou, M. Offspring from Oocytes Derived from in Vitro Primordial Germ Cell–like Cells in Mice. Science 2012, 338, 971–975. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Xiong, J.; Ma, L.; Lu, Z.; Qin, X.; Luo, A.; Zhang, J.; Xie, H.; Shen, W.; Wang, S. Enrichment of female germline stem cells from mouse ovaries using the differential adhesion method. Cell Physiol. Biochem. 2018, 46, 2114–2126. [Google Scholar] [CrossRef] [Green Version]



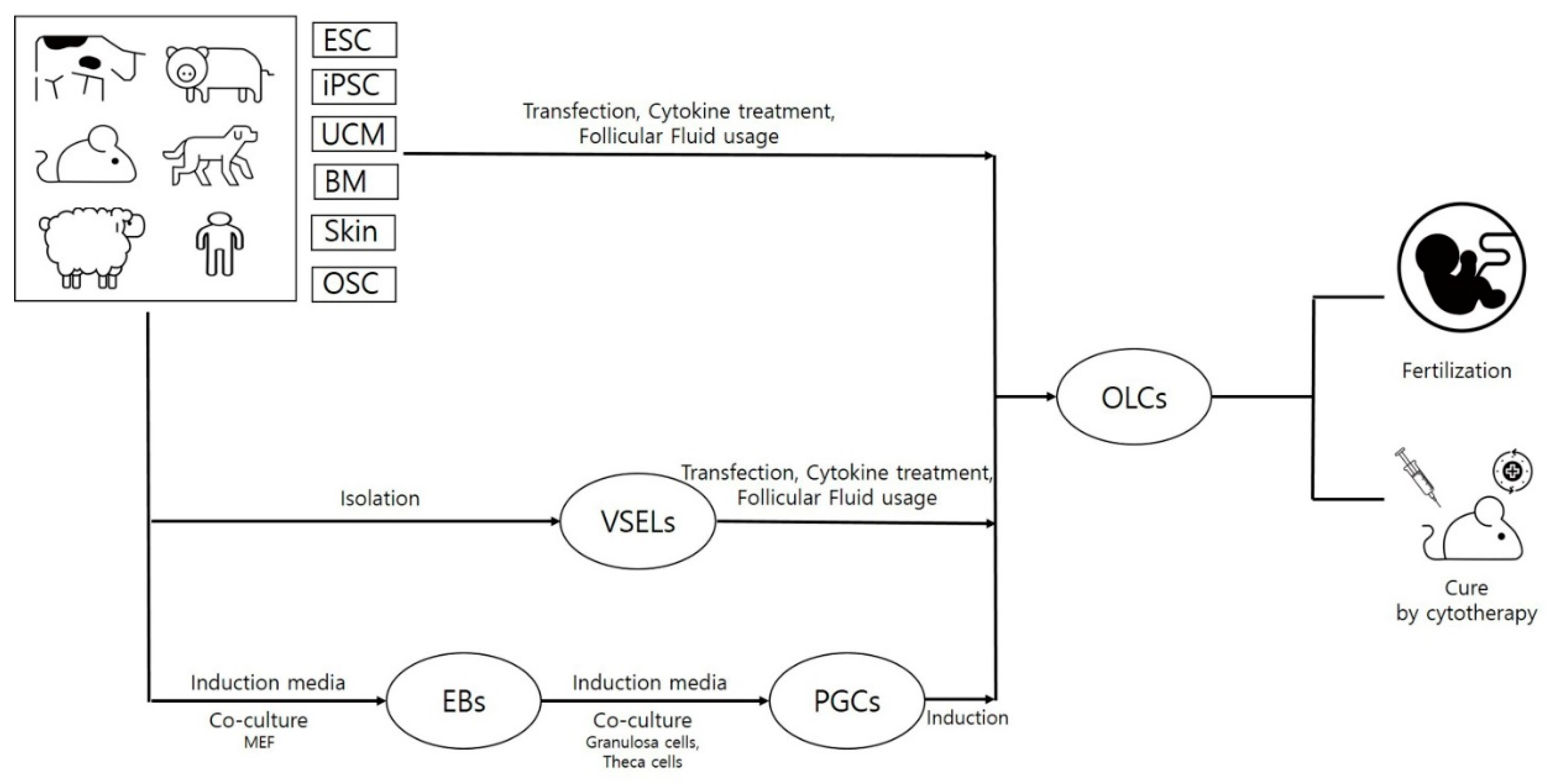{kind=link}
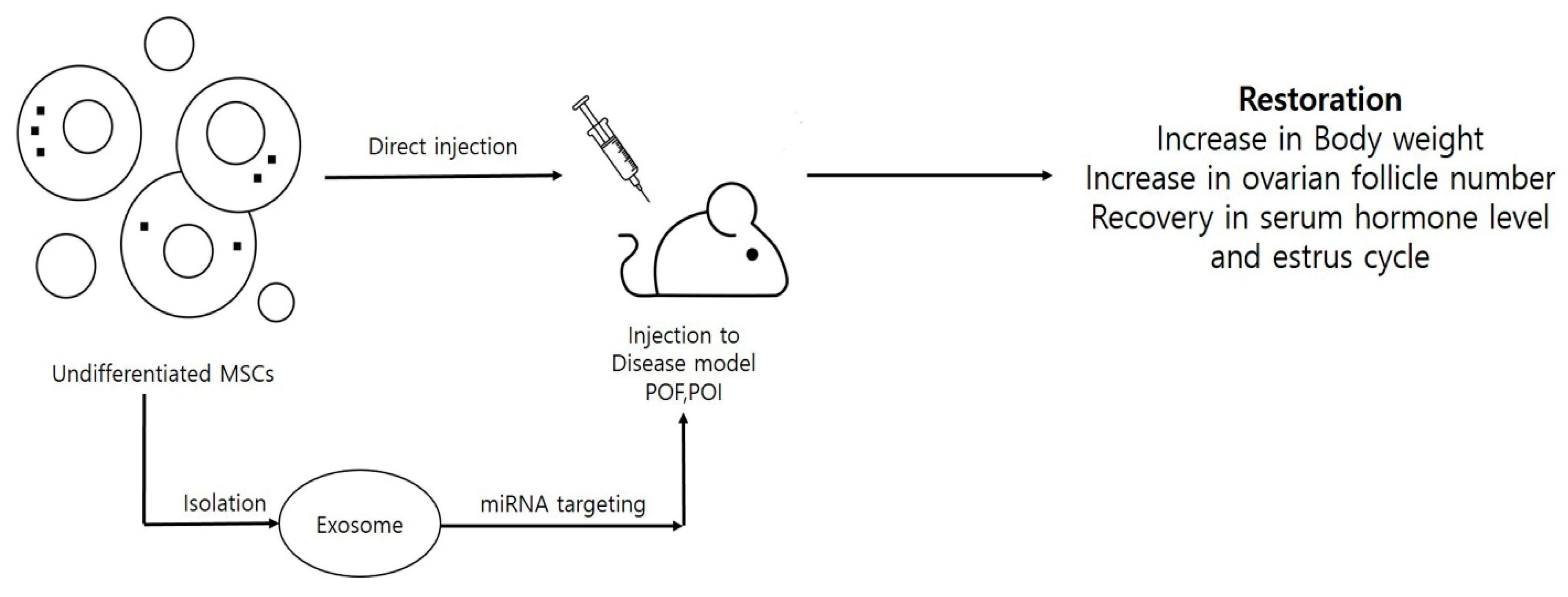{kind=link}
| Category | Name of Gene | Function |
|---|---|---|
| Pluripotency marker | Oct4 | Regulates pluripotency and also acts as germ cell marker. |
| Nanog | Maintains pluripotency. | |
| Sox2 | Maintains pluripotency. | |
| Rex | Maintains pluripotent state. | |
| Ssea4 | Regulates the pre-implantation development and pluripotency of human embryonic stem cells. | |
| Oocyte-specific marker | Gdf9 | Oocyte maturation marker. |
| C-Mos | Oocyte maturation factor Mos. | |
| Zpc | Contributes to the structural integrity of oocyte and its coat. | |
| Stella | Involved in Chromosomal organization or RNA processing. | |
| Germ cell-specific marker | Dazl | Primordial germ cell migration, germ stem cell proliferation differentiation, Suppresses pluripotency markers expression. |
| Vasa | Germ cell determination and oocyte development. | |
| Blimp1 | Master regulator of the foundation of the germ cell lineage and primordial germ cell formation. | |
| Fragilis | Thought to be involved in the initiation of homeobox genes repression in the early germ cell precursors, and pluripotency maintenance. | |
| Meiosis-specific marker | Dmc1 | Essential for meiotic recombination and meiosis regulation. |
| Scp (1,2,3) | Synaptonemal complex protein (1,2,3) are required for normal centromere pairing, chromosomal synapsis during male and female germ cell development, important meiosis marker. | |
| Rec8 | Meiotic recombination protein which participate in meiotic process (meiotic cohesion complex, sister chromatid cohesion and homologous chromosome recombination). | |
| Stra8 | Regulation of meiotic initiation in both spermatogenesis and oogenesis. | |
| Primordial germ cell marker | Prdm1/Blimp-1 | Repress beta-interferon gene expression, plays critical role in primordial germ cells and germ cell formation. |
| Prdm14 | Maintain pluripotency and suppress the expression of differentiation marker genes, helps in primordial germ cell formation. | |
| Dppa3 | Developmental pluripotency-associated protein 3 participate in epigenetic chromatin reprogramming, prevent 5mC to conversion (DNA methylation protection), the condensation of chromatin during oocytogenesis. | |
| Dnd1 | Survival of primordial germ cells, germ cell tumor suppression in mice, apoptosis suppression. | |
| Nanos3 | Maintain undifferentiated germ cell state, participate in human germ cell development, apoptosis suppression. | |
| Tcfap2c/Ap2-γ | Placenta development, primordial germ cell maintenance. | |
| Granulosa cell marker | Foxl2 | Important for ovarian development and function, granulosa cell differentiation marker, support pre-ovulatory follicles growth. |
| Amh | TGF-β family member, help in the proliferation of mitotically active germ cell, folliculogenesis regulation, molecule biomarker for ovarian reserve and menopause timing. | |
| FshR | Expressed by granulosa cell, necessary follicular development, controls the growth and maturation of spermatogenesis and follicle formation. | |
| CYP19A1 | Helps in estrogen biosynthesis. | |
| Theca cell marker | LHR | Helps in follicular maturation, ovulation and luteal function. |
| Stem Cell Source | Co-Culture | FF Concentration | Final Product | Reference | |
|---|---|---|---|---|---|
| Human | Ovarian stem cell | MEF layer | 0 | OLCs | [57] |
| Wharton’s jelly | Placental cells | 0 | OLCs | [12] | |
| Theca cell | GCs | 5% human | OLCs | [5] | |
| Menstrual blood-derived endometrial MSC | GCs | 0 | OLCs | [15] | |
| Induced Pluripotent Stem Cells | Fetal gonadal stromal cells | 0 | PGCs | [82] | |
| Embryonic stem cell | Fetal gonadal stromal cells | 0 | PGCs | [82] | |
| Embryonic stem cell | MEF layer | 0 | PGCs | [73] | |
| Porcine | Female germline stem cell | GCs | 5% porcine | OLCs | [83] |
| Female germline stem cell | MEF layer | 5% porcine | OLCs | [83] | |
| Mouse | Skin-derived stem cell | MEF layer | 0 | PGCs | [21] |
| Skin-derived stem cell | MEF layer | 0 | PGCs | [37] | |
| Skin-derived stem cell | Newborn mouse ovarian cells | 0 | OLCs | [18] | |
| Embryonic stem cell | Ovarian somatic cells | 0 | OLCs | [20] | |
| Embryonic stem cell | GCs | 0 | OLCs | [20] | |
| Embryonic stem cell | GCs | 0 | Germ-like cells | [84] | |
| Embryonic stem cell | New born mouse granulosa cells | 0 | OLCs | [17] | |
| Embryonic stem cell | OP9 stromal cells | 0 | PGC-like cells | [85] | |
| Ovarian stem cell | GCs | 0 | OLCs | [22] | |
| Female germline stem cell | GCs | 0 | OLCs | [86] | |
| Embryonic stem cell | MEF layer | 0 | PGCs | [41] | |
| Cell Source | Transfection | Final Product | Implication | Knockdown | Reference | |
|---|---|---|---|---|---|---|
| Human | ASCs | BMP15 | OLCs | BMP15 activation, expressed oocyte-specific markers (ZP1, ZP2, ZP3, c-kit), detection of VASA protein during later stage of induction. | x | [94] |
| ESCs | NANOS3 | Germ cells | Over expressed protein levels PRKSCH. No expression of OLFM2, delayed in vitro differentiation. | x | [95] | |
| ESCs | DAZL | Germ cells | Up-regulated DAZL, IFITM3, BMP7, HIP1R, ISYNA1, JAG2, PIDD1, RPRM. Down-regulated CXCL5, GABRP, FAM110C, LCP1. | x | [95] | |
| ESCs | DAZL, BOULE | Follicle-like cells | Increased DAZL and BOULE expression. down-regulation of pluripotency markers (OCT4, NANOG, PRDM14), up-regulated luciferase activity. Promoted entry into meiosis. | shRNA (DAZL) | [16] | |
| ASCs | BMP15 | OLCs, GC | Differentiation into OLC, GC, with increased size, BMP15 activation and parthenogenetic embryo-like structures formation. | x | [11] | |
| UCMSCs | FIGLα | OLCs | Over expressed Figlα, stimulated germ cell-specific markers (DAZL, GDF9, ZP2) | x | [7] | |
| ESCs | PRDM1 | PGCs | Suppressed Sox2 expression, promoted germline differentiation. | shRNA (PRDM1) | [74] | |
| Mouse | ESCs | FIGLα | Female germ-like cells | Differentiation into germ-like cells, high expression of germ cell markers (VASA), meiotic-specific genes (STRA8, SCP3), and oocyte markers (GDF9, ZP3, FIGLα). | x | [96] |
| ESCs | DAZL | Germ cells | Overexpression of DAZL. Suppression of Nanog. Induced germ cell nuclear antigen expression. | siRNA (DAZL) | [42] | |
| ESCs | PRDM1 | Germ cells | Suppressed SOX2 expression and neural fate, increased expression of markers like PRDM1, OCT4, NANOS3, VASA and SCP3, enhances BMP4 & WNT3A induced germline differentiation. | x | [73] | |
| Embryonal carcinoma cells | BLIMP1, PRDM14, AP2γ | PGCs | Directed EpiLCs formation, up-regulated PGC-specification/development genes BLIMP1, PRDM14, TFAP2C, NANOS3, STELLA, down-regulated epigenetic modifier (DNMT3a, DNMT3b). | x | [71] | |
| Porcine | Ovarian cortex | OCT4 | OLCs | Oct4 overexpression. Enhanced AP activity and cell proliferation. Higher oogenesis potential. | x | [29] |
| Skin-derived stem cells | DAZL | PGCs Or OLCs | Induced differentiation. Increased expression of germ cell markers (Oct4, Stella, Vasa) and meiosis markers (Scp3, Dmc1, Rec8, Stra8). | siRNA (DAZL) | [28] | |
| Goat | Ear Pinnae cells | OCT4, SOX2, NANOG | iPSCs OLCs | Cell reprogramming, increased potency. | x | [32] |
| Stem Cell Source | Target Disease Model | Implication | Reference | |
|---|---|---|---|---|
| Bone marrow mesenchymal stem cell | Mouse | POF | Formed primordial follicles, increased E2 level, decreased FSH level, attained pregnant after natural breeding. | [48] |
| Rabbit | POF | Increased E2 and VEGF levels, decrease FSH level, increased follicle number. | [109] | |
| Human | POF | After transplantation menstruation improved, one case delivered healthy full baby, increased ovarian reserve score. | [115] | |
| Umbilical cord mesenchymal stem cell | Human | POF | Decreased serum FSH level, recovery in serum E2 and AMH levels, increase in secondary follicles, reduced ovarian cell’s apoptosis. | [112] |
| Human | POI | Increased body weight, estrous cycle recovery, increase in ovarian follicles, increase in serum E2 level, decrease in serum FSH level, induced angiogenesis and cytokine expression in the ovary. | [56] | |
| Human | POI | Increase in ovarian weight, increased weight in E2-dependent organs, increased follicular number, decrease in serum FSH level, increase in AMH level, increased AMH expression, elevation in pregnancy rate. | [54] | |
| Menstrual blood stem cell | Human | POF | Increased expression of ovarian markers [AMH, inhibin α/β, and follicle-stimulating hormone receptor (FSHR)], increased Ki67 expression, increased ovarian weight, increased plasma E2 level and increased number of normal follicles. | [9] |
| Human | POI | Increased ovarian weight and total follicle number, estrus cycle restoration, decreased serum FSH level, increased serum E2, AMH level, reduced apoptosis, increase in the expression of AMH, DDX4 and VEGFA. | [51] | |
| Amniotic MSC | Human | Natural ovarian aging | Increase in all stage follicle number, recovery in E2 and AMH levels, decreased FSH levels, promoted the proliferation rate, high expression of ovarian and granular cell markers (AMH, FSHR, FOXL2, CYP19A1). | [111] |
| Human | POF | Recovered estrus cycle, increase in estrogen level, decrease in FSH levels, increase in ovarian index, fertility rate and population of follicles at different stages. | [53] | |
| Chorionic MSC | Human | POF | Decreased serum FSH level, increased serum E2 level and number of follicles, restored estrous cycle, increased oocyte population. | [50] |
| Adipose-derived MSC | Human | Damaged ovarian | Induced angiogenesis, increase in ovarian follicles, corpus luteal and also in number of litters. | [113] |
| Endometrial MSC | Human | POF | Increased body weight, restored estrus cycle, re- established fertility, MSC infiltration to damaged ovarian tissue and differentiated into granulosa cell, improved renewal of germline stem cells. | [110] |
| Skin-derived MSC | Mouse | POF | Increased body weight, increased weight of reproductive organs, restored fertility, reduced pro-inflammatory cytokines (TNF-a, TGF-b, IL-8, IL-6, IL-1b, and IFN γ), increased expression of genes Nobox, Nanos3, and Lhx8 genes in the transplanted ovaries. | [46] |
| Pathological Condition | Cell Therapy | Estimated Enrollment | Intervention/ Treatment | Cliniclatrials.Gov Identifier | Outcome Measures |
|---|---|---|---|---|---|
| Women infertility with thin endometrium or endometrial scarring. | UCMSCs loaded in collagen scaffold | 50 participants | UCMSC-based therapy | NCT03592849 | Evaluation of endometrial thickness, pregnancy rate, living intrauterine fetus, live birth rate, endometrial blood flow, menstrual blood volume and adversities such as infection, allergies, abdominal pain post-operation. |
| Women with POI | AECs | 20 participants | Minimally invasive implantation AECs used with ultrasound guidance | NCT03207412 | Evaluation of serum FSH, primordial egg follicles number, menstrual cycle and menstrual period, LH, AMH and E2, ovarian volume. |
| Repeated IVF failure women with atrophic endometrium | Autologous BMSCs | 46 participants | Transplantation of BMSCs | NCT03166189 | Evaluation of endometrial receptivity, endometrial thickness, pregnancy rate, adversities such as side effects, abdominal discomfort and patient’s tolerance. |
| Women with POF | Autologous BMSCs | 33 participants | Transplantation of BMSCs | NCT02696889 | Evaluation of FSH, AMH, E2 level, menstruation resumption, pregnancy achievement. |
| Women with POF | Autologous AMSCs | 9 participants | Intra-ovarian injection of AMSCs | NCT02603744 | Evaluation of FSH, AMH, antral follicle number and volume, menstruation recurrence rate and pregnancy rate. |
| Women infertility with intrauterine adhesions | UCMSCs loaded in collagen scaffold | 26 participants | UCMSC-based therapy | NCT02313415 | Evaluation of live birth rate, intrauterine adhesion reduction, change of endometrial thickness, menstrual blood volume. |
| Women infertility with intrauterine adhesions and endometrial dysplasia | Autologous BMSCs loaded in collagen scaffold | 30 participants | BMSC-based therapy | NCT02204358 | Evaluation of intrauterine scar reduction, endometrial thickness, menstrual blood volume and pregnancy rate. |
| Women with POF | Autologous BMSCs | 60 participants | Transplantation of BMSCs | NCT02062931 | Evaluation of FSH, AMH, E2 level, hormonal, clinical, ultra sound, menopausal symptoms and pregnancy rate. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bharti, D.; Jang, S.-J.; Lee, S.-Y.; Lee, S.-L.; Rho, G.-J. In Vitro Generation of Oocyte Like Cells and Their In Vivo Efficacy: How Far We have been Succeeded. Cells 2020, 9, 557. https://doi.org/10.3390/cells9030557
Bharti D, Jang S-J, Lee S-Y, Lee S-L, Rho G-J. In Vitro Generation of Oocyte Like Cells and Their In Vivo Efficacy: How Far We have been Succeeded. Cells. 2020; 9(3):557. https://doi.org/10.3390/cells9030557
Chicago/Turabian StyleBharti, Dinesh, Si-Jung Jang, Sang-Yun Lee, Sung-Lim Lee, and Gyu-Jin Rho. 2020. "In Vitro Generation of Oocyte Like Cells and Their In Vivo Efficacy: How Far We have been Succeeded" Cells 9, no. 3: 557. https://doi.org/10.3390/cells9030557
APA StyleBharti, D., Jang, S. -J., Lee, S. -Y., Lee, S. -L., & Rho, G. -J. (2020). In Vitro Generation of Oocyte Like Cells and Their In Vivo Efficacy: How Far We have been Succeeded. Cells, 9(3), 557. https://doi.org/10.3390/cells9030557