Response to Stimulations Inducing Circadian Rhythm in Human Induced Pluripotent Stem Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Stimulation to Synchronize Circadian Rhythm
2.3. Quantitative Real-Time PCR
2.4. Western Blot Analysis
2.5. Flow Cytometry
2.6. Microarray
2.7. Statistical Analyses
3. Results
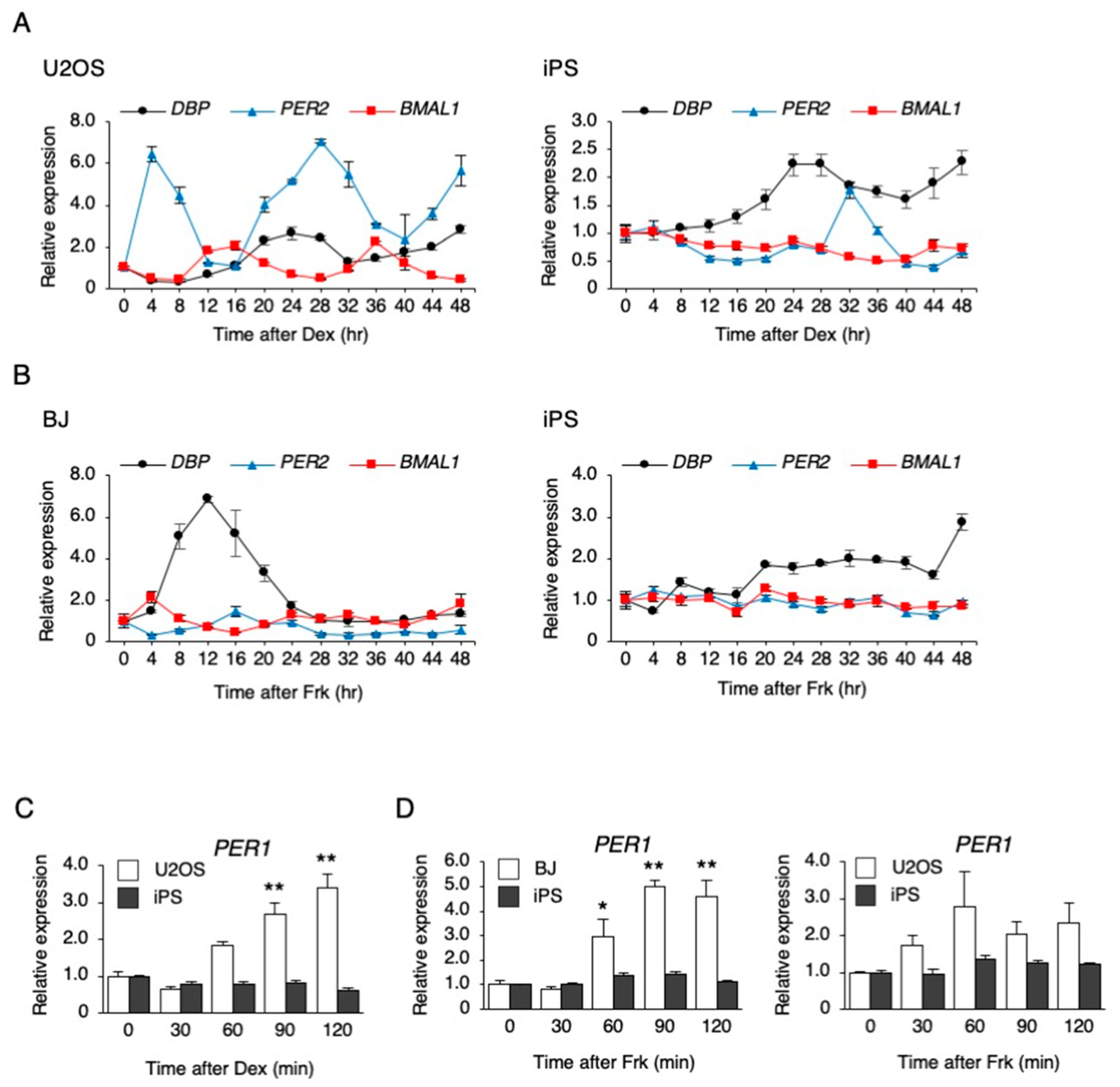
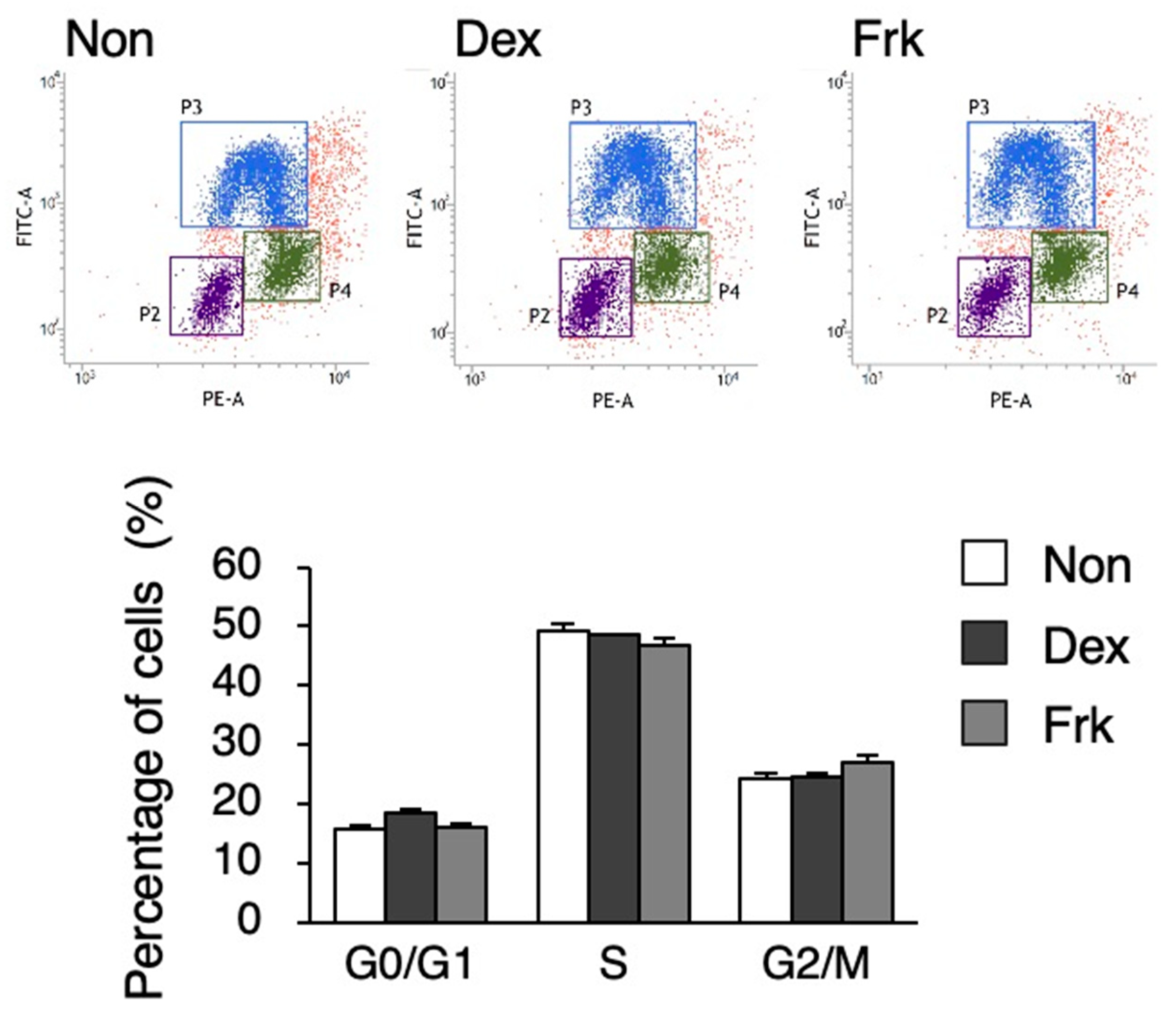
3.1. Dexamethasone (Dex) and Forskolin (Frk) Stimulation did not Elicit a Circadian Rhythm in hiPSCs
3.2. Characteristics of Clock Gene Expression in hiPSCs
3.3. Circadian-Like Rhythms Occurred under a Circadian Temperature in hiPSCs
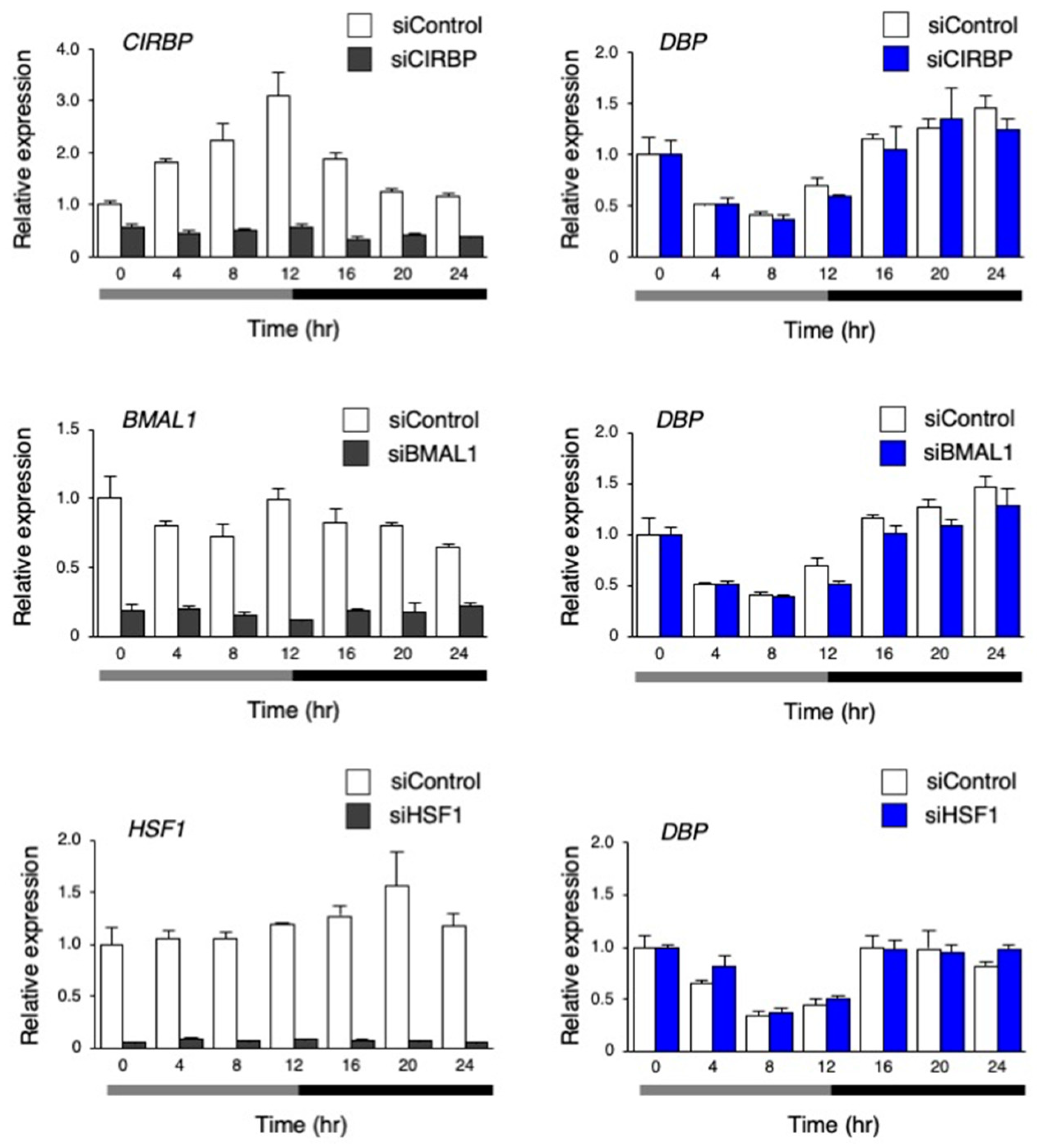
3.4. The Circadian-Like Rhythm of DBP Expression was Not Abolished by the Inhibition of CIRBP, BMAL1, and HSF1
3.5. Hypoxia Signaling is Involved in the Circadian-Like Rhythm of Gene Expression by the Temperature Rhythm
3.6. The Circadian-Like Rhythm of DBP was Abolished by HIF-1α Inhibition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Inoue, H.; Nagata, N.; Kurokawa, H.; Yamanaka, S. iPS cells: A game changer for future medicine. EMBO J. 2014, 33, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, P.; Takahashi, K.; Saito, M.; Yoshida, Y.; Okita, K.; Watanabe, A.; Inoue, H.; Yamashita, J.K.; Todani, M.; Nakagawa, M.; et al. Induced Pluripotent Stem Cells and Their Use in Human Models of Disease and Development. Physiol. Rev. 2019, 99, 79–114. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Cheng, A.W.; Saha, K.; Kim, J.; Lengner, C.J.; Soldner, F.; Cassady, J.P.; Muffat, J.; Carey, B.W.; Jaenisch, R. Human embryonic stem cells with biological and epigenetic characteristics similar to those of mouse ESCs. Proc. Natl. Acad. Sci. USA 2010, 107, 9222–9227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duggal, G.; Warrier, S.; Ghimire, S.; Broekaert, D.; Van der Jeught, M.; Lierman, S.; Deroo, T.; Peelman, L.; Van Soom, A.; Cornelissen, R.; et al. Alternative Routes to Induce Naïve Pluripotency in Human Embryonic Stem Cells. Stem Cells 2015, 33, 2686–2698. [Google Scholar] [CrossRef]
- Liu, Y.; Goldberg, A.J.; Dennis, J.E.; Gronowicz, G.A.; Kuhn, L.T. One-step derivation of mesenchymal stem cell (MSC)-like cells from human pluripotent stem cells on a fibrillar collagen coating. PLoS ONE 2012, 7, e33225. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.; Luo, Y.; Chen, M.; Wang, G.; Ding, M.; Petersen, C.C.; Kang, R.; Dagnaes-Hansen, F.; Zeng, Y.; Lv, N.; et al. A simple method for deriving functional MSCs and applied for osteogenesis in 3D scaffolds. Sci. Rep. 2013, 3, 2243. [Google Scholar] [CrossRef]
- Lu, C.; Yang, Y.; Zhao, R.; Hua, B.; Xu, C.; Yan, Z.; Sun, N.; Qian, R. Role of circadian gene Clock during differentiation of mouse pluripotent stem cells. Protein. Cell. 2016, 7, 820–832. [Google Scholar] [CrossRef] [Green Version]
- Pauklin, S.; Vallier, L. The cell-cycle state of stem cells determines cell fate propensity. Cell 2013, 155, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Chetty, S.; Pagliuca, F.W.; Honore, C.; Kweudjeu, A.; Rezania, A.; Melton, D.A. A simple tool to improve pluripotent stem cell differentiation. Nat. Methods 2013, 10, 553–556. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [Green Version]
- Preitner, N.; Damiola, F.; Lopez-Molina, L.; Zakany, J.; Duboule, D.; Albrecht, U.; Schibler, U. The orphan nuclear receptor REV- ERBα controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 2002, 110, 251–260. [Google Scholar] [CrossRef]
- Sato, T.; Panda, S.; Miraglia, L.J.; Reyes, T.M.; Rudic, R.D.; McNamara, P.; Naik, K.A.; FitzGerald, G.A.; Kay, S.A.; Hogenesch, J.B. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron 2004, 43, 527–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Fang, B.; Emmett, M.J.; Damle, M.; Sun, Z.; Feng, D.; Armour, S.M.; Remsberg, J.R.; Jager, J.; Soccio, R.E.; et al. Discrete functions of nuclear receptor Rev-erbα couple metabolism to the clock. Science 2015, 348, 1488–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsalobre, A.; Damiola, F.; Schibler, U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell 1998, 93, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, J.S.; Hong, H.K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Yagita, K.; Horie, K.; Koinuma, S.; Nakamura, W.; Yamanaka, I.; Urasaki, A.; Shigeyoshi, Y.; Kawakami, K.; Shimada, S.; Takeda, J.; et al. Development of the circadian oscillator during differentiation of mouse embryonic stem cells in vitro. Proc. Natl. Acad. Sci. USA 2010, 107, 3846–3851. [Google Scholar] [CrossRef] [Green Version]
- Paulose, J.K.; Rucker, E.B., III; Cassone, V.M. Toward the beginning of time: Circadian rhythms in metabolism precede rhythms in clock gene expression in mouse embryonic stem cells. PLoS ONE 2012, 7, e49555. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, E.; Moriggi, E.; Bauer, C.; Dibner, C.; Brown, S.A. The circadian clock starts ticking at a developmentally early stage. J. Biol. Rhythms 2010, 25, 442–449. [Google Scholar] [CrossRef] [Green Version]
- Umemura, Y.; Koike, N.; Ohashi, M.; Tsuchiya, Y.; Meng, Q.J.; Minami, Y.; Hara, M.; Hisatomi, M.; Yagita, K. Involvement of posttranscriptional regulation of Clock in the emergence of circadian clock oscillation during mouse development. Proc. Natl. Acad. Sci. USA 2017, 114, E7479–E7488. [Google Scholar] [CrossRef] [Green Version]
- Dierickx, P.; Vermunt, M.W.; Muraro, M.J.; Creyghton, M.P.; Doevendans, P.A.; van Oudenaarden, A.; Geijsen, N.; Van Laake, L.W. Circadian networks in human embryonic stem cell-derived cardiomyocytes. EMBO Rep. 2017, 18, 1199–1212. [Google Scholar] [CrossRef]
- Umemura, Y.; Maki, I.; Tsuchiya, Y.; Koike, N.; Yagita, K. Human Circadian Molecular Oscillation Development Using Induced Pluripotent Stem Cells. J. Biol. Rhythms. 2019, 34, 525–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagita, K.; Okamura, H. Forskolin induces circadian gene expression of rPer1, rPer2 and dbp in mammalian rat-1 fibroblasts. FEBS Lett. 2000, 465, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.A.; Zumbrunn, G.; Fleury-Olela, F.; Preitner, N.; Schibler, U. Rhythms of mammalian body temperature can sustain peripheral circadian clocks. Curr. Biol. 2002, 12, 1574–1583. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molcan, L. Time distributed data analysis by Cosinor. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Balsalobre, A.; Marcacci, L.; Schibler, U. Multiple signaling pathways elicit circadian gene expression in cultured Rat-1 fibroblasts. Curr. Biol. 2000, 10, 1291–1294. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Lee, J.; Kwon, I.; Nakajima, Y.; Ohmiya, Y.; Son, G.H.; Lee, K.H.; Kim, K. Coactivation of the CLOCK-BMAL1 complex by CBP mediates resetting of the circadian clock. J. Cell. Sci. 2010, 123, 3547–3557. [Google Scholar] [CrossRef] [Green Version]
- Aschoff, J. Circadian control of body temperature. J. Therm. Biol. 1983, 8, 143–147. [Google Scholar] [CrossRef]
- Saini, C.; Morf, J.; Stratmann, M.; Gos, P.; Schibler, U. Simulated body temperature rhythms reveal the phase-shifting behavior and plasticity of mammalian circadian oscillators. Genes Dev. 2012, 26, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Morf, J.; Rey, G.; Schneider, K.; Stratmann, M.; Fujita, J.; Naef, F.; Schibler, U. Cold-inducible RNA-binding protein modulates circadian gene expression posttranscriptionally. Science 2012, 338, 379–383. [Google Scholar] [CrossRef]
- Kaelin, W.G. Proline hydroxylation and gene expression. Annu. Rev. Biochem. 2005, 74, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Balsalobre, A.; Brown, S.A.; Marcacci, L.; Tronche, F.; Kellendonk, C.; Reichardt, H.M.; Schütz, G.; Schibler, U. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 2000, 289, 2344–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, A.Y.; Bernal, T.U.; Pillsbury, M.L.; Yamamoto, K.R.; Feldman, B.J. Glucocorticoid regulation of the circadian clock modulates glucose homeostasis. Proc. Natl. Acad. Sci. USA 2009, 106, 17582–17587. [Google Scholar] [CrossRef] [Green Version]
- Sassone-Corsi, P.; Visvader, J.; Ferland, L.; Mellon, P.L.; Verma, I.M. Induction of proto-oncogene fos transcription through the adenylate cyclase pathway: Characterization of a cAMP-responsive element. Genes Dev. 1988, 2, 1529–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, G.A.; Montminy, M.R. Cyclic AMP stimulates somatostatin gene transcription by phosphorylation of CREB at serine 133. Cell 1989, 59, 675–680. [Google Scholar] [CrossRef]
- Holcik, M.; Sonenberg, N. Translational control in stress and apoptosis. Nat. Rev. Mol. Cell. Biol. 2005, 6, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Meides, A.; Zhang, D.P.; Finley, D. A ubiquitin stress response induces altered proteasome composition. Cell 2007, 129, 747–759. [Google Scholar] [CrossRef] [Green Version]
- Masson, N.; Ratcliffe, P.J. HIF prolyl and asparaginyl hydroxylases in the biological response to intracellular O(2) levels. J. Cell. Sci. 2003, 116, 3041–3049. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Regulation of mammalian O2 homeostasis by hypoxia-inducible factor 1. Annu. Rev. Cell. Dev. Biol. 1999, 15, 551–578. [Google Scholar] [CrossRef]
- Epstein, A.C.; Gleadle, J.M.; McNeill, L.A.; Hewitson, K.S.; O’Rourke, J.; Mole, D.R.; Mukherji, M.; Metzen, E.; Wilson, M.I.; Dhanda, A.; et al. Elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell 2001, 107, 43–54. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene | Forward Primer | Reverse Primer |
|---|---|---|---|
| Human | 18S | GAGGATGAGGTGGAACGTGT | GGACCTGGCTGTATTTTCCA |
| GAPDH | TGTCAAGCTCATTTCCTGGTA | CACAGGGTACTTTATTGATGG | |
| DBP | CCAATCATGAAGAAGGCAAGAAA | GGCTGCCTCGTTGTTCTTGT | |
| PER2 | GCTGGCCATCCACAAAAAGA | GCGAAACCGAATGGGAGAAT | |
| BMAL1 | GAGAAGGTGGCCCAAAGAGG | GGAGGCGTACTCGTGATGTT | |
| NR3C1 | AGCAGTGGAAGGTAGACAGC | CCTGTAGTGGCCTGCTGAAT | |
| CREB1 | TTCTCCGGAACACAGATTTCA | AATCCTTGGCACTCCTGGTG | |
| CREB3 | AGAGTGAGAGCTGTAGAAAAGAGG | AATCTTCCTCCGCACACGTT | |
| PRKACA | AAGAAGGGCAGCGAGCAG | CTGTGTTCTGAGCGGGACTT | |
| PRKACB | AGAGAACCACCTTGTAACCAGTA | TGGCTTTGGCTAGAAACTCTT | |
| CREBBP | TGGCTGAGAACTTGCTGGAC | TGGAAGCAGCATCTGGAACA | |
| EP300 | ACCAGGAATGACTTCTAGTTTGA | TACGAGGCCCATAGCCCATA | |
| CRTC1 | CAGCCGAGGCCAGTACTATG | AAGGGGGTCAGAGAGACAGG | |
| CRTC2 | GGTGATGATGGACATCGGCT | CCGAGTGCTCCGAGATGAAT | |
| CRTC3 | TGTGGGTTTTGACCAGCAGT | TCTTTGAACAGGCTGGTGCT | |
| CLOCK | ACGACGAGAACTTGGCATTG | TCCGAGAAGAGGCAGAAGG | |
| PER1 | CCCAGCACCACTAAGCGTAAA | TGCTGACGGCGGATCTTT | |
| PER3 | GCCTTACAAGCTGGTTTGCAA | CTGTGTCTATGGACCGTCCATTT | |
| CRY1 | ACTCCCGTCTGTTTGTGATTCG | GCTGCGTCTCGTTCCTTTCC | |
| CRY2 | TCTTCCAGCAGTTCTTCC | GTAGTCCACACCAATGATG | |
| CIRBP | CAGATCTCTGAAGTGGTGGT | CCTGCCTGGTCTACTCGGAT | |
| HSF1 | TGCAGCTGATGAAGGGGAAG | AGGATCCGGTTTGACTGCAC | |
| DDIT4 | GGTTCGCACACCCATTCAAG | CAGGGCGTTTGCTGATGAAC | |
| TFRC | GGACGCGCTAGTGTTCTTCT | CATCTACTTGCCGAGCCAGG | |
| VEGFA | CCCTGATGAGATCGAGTACAT | CGGCTTGTCACATCTGCAAGT | |
| EGLN1 | AGCCCAGTTTGCTGACATTG | TCGTGCTCTCTCATCTGCATC | |
| HIF1A | CCTCTGGACTTGCCTTTCCT | TGGCTGCATCTCGAGACTTT | |
| Mouse | 18s | GAGGATGAGGTGGAGCGAGT | GAACCTGGCTGTACTTCCCA |
| Dbp | CGTGGAGGTGCTTAATGACCTTT | CATGGCCTGGAATGCTTGA | |
| Per2 | ATGCTCGCCATCCACAAGA | GCGGAATCGAATGGGAGAAT | |
| Bmal1 | CCAAGAAAGTATGGACACAGACAAA | GCATTCTTGATCCTTCCTTGGT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaneko, H.; Kaitsuka, T.; Tomizawa, K. Response to Stimulations Inducing Circadian Rhythm in Human Induced Pluripotent Stem Cells. Cells 2020, 9, 620. https://doi.org/10.3390/cells9030620
Kaneko H, Kaitsuka T, Tomizawa K. Response to Stimulations Inducing Circadian Rhythm in Human Induced Pluripotent Stem Cells. Cells. 2020; 9(3):620. https://doi.org/10.3390/cells9030620
Chicago/Turabian StyleKaneko, Hitomi, Taku Kaitsuka, and Kazuhito Tomizawa. 2020. "Response to Stimulations Inducing Circadian Rhythm in Human Induced Pluripotent Stem Cells" Cells 9, no. 3: 620. https://doi.org/10.3390/cells9030620
APA StyleKaneko, H., Kaitsuka, T., & Tomizawa, K. (2020). Response to Stimulations Inducing Circadian Rhythm in Human Induced Pluripotent Stem Cells. Cells, 9(3), 620. https://doi.org/10.3390/cells9030620