Innate Cytokine Induced Early Release of IFNγ and CC Chemokines from Hypoxic Human NK Cells Is Independent of Glucose


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Isolation and Culture
2.2. Cell Treatment and Co-Culture
2.3. Extracellular Flux Assay
2.4. Flow Cytometry Analysis
2.5. Western Blotting
2.6. Bead Array Analysis
2.7. Statistical Analysis
3. Results
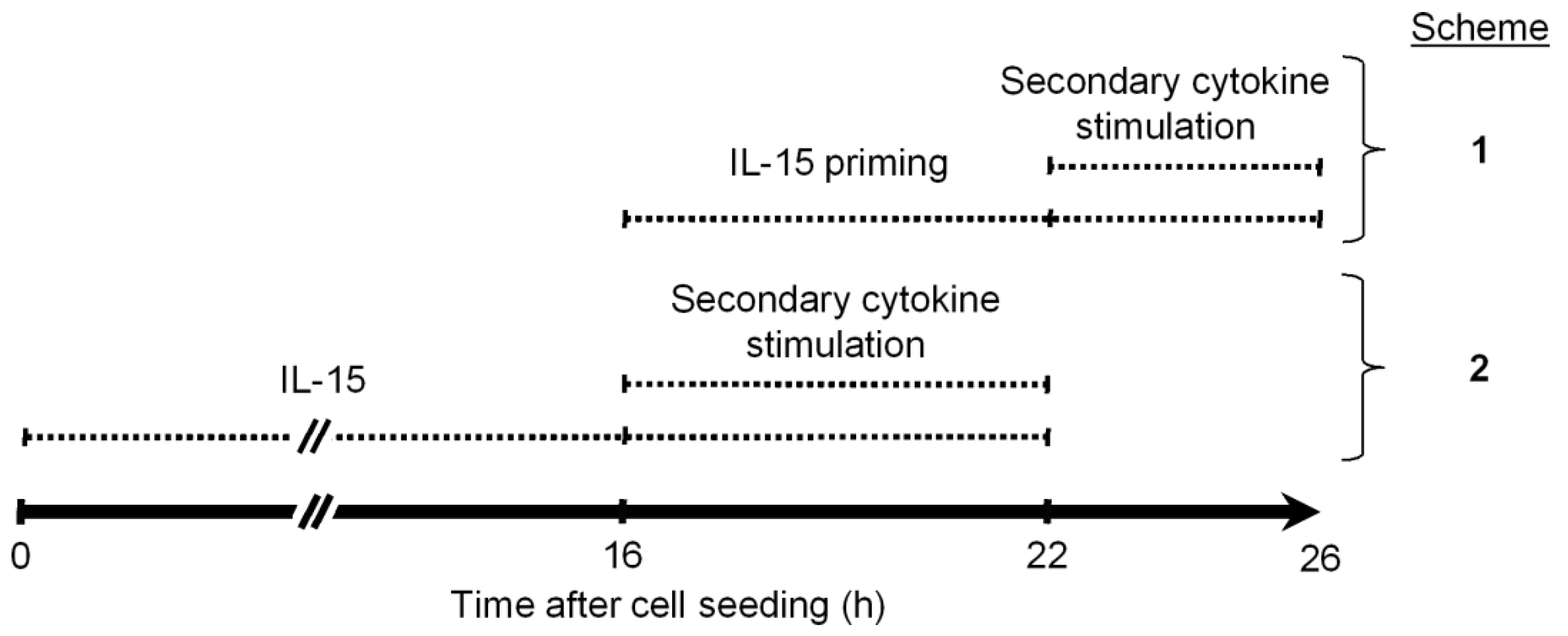
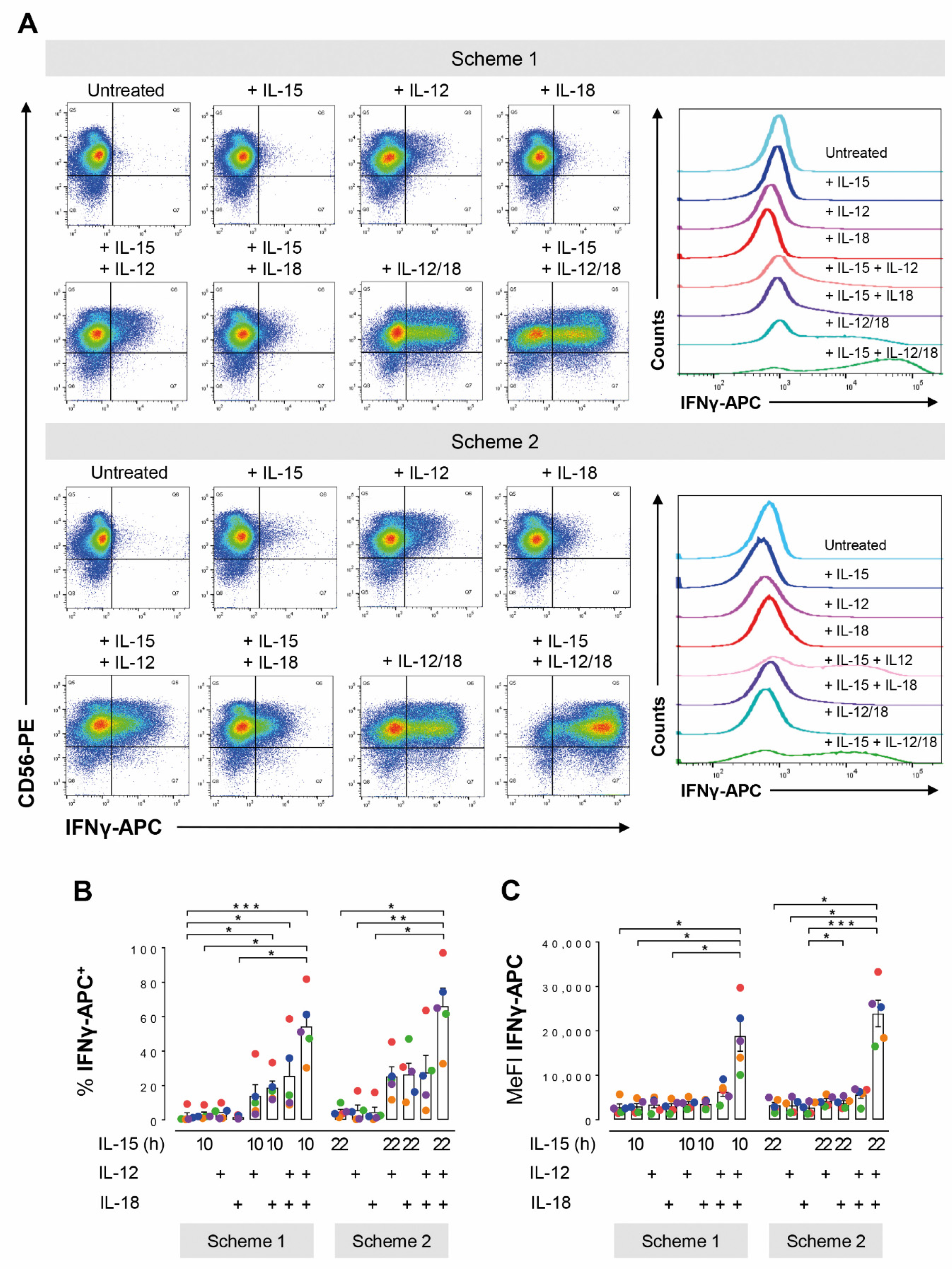
3.1. Long- and Short-Term IL-15 Priming Equally Support Cytokine Induced IFNγ Production in NK Cells
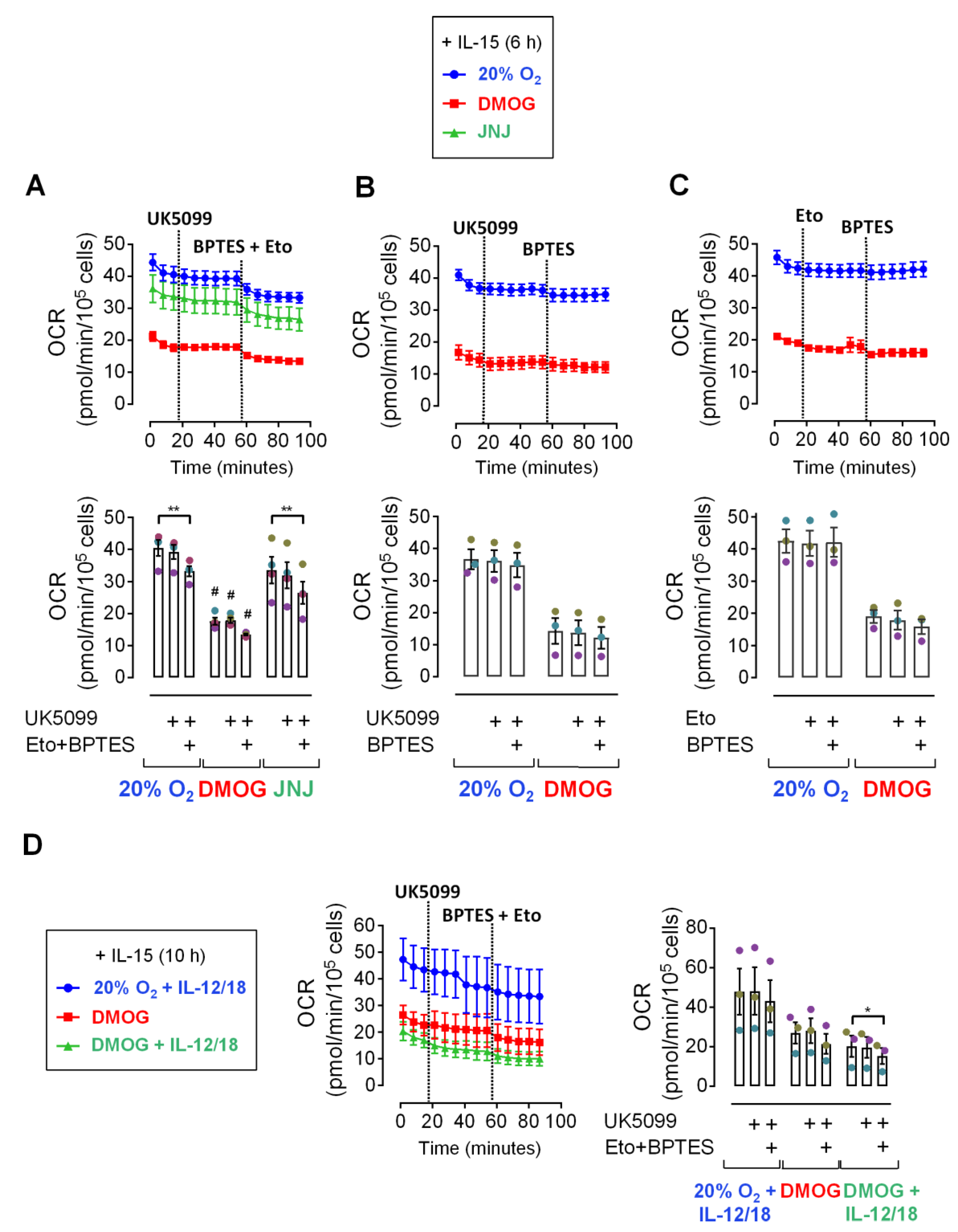
3.2. Pyruvat Does Not Fuel Respiration in IL-15 Primed and IL-12/IL-18 Stimulated NK Cells
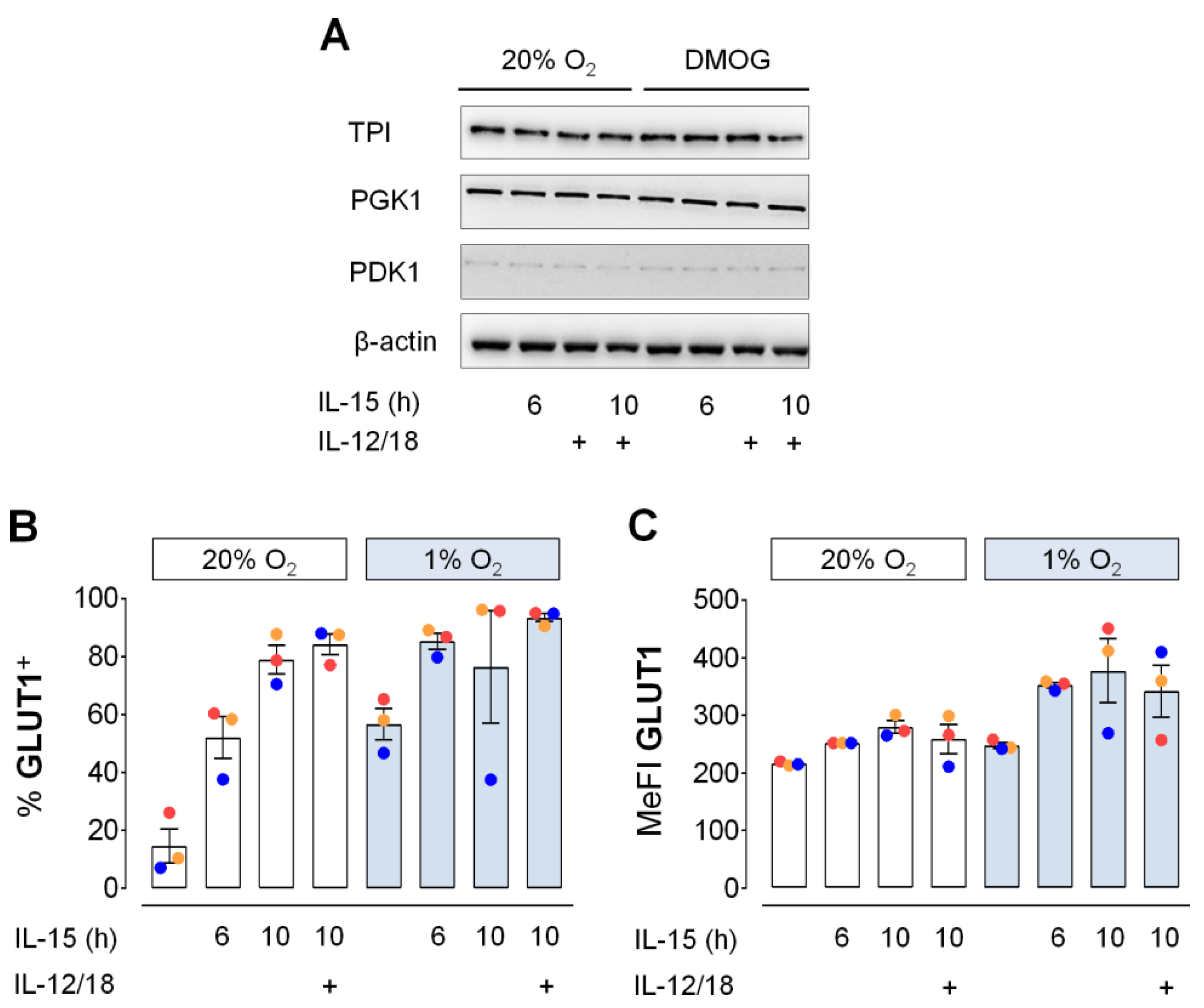
3.3. Glycolytic Protein Expression in Primed NK Cells under Hypoxia
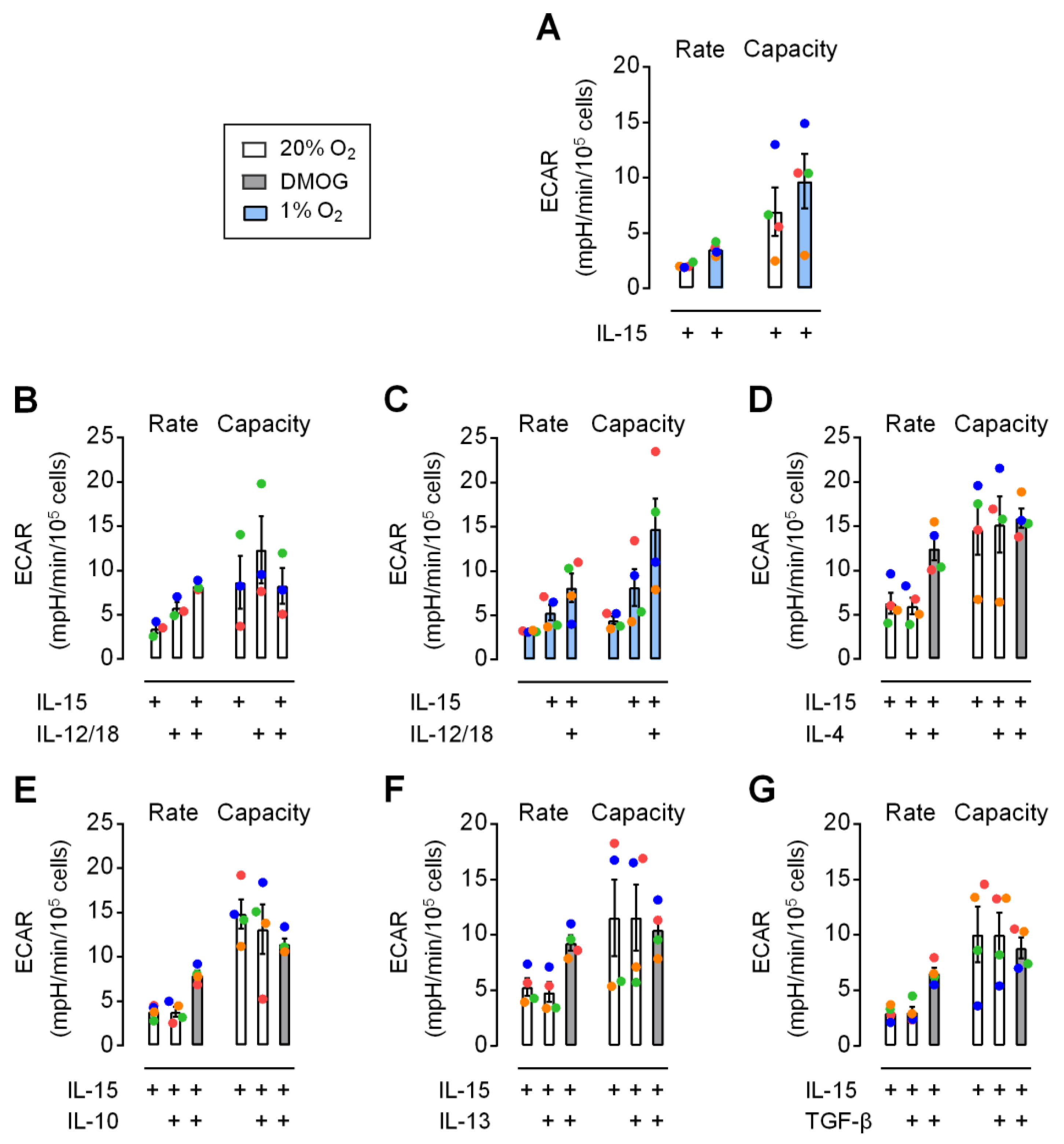
3.4. Moderate Early Increase in NK Cell Glycolysis through Hypoxia and Inflammatory Cytokines
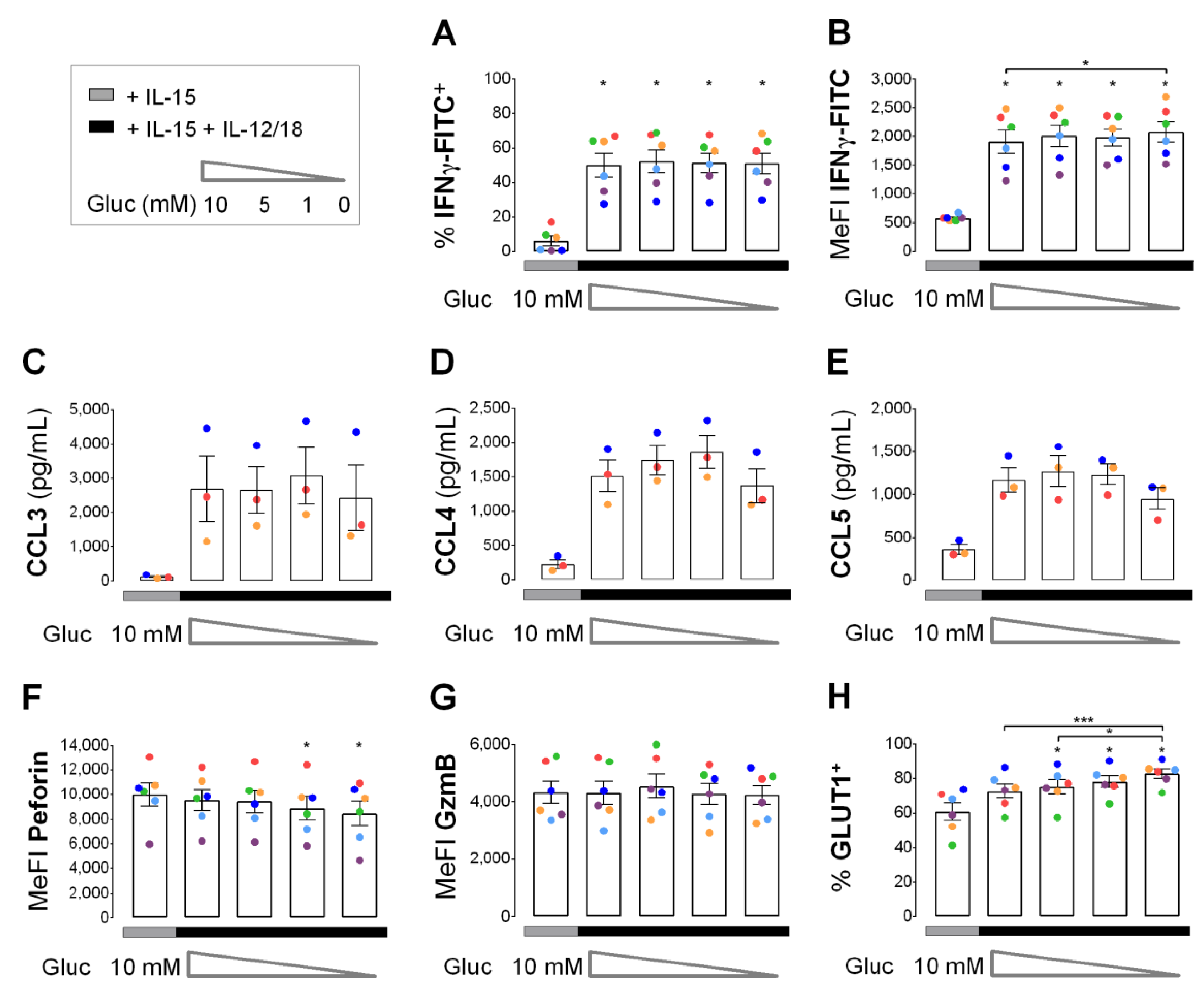
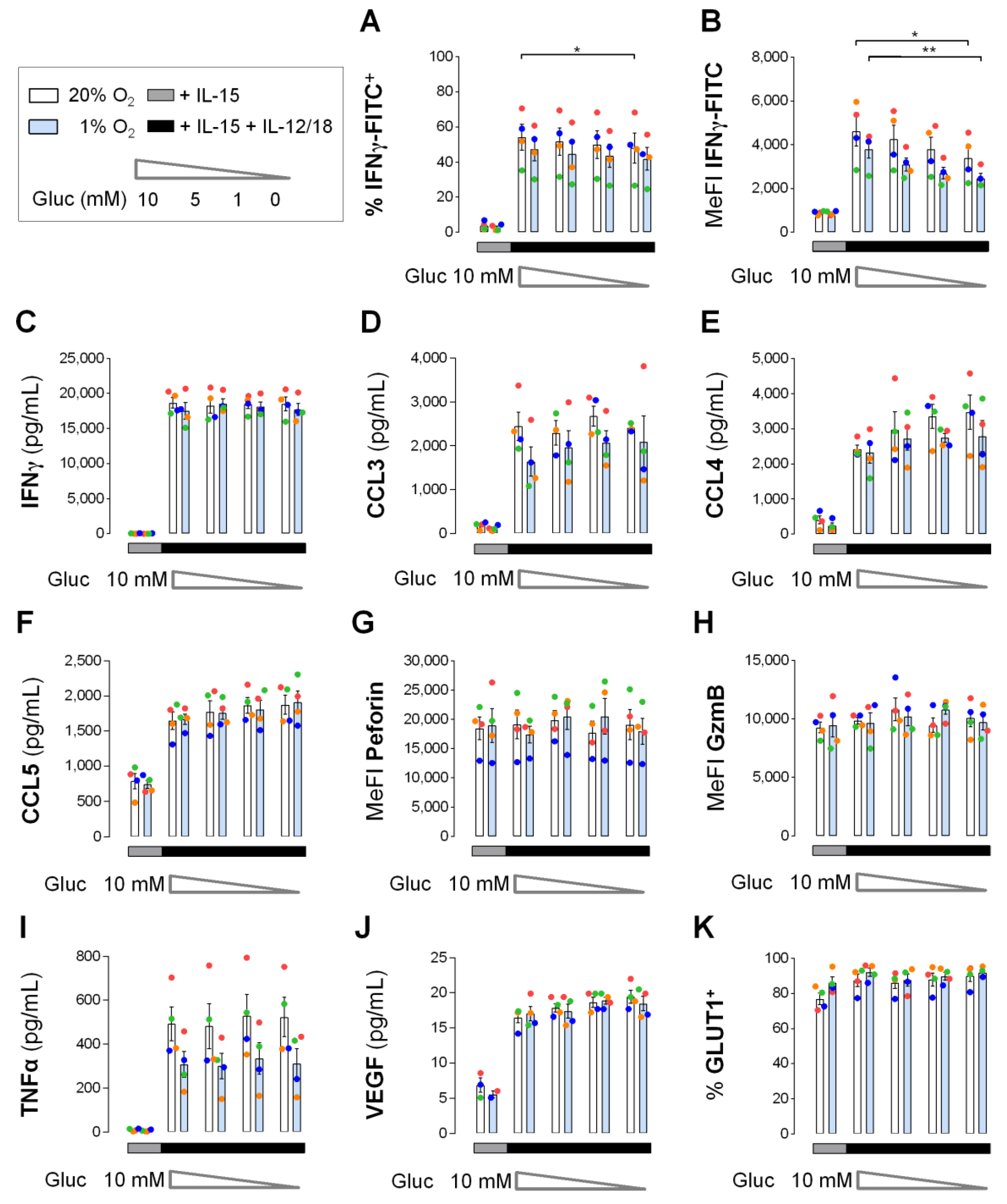
3.5. Early Cytokine Induced IFNγ and Chemokine Responses in NK Cells do not Depend on Glucose
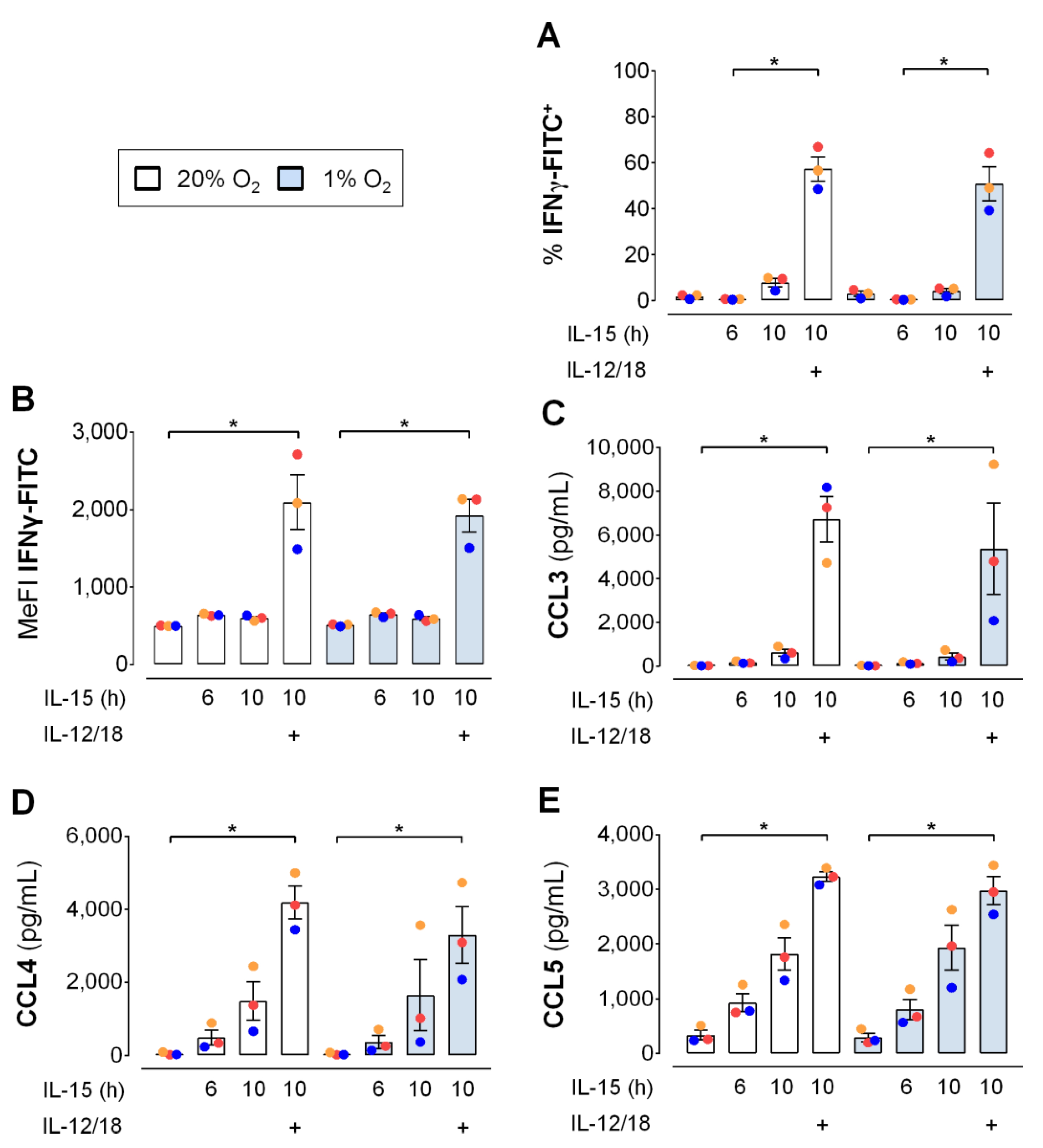
3.6. Hypoxia Hardly Affects Early Cytokine Induced IFNγ Production and Chemokine Release by NK Cells
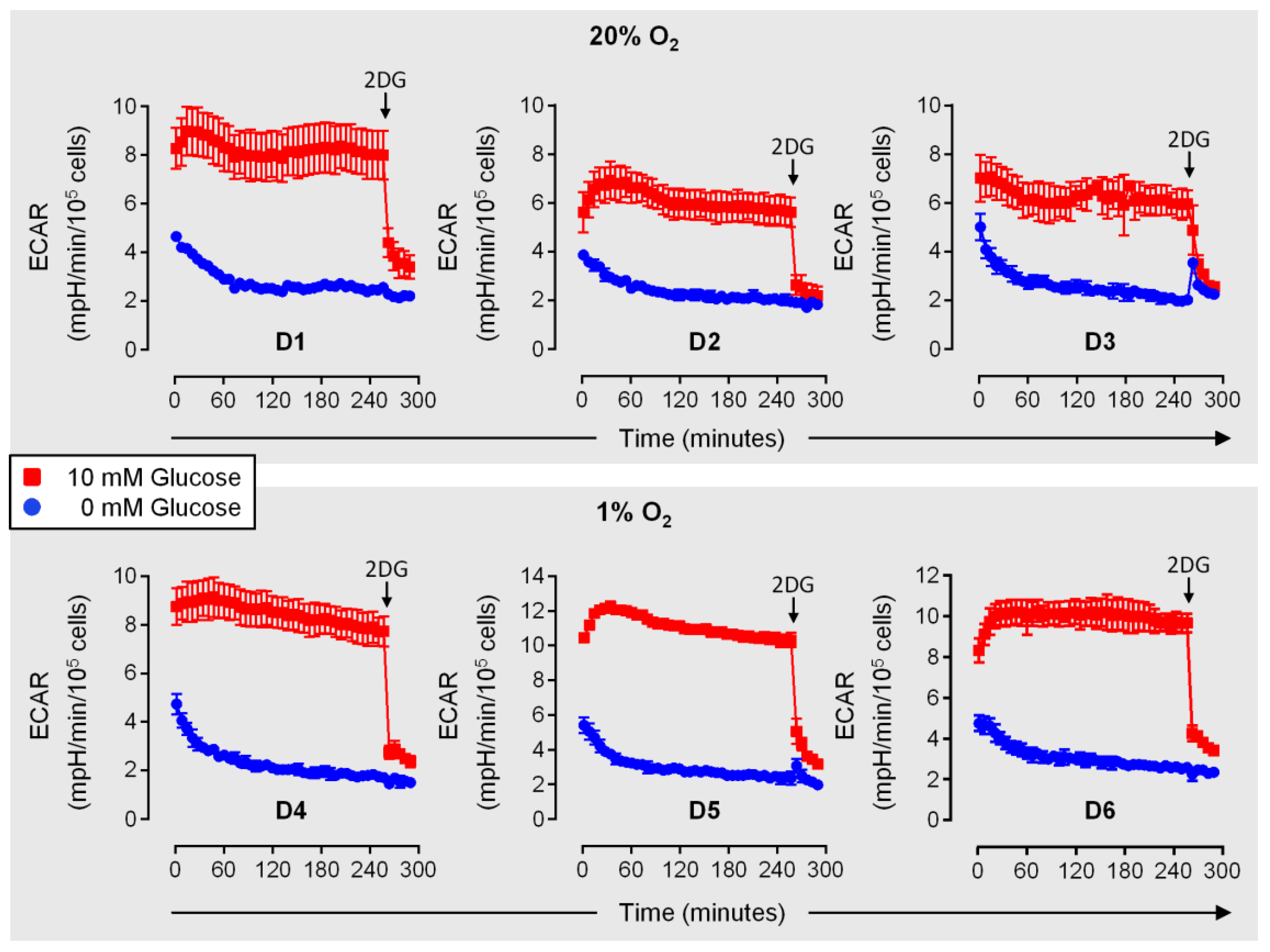
3.7. Efficient Early Cytokine Release from Hypoxic NK Cells Despite Glucose Deprivation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Caligiuri, M.A. Human natural killer cells. Blood 2008, 112, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Gyurova, I.E.; Waggoner, S.N. Mutually assured destruction: The cold war between viruses and natural killer cells. Curr. Opin. Virol. 2019, 34, 130–139. [Google Scholar] [CrossRef]
- Geiger, T.L.; Sun, J.C. Development and maturation of natural killer cells. Curr. Opin. Immunol. 2016, 39, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Carson, W.E.; Fehniger, T.A.; Haldar, S.; Eckhert, K.; Lindemann, M.J.; Lai, C.F.; Croce, C.M.; Baumann, H.; Caligiuri, M.A. A potential role for interleukin-15 in the regulation of human natural killer cell survival. J. Clin. Investig. 1997, 99, 937–943. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.A.; Bush, J.E.; Fehniger, T.A.; VanDeusen, J.B.; Waite, R.E.; Liu, Y.; Aguila, H.L.; Caligiuri, M.A. In vivo evidence for a dependence on interleukin 15 for survival of natural killer cells. Blood 2002, 100, 3633–3638. [Google Scholar] [CrossRef]
- Velasquez, S.Y.; Killian, D.; Schulte, J.; Sticht, C.; Thiel, M.; Lindner, H.A. Short term hypoxia synergizes with interleukin 15 priming in driving glycolytic gene transcription and supports human natural killer cell activities. J. Biol. Chem. 2016, 291, 12960–12977. [Google Scholar] [CrossRef] [Green Version]
- Allavena, P.; Giardina, G.; Bianchi, G.; Mantovani, A. IL-15 is chemotactic for natural killer cells and stimulates their adhesion to vascular endothelium. J. Leukoc. Biol. 1997, 61, 729–735. [Google Scholar] [CrossRef]
- Lucas, M.; Schachterle, W.; Oberle, K.; Aichele, P.; Diefenbach, A. Dendritic cells prime natural killer cells by trans-presenting interleukin 15. Immunity 2007, 26, 503–517. [Google Scholar] [CrossRef] [Green Version]
- Fehniger, T.A.; Cai, S.F.; Cao, X.; Bredemeyer, A.J.; Presti, R.M.; French, A.R.; Ley, T.J. Acquisition of murine NK cell cytotoxicity requires the translation of a pre-existing pool of granzyme B and perforin mRNAs. Immunity 2007, 26, 798–811. [Google Scholar] [CrossRef] [Green Version]
- Nandagopal, N.; Ali, A.K.; Komal, A.K.; Lee, S.H. The critical role of IL-15-PI3K-mTOR pathway in natural killer cell effector functions. Front. Immunol. 2014, 5, 187. [Google Scholar] [CrossRef]
- Luu, T.T.; Nguyen, N.-A.; Ganesan, S.; Meinke, S.; Kadri, N.; Alici, E.; Höglund, P. A brief IL-15 pulse results in JAK3-dependent phosphorylation of ITAM-associated signaling molecules and a long-lasting priming imprint in mouse NK cells. Available online: https://ssrn.com/abstract=3351831 (accessed on 14 March 2020).
- Brady, J.; Carotta, S.; Thong, R.P.; Chan, C.J.; Hayakawa, Y.; Smyth, M.J.; Nutt, S.L. The interactions of multiple cytokines control NK cell maturation. J. Immunol. 2010, 185, 6679–6688. [Google Scholar] [CrossRef] [Green Version]
- Fauriat, C.; Long, E.O.; Ljunggren, H.G.; Bryceson, Y.T. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood 2010, 115, 2167–2176. [Google Scholar] [CrossRef] [Green Version]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- O’Sullivan, T.; Saddawi-Konefka, R.; Vermi, W.; Koebel, C.M.; Arthur, C.; White, J.M.; Uppaluri, R.; Andrews, D.M.; Ngiow, S.F.; Teng, M.W.L.; et al. Cancer immunoediting by the innate immune system in the absence of adaptive immunity. J. Exp. Med. 2012, 209, 1869–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grayson, M.H.; Holtzman, M.J. Chemokine complexity—The case for CCL5. Am. J. Resp. Cell. Mol. Biol. 2006, 35, 143–146. [Google Scholar] [CrossRef]
- Maghazachi, A.A. Role of chemokines in the biology of natural killer cells. In The Chemokine System in Experimental and Clinical Hematology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 37–58. [Google Scholar]
- Balkwill, F. Tumour necrosis factor and cancer. Nat. Rev. Cancer 2009, 9, 361–371. [Google Scholar] [CrossRef]
- Goldszmid, R.S.; Caspar, P.; Rivollier, A.; White, S.; Dzutsev, A.; Hieny, S.; Kelsall, B.; Trinchieri, G.; Sher, A. NK cell-derived interferon-gamma orchestrates cellular dynamics and the differentiation of monocytes into dendritic cells at the site of infection. Immunity 2012, 36, 1047–1059. [Google Scholar] [CrossRef] [Green Version]
- Mattei, F.; Schiavoni, G.; Belardelli, F.; Tough, D.F. IL-15 is expressed by dendritic cells in response to type iifn, double-stranded RNA or lipopolysaccharide and promotes dendritic cell activation. J. Immunol. 2001, 167, 1179–1187. [Google Scholar] [CrossRef]
- Ahmad, R.; Ennaciri, J.; Cordeiro, P.; El Bassam, S.; Menezes, J. Herpes simplex virus-1 up-regulates IL-15 gene expression in monocytic cells through the activation of protein tyrosine kinase and PKC zeta/lambda signaling pathways. J. Mol. Biol. 2007, 367, 25–35. [Google Scholar] [CrossRef] [PubMed]
- de Veerdonk, F.L.V.; Netea, M.G.; Dinarello, C.A.; Joosten, L.A.B. Inflammasome activation and IL-1 beta and IL-18 processing during infection. Trends Immunol. 2011, 32, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Varma, T.K.; Lin, C.Y.; Toliver-Kinsky, T.E.; Sherwood, E.R. Endotoxin-induced gamma interferon production: Contributing cell types and key regulatory factors. Clin. Diagn. Lab. Immunol. 2002, 9, 530–543. [Google Scholar] [CrossRef] [Green Version]
- Pahl, J.H.W.; Cerwenka, A.; Ni, J. Memory-like NK cells: Remembering a previous activation by cytokines and nk cell receptors. Front. Immunol. 2018, 9, 2796. [Google Scholar] [CrossRef] [PubMed]
- Glas, R.; Franksson, L.; Une, C.; Eloranta, M.L.; Ohlen, C.; Orn, A.; Karre, K. Recruitment and activation of natural killer (NK) cells in vivo determined by the target cell phenotype. An adaptive component of NK cell-mediated responses. J. Exp. Med. 2000, 191, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregoire, C.; Chasson, L.; Luci, C.; Tomasello, E.; Geissmann, F.; Vivier, E.; Walzer, T. The trafficking of natural killer cells. Immunol. Rev. 2007, 220, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Natuk, R.J.; Welsh, R.M. Accumulation and chemotaxis of natural killer/large granular lymphocytes at sites of virus replication. J. Immunol. 1987, 138, 877–883. [Google Scholar]
- Basse, P.; Herberman, R.B.; Nannmark, U.; Johansson, B.R.; Hokland, M.; Wasserman, K.; Goldfarb, R.H. Accumulation of adoptively transferred adherent, lymphokine-activated killer cells in murine metastases. J. Exp. Med. 1991, 174, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Herzig, D.S.; Driver, B.R.; Fang, G.P.; Toliver-Kinsky, T.E.; Shute, E.N.; Sherwood, E.R. Regulation of lymphocyte trafficking by CXC chemokine receptor 3 during septic shock. Am. J. Resp. Crit. Care Med. 2012, 185, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Herzig, D.S.; Luan, L.M.; Bohannon, J.K.; Toliver-Kinsky, T.E.; Guo, Y.; Sherwood, E.R. The role of CXCL10 in the pathogenesis of experimental septic shock. Crit. Care 2014, 18, R113. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Patil, N.K.; Luan, L.M.; Bohannon, J.K.; Sherwood, E.R. The biology of natural killer cells during sepsis. Immunology 2018, 153, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Unsinger, J.; Davis, C.G.; Muenzer, J.T.; Ferguson, T.A.; Chang, K.; Osborne, D.F.; Clark, A.T.; Coopersmith, C.M.; McDunn, J.E.; et al. IL-15 prevents apoptosis, reverses innate and adaptive immune dysfunction and improves survival in sepsis. J. Immunol. 2010, 184, 1401–1409. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Luan, L.M.; Patil, N.K.; Sherwood, E.R. Immunobiology of the IL-15/IL-15r alpha complex as an antitumor and antiviral agent. Cytokine Growth Factor Rev. 2017, 38, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.S.; Ahmed, N.; Berridge, M.V. Acute regulation of glucose transport after activation of human peripheral blood neutrophils by phorbol myristate acetate, fMLP and granulocyte-macrophage colony-stimulating factor. Blood 1998, 91, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Eltzschig, H.K.; Carmeliet, P. Hypoxia and inflammation. N. Engl. J. Med. 2011, 364, 656–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, C.T.; Colgan, S.P. Regulation of immunity and inflammation by hypoxia in immunological niches. Nat. Rev. Immunol. 2017, 17, 774–785. [Google Scholar] [CrossRef]
- Buck, M.D.; Sowell, R.T.; Kaech, S.M.; Pearce, E.L. Metabolic instruction of immunity. Cell 2017, 169, 570–586. [Google Scholar] [CrossRef]
- Palsson-McDermott, E.M.; O’Neill, L.A.J. The Warburg effect then and now: From cancer to inflammatory diseases. Bioessays 2013, 35, 965–973. [Google Scholar] [CrossRef]
- Schafer, J.R.; Salzillo, T.C.; Chakravarti, N.; Kararoudi, M.N.; Trikha, P.; Foltz, J.A.; Wang, R.N.; Li, S.L.; Lee, D.A. Education-dependent activation of glycolysis promotes the cytolytic potency of licensed human natural killer cells. J. Allergy. Clin. Immunol. 2019, 143, 346–358. [Google Scholar] [CrossRef]
- Pfeifer, C.; Highton, A.J.; Peine, S.; Sauter, J.; Schmidt, A.H.; Bunders, M.J.; Altfeld, M.; Korner, C. Natural killer cell education is associated with a distinct glycolytic profile. Front. Immunol. 2018, 9, 3020. [Google Scholar] [CrossRef] [Green Version]
- Mah, A.Y.; Cooper, M.A. Metabolic regulation of natural killer cell IFN-gamma production. Crit. Rev. Immunol. 2016, 36, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, C.M.; Finlay, D.K. What fuels natural killers? Metabolism and NK cell responses. Front. Immunol. 2017, 8, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assmann, N.; O’Brien, K.L.; Donnelly, R.P.; Dyck, L.; Zaiatz-Bittencourt, V.; Loftus, R.M.; Heinrich, P.; Oefner, P.J.; Lynch, L.; Gardiner, C.M.; et al. Srebp-controlled glucose metabolism is essential for NK cell functional responses. Nat. Immunol. 2017, 18, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.L.; Finlay, D.K. Immunometabolism and natural killer cell responses. Nat. Rev. Immunol. 2019, 19, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Mah, A.Y.; Rashidi, A.; Keppel, M.P.; Saucier, N.; Moore, E.K.; Alinger, J.B.; Tripathy, S.K.; Agarwal, S.K.; Jeng, E.K.; Wong, H.C.; et al. Glycolytic requirement for NK cell cytotoxicity and cytomegalovirus control. JCI Insight 2017, 2, e95128. [Google Scholar] [CrossRef]
- Cong, J.J.; Wang, X.W.; Zheng, X.H.; Wang, D.; Fu, B.Q.; Sun, R.; Tian, Z.G.; Wei, H.M. Dysfunction of natural killer cells by FBP1-induced inhibition of glycolysis during lung cancer progression. Cell Metab. 2018, 28, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Keppel, M.P.; Saucier, N.; Mah, A.Y.; Vogel, T.P.; Cooper, M.A. Activation-specific metabolic requirements for NK cell IFN-gamma production. J. Immunol. 2015, 194, 1954–1962. [Google Scholar] [CrossRef]
- Coulibaly, A.; Bettendorf, A.; Kostina, E.; Figueiredo, A.S.; Velasquez, S.Y.; Bock, H.G.; Thiel, M.; Lindner, H.A.; Barbarossa, M.V. Interleukin-15 signaling in HIF-1alpha regulation in natural killer cells, insights through mathematical models. Front. Immunol. 2019, 10, 2401. [Google Scholar] [CrossRef]
- Semenza, G.L. Oxygen sensing, homeostasis and disease reply. N. Engl. J. Med. 2011, 365, 1846. [Google Scholar] [CrossRef] [Green Version]
- Loeffler, D.A.; Juneau, P.L.; Heppner, G.H. Natural killer-cell activity under conditions reflective of tumor micro-environment. Int. J. Cancer 1991, 48, 895–899. [Google Scholar] [CrossRef]
- Semenza, G.L. Regulation of metabolism by hypoxia-inducible factor 1. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, J.J.; Qin, S.; Unutmaz, D.; Soler, D.; Murphy, K.E.; Hodge, M.R.; Wu, L.; Butcher, E.C. Unique subpopulations of CD56+ NK and NK-T peripheral blood lymphocytes identified by chemokine receptor expression repertoire. J. Immunol. 2001, 166, 6477–6482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stonier, S.W.; Schluns, K.S. Trans-presentation: A novel mechanism regulating IL-15 delivery and responses. Immunol. Lett. 2010, 127, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppenheimer-Marks, N.; Brezinschek, R.I.; Mohamadzadeh, M.; Vita, R.; Lipsky, P.E. Interleukin 15 is produced by endothelial cells and increases the transendothelial migration of t cells in vitro and in the scid mouse-human rheumatoid arthritis model in vivo. J. Clin. Investig. 1998, 101, 1261–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsen, E.M.; Johansen, F.E.; Jahnsen, F.L.; Lundin, K.E.; Scholz, T.; Brandtzaeg, P.; Haraldsen, G. Cytokine profiles of cultured microvascular endothelial cells from the human intestine. Gut 1998, 42, 635–642. [Google Scholar] [CrossRef]
- Liu, X.; Zuo, Y.; Zhang, W.; Yang, D.; Xiong, C.; Zhang, X. Expression of interleukin-15 and its receptor on the surface of stimulated human umbilical vein endothelial cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2009, 29, 527–534. [Google Scholar] [CrossRef]
- Walzer, T.; Dalod, M.; Robbins, S.H.; Zitvogel, L.; Vivier, E. Natural-killer cells and dendritic cells: “L’union fait la force”. Blood 2005, 106, 2252–2258. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.M.; Wolf, A.S.; Goodier, M.R.; Riley, E.M. Synergy between common γ chain family cytokines and IL-18 potentiates innate and adaptive pathways of NK cell activation. Front. Immunol. 2016, 7, 101. [Google Scholar] [CrossRef] [Green Version]
- Wagstaffe, H.R.; Nielsen, C.M.; Riley, E.M.; Goodier, M.R. IL-15 promotes polyfunctional NK cell responses to influenza by boosting IL-12 production. J. Immunol. 2018, 200, 2738–2747. [Google Scholar] [CrossRef] [Green Version]
- Tanner, L.B.; Goglia, A.G.; Wei, M.H.; Sehgal, T.; Parsons, L.R.; Park, J.O.; White, E.; Toettcher, J.E.; Rabinowitz, J.D. Four key steps control glycolytic flux in mammalian cells. Cell Syst. 2018, 7, 49–62. [Google Scholar] [CrossRef]
- Felices, M.; Lenvik, A.J.; McElmurry, R.; Chu, S.; Hinderlie, P.; Bendzick, L.; Geller, M.A.; Tolar, J.; Blazar, B.R.; Miller, J.S. Continuous treatment with IL-15 exhausts human NK cells via a metabolic defect. JCI Insight 2018, 3, e96219. [Google Scholar] [CrossRef] [PubMed]
- Parodi, M.; Raggi, F.; Cangelosi, D.; Manzini, C.; Balsamo, M.; Blengio, F.; Eva, A.; Varesio, L.; Pietra, G.; Moretta, L.; et al. Hypoxia modifies the transcriptome of human NK cells, modulates their immunoregulatory profile and influences nk cell subset migration. Front. Immunol. 2018, 9, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poznanski, S.M.; Ashkar, A.A. What defines NK cell functional fate: Phenotype or metabolism? Front. Immunol. 2019, 10, 1414. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Wang, X.; Gao, X.; Liu, X. Regulation of T cell development and activation by creatine kinase B. PLoS ONE 2009, 4, e5000. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velásquez, S.Y.; Himmelhan, B.S.; Kassner, N.; Coulibaly, A.; Schulte, J.; Brohm, K.; Lindner, H.A. Innate Cytokine Induced Early Release of IFNγ and CC Chemokines from Hypoxic Human NK Cells Is Independent of Glucose. Cells 2020, 9, 734. https://doi.org/10.3390/cells9030734
Velásquez SY, Himmelhan BS, Kassner N, Coulibaly A, Schulte J, Brohm K, Lindner HA. Innate Cytokine Induced Early Release of IFNγ and CC Chemokines from Hypoxic Human NK Cells Is Independent of Glucose. Cells. 2020; 9(3):734. https://doi.org/10.3390/cells9030734
Chicago/Turabian StyleVelásquez, Sonia Y., Bianca S. Himmelhan, Nina Kassner, Anna Coulibaly, Jutta Schulte, Kathrin Brohm, and Holger A. Lindner. 2020. "Innate Cytokine Induced Early Release of IFNγ and CC Chemokines from Hypoxic Human NK Cells Is Independent of Glucose" Cells 9, no. 3: 734. https://doi.org/10.3390/cells9030734
APA StyleVelásquez, S. Y., Himmelhan, B. S., Kassner, N., Coulibaly, A., Schulte, J., Brohm, K., & Lindner, H. A. (2020). Innate Cytokine Induced Early Release of IFNγ and CC Chemokines from Hypoxic Human NK Cells Is Independent of Glucose. Cells, 9(3), 734. https://doi.org/10.3390/cells9030734