Kalirin Interacts with TRAPP and Regulates Rab11 and Endosomal Recycling

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid DNA
2.2. Expression and Purification of Tagged Proteins
2.3. Cell Culture and Transfection
2.4. Preparation of Total Cellular Membranes
2.5. Guanine Nucleotide Exchange Assay
2.6. Transferrin Recycling
2.7. Immunohistochemistry and Western Blot
2.8. Cryo-Immunogold Electron Microscopy
3. Results
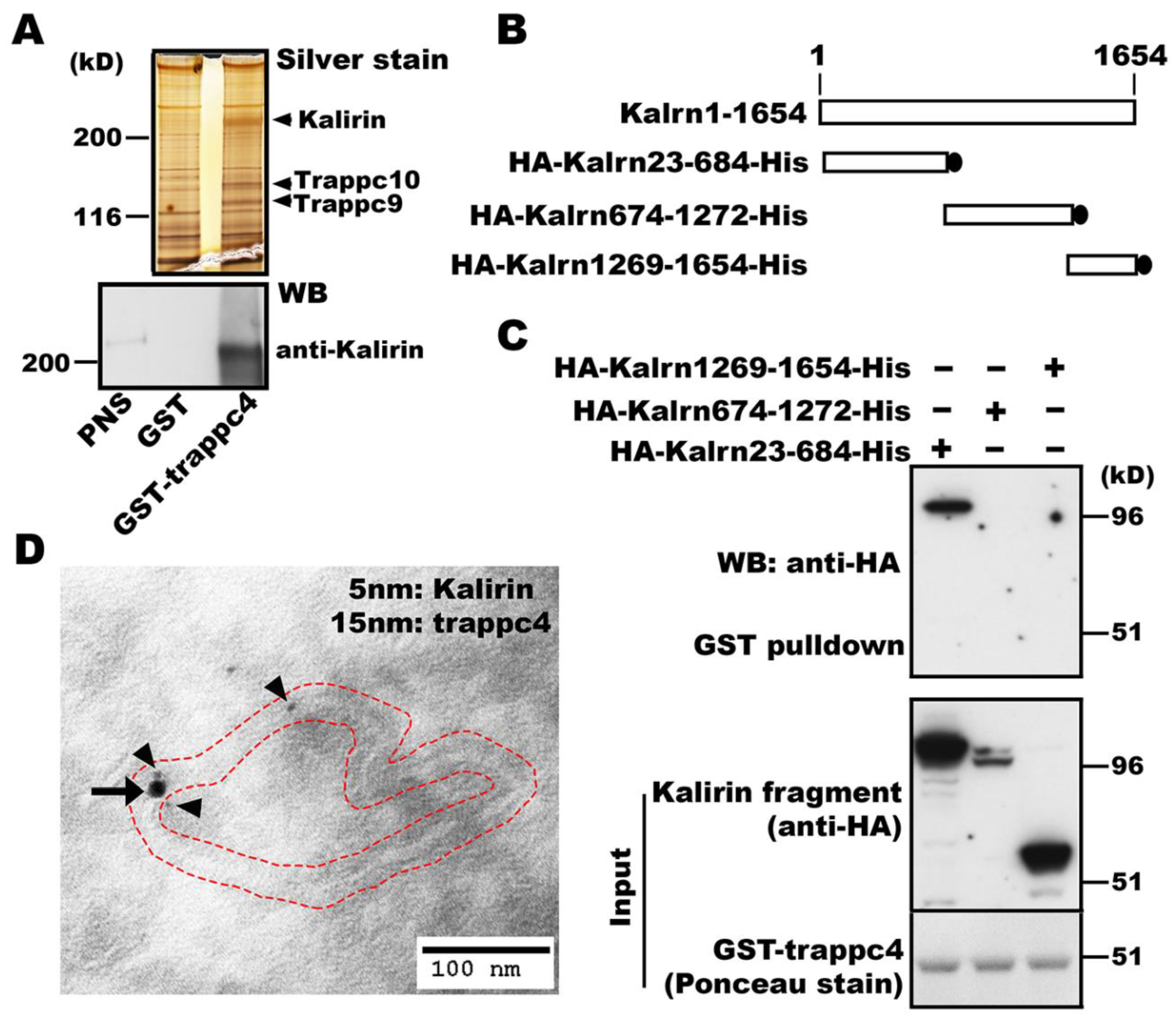
3.1. Kalirin Interacts with mTRAPP
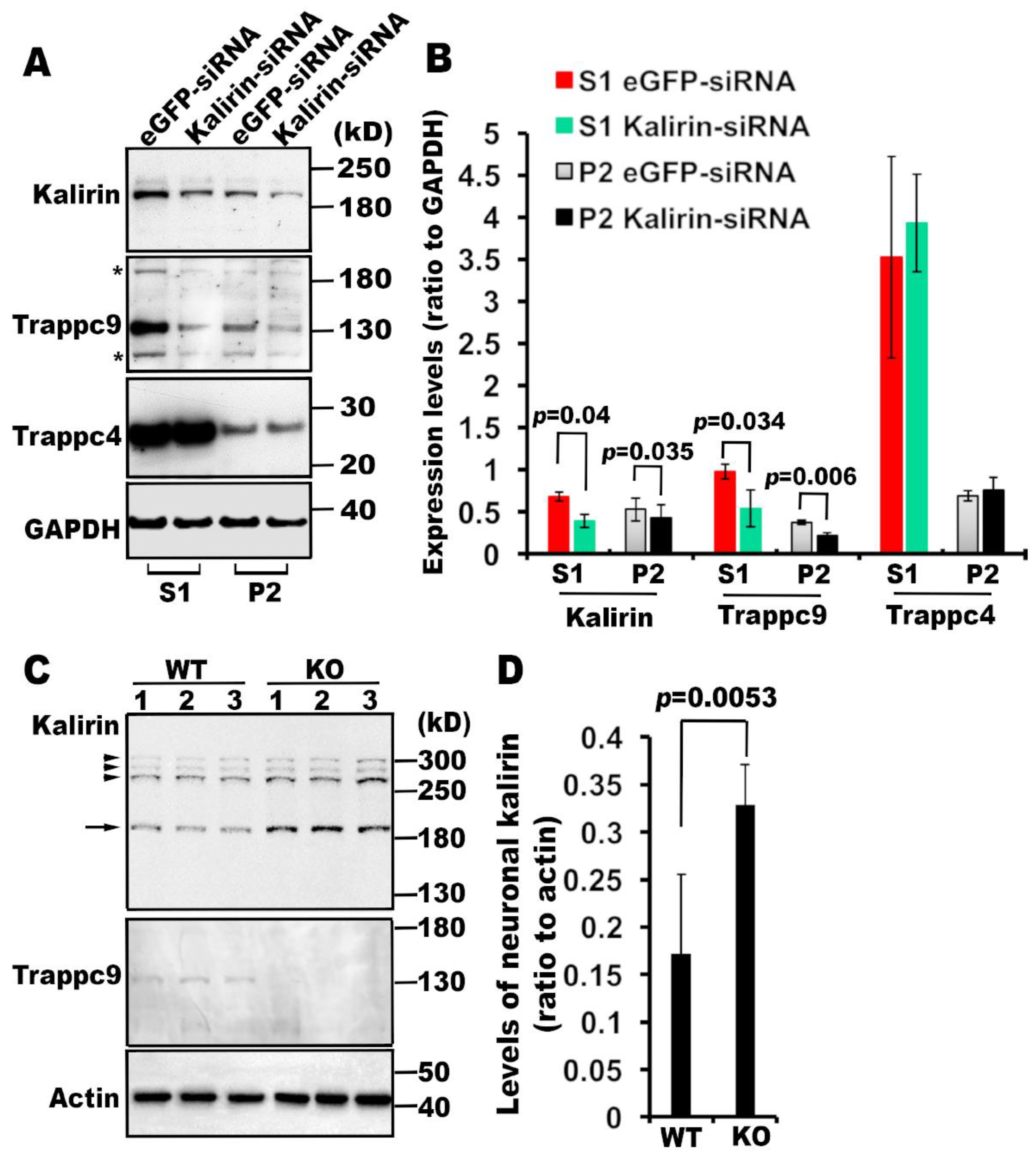
3.2. Kalirin is Associated with mTRAPP-II
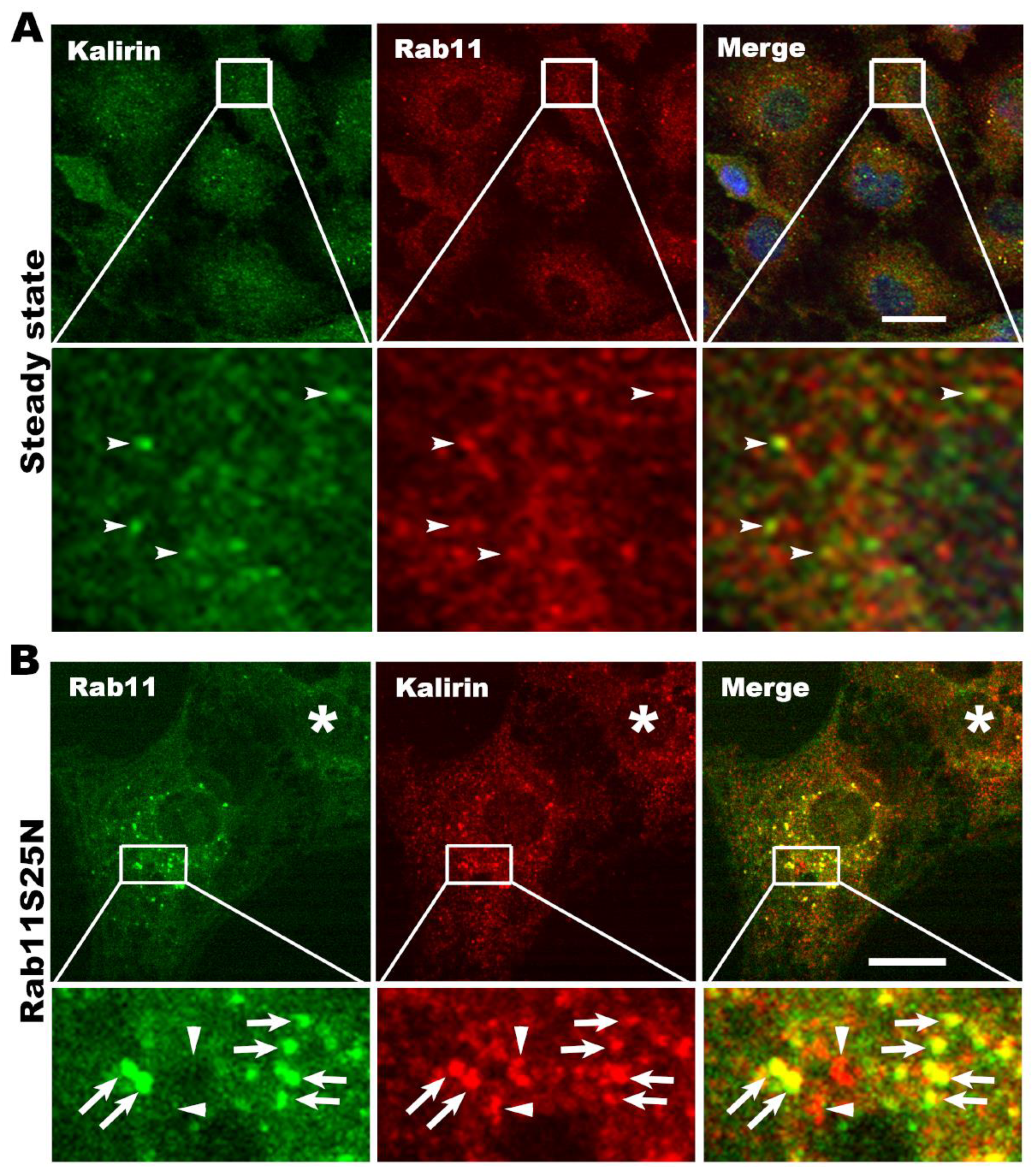
3.3. Kalirin Regulates the Function of Rab11
3.4. Kalirin Functions at Rab11 Positive Recycling Endosomes
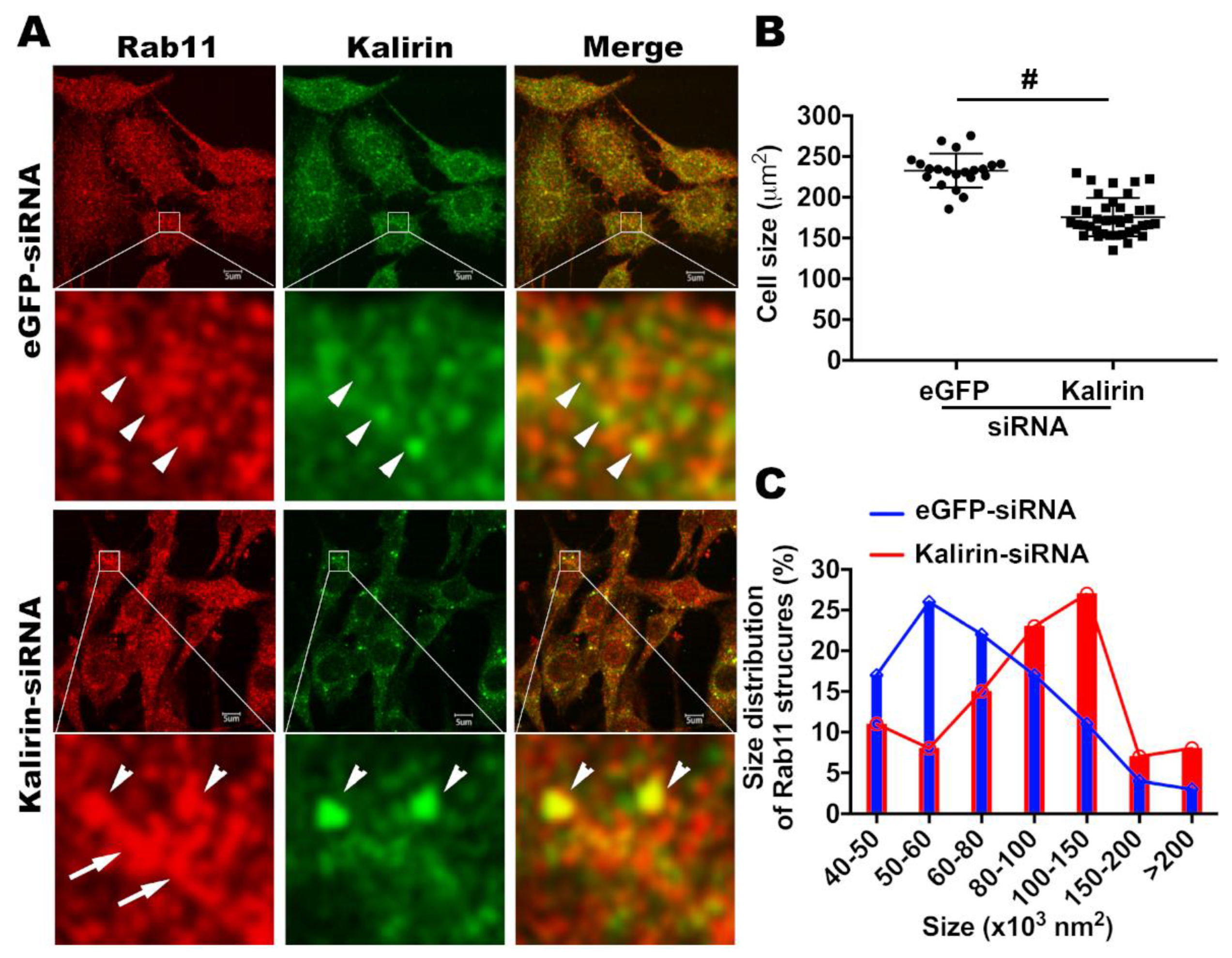
3.5. Knockdown of Kalirin Causes Tubulation of Recycling Endosomes
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, M.C.; Caswell, P.T.; Norman, J.C. Endocytic recycling pathways: Emerging regulators of cell migration. Curr. Opin. Cell Biol. 2006, 18, 549–557. [Google Scholar] [CrossRef]
- Woolfrey, K.M.; Srivastava, D.P. Control of Dendritic Spine Morphological and Functional Plasticity by Small GTPases. Neural Plast. 2016, 2016, 3025948. [Google Scholar] [CrossRef] [Green Version]
- Holtmaat, A.; Caroni, P. Functional and structural underpinnings of neuronal assembly formation in learning. Nat. Neurosci. 2016, 19, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Anitei, M.; Hoflack, B. Bridging membrane and cytoskeleton dynamics in the secretory and endocytic pathways. Nat. Cell Biol. 2012, 14, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2001, 2, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Phuyal, S.; Farhan, H. Multifaceted Rho GTPase Signaling at the Endomembranes. Front. Cell Dev. Biol. 2019, 7, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, C.D.; Ridley, A.J. Rho GTPase signaling complexes in cell migration and invasion. J. Cell Biol. 2018, 217, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Margiotta, A.; Bucci, C. Coordination between Rac1 and Rab Proteins: Functional Implications in Health and Disease. Cells 2019, 8, 396. [Google Scholar] [CrossRef] [Green Version]
- Bouchet, J.; McCaffrey, M.W.; Graziani, A.; Alcover, A. The functional interplay of Rab11, FIP3 and Rho proteins on the endosomal recycling pathway controls cell shape and symmetry. Small GTPases 2018, 9, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Vestre, K.; Kjos, I.; Guadagno, N.A.; Borg Distefano, M.; Kohler, F.; Fenaroli, F.; Bakke, O.; Progida, C. Rab6 regulates cell migration and invasion by recruiting Cdc42 and modulating its activity. Cell. Mol. Life Sci. 2019, 76, 2593–2614. [Google Scholar] [CrossRef]
- Kunita, R.; Otomo, A.; Mizumura, H.; Suzuki-Utsunomiya, K.; Hadano, S.; Ikeda, J.E. The Rab5 activator ALS2/alsin acts as a novel Rac1 effector through Rac1-activated endocytosis. J. Biol. Chem. 2007, 282, 16599–16611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frasa, M.A.; Maximiano, F.C.; Smolarczyk, K.; Francis, R.E.; Betson, M.E.; Lozano, E.; Goldenring, J.; Seabra, M.C.; Rak, A.; Ahmadian, M.R.; et al. Armus is a Rac1 effector that inactivates Rab7 and regulates E-cadherin degradation. Curr. Biol. 2010, 20, 198–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.M.; Huang, J.; Wang, Y.; Eipper, B.A.; Mains, R.E. Kalirin, a multifunctional Rho guanine nucleotide exchange factor, is necessary for maintenance of hippocampal pyramidal neuron dendrites and dendritic spines. J. Neurosci. 2003, 23, 10593–10603. [Google Scholar] [CrossRef] [PubMed]
- Lipatova, Z.; Segev, N. Ypt/Rab GTPases and their TRAPP GEFs at the Golgi. FEBS Lett. 2019, 593, 2488–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacher, M.; Shahrzad, N.; Kamel, H.; Milev, M.P. TRAPPopathies: An emerging set of disorders linked to variations in the genes encoding transport protein particle (TRAPP)-associated proteins. Traffic 2019, 20, 5–26. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.G.; Raunser, S.; Munger, C.; Wagner, J.; Song, Y.L.; Cygler, M.; Walz, T.; Oh, B.H.; Sacher, M. The architecture of the multisubunit TRAPP I complex suggests a model for vesicle tethering. Cell 2006, 127, 817–830. [Google Scholar] [CrossRef]
- Li, X.; Standley, C.; Sapp, E.; Valencia, A.; Qin, Z.H.; Kegel, K.B.; Yoder, J.; Comer-Tierney, L.A.; Esteves, M.; Chase, K.; et al. Mutant huntingtin impairs vesicle formation from recycling endosomes by interfering with Rab11 activity. Mol. Cell. Biol. 2009, 29, 6106–6116. [Google Scholar] [CrossRef] [Green Version]
- McClory, H.; Wang, X.; Sapp, E.; Gatune, L.W.; Iuliano, M.; Wu, C.Y.; Nathwani, G.; Kegel-Gleason, K.B.; DiFiglia, M.; Li, X. The COOH-terminal domain of huntingtin interacts with RhoGEF kalirin and modulates cell survival. Sci. Rep. 2018, 8, 8000. [Google Scholar] [CrossRef]
- Li, X.; Sapp, E.; Valencia, A.; Kegel, K.B.; Qin, Z.H.; Alexander, J.; Masso, N.; Reeves, P.; Ritch, J.J.; Zeitlin, S.; et al. A function of huntingtin in guanine nucleotide exchange on Rab11. Neuroreport 2008, 19, 1643–1647. [Google Scholar] [CrossRef]
- Li, X.; Sapp, E.; Chase, K.; Comer-Tierney, L.A.; Masso, N.; Alexander, J.; Reeves, P.; Kegel, K.B.; Valencia, A.; Esteves, M.; et al. Disruption of Rab11 activity in a knock-in mouse model of Huntington’s disease. Neurobiol. Dis. 2009, 36, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Slot, J.W.; Geuze, H.J.; Gigengack, S.; Lienhard, G.E.; James, D.E. Immuno-localization of the insulin regulatable glucose transporter in brown adipose tissue of the rat. J. Cell Biol. 1991, 113, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacher, M.; Jiang, Y.; Barrowman, J.; Scarpa, A.; Burston, J.; Zhang, L.; Schieltz, D.; Yates, J.R., 3rd; Abeliovich, H.; Ferro-Novick, S. TRAPP, a highly conserved novel complex on the cis-Golgi that mediates vesicle docking and fusion. EMBO J. 1998, 17, 2494–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrowman, J.; Bhandari, D.; Reinisch, K.; Ferro-Novick, S. TRAPP complexes in membrane traffic: Convergence through a common Rab. Nat. Rev. Mol. Cell Biol. 2010, 11, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Bassik, M.C.; Kampmann, M.; Lebbink, R.J.; Wang, S.; Hein, M.Y.; Poser, I.; Weibezahn, J.; Horlbeck, M.A.; Chen, S.; Mann, M.; et al. A systematic mammalian genetic interaction map reveals pathways underlying ricin susceptibility. Cell 2013, 152, 909–922. [Google Scholar] [CrossRef] [Green Version]
- Rabiner, C.A.; Mains, R.E.; Eipper, B.A. Kalirin: A dual Rho guanine nucleotide exchange factor that is so much more than the sum of its many parts. Neuroscientist 2005, 11, 148–160. [Google Scholar] [CrossRef]
- Penzes, P.; Johnson, R.C.; Alam, M.R.; Kambampati, V.; Mains, R.E.; Eipper, B.A. An isoform of kalirin, a brain-specific GDP/GTP exchange factor, is enriched in the postsynaptic density fraction. J. Biol. Chem. 2000, 275, 6395–6403. [Google Scholar] [CrossRef] [Green Version]
- Morozova, N.; Liang, Y.; Tokarev, A.A.; Chen, S.H.; Cox, R.; Andrejic, J.; Lipatova, Z.; Sciorra, V.A.; Emr, S.D.; Segev, N. TRAPPII subunits are required for the specificity switch of a Ypt-Rab GEF. Nat. Cell Biol. 2006, 8, 1263–1269. [Google Scholar] [CrossRef]
- Thomas, L.L.; Fromme, J.C. GTPase cross talk regulates TRAPPII activation of Rab11 homologues during vesicle biogenesis. J. Cell Biol. 2016, 215, 499–513. [Google Scholar] [CrossRef] [Green Version]
- Maxfield, F.R.; McGraw, T.E. Endocytic recycling. Nat. Rev. Mol. Cell Biol. 2004, 5, 121–132. [Google Scholar] [CrossRef]
- Lin, S.X.; Gundersen, G.G.; Maxfield, F.R. Export from pericentriolar endocytic recycling compartment to cell surface depends on stable, detyrosinated (glu) microtubules and kinesin. Mol. Biol. Cell 2002, 13, 96–109. [Google Scholar] [CrossRef]
- Wilcke, M.; Johannes, L.; Galli, T.; Mayau, V.; Goud, B.; Salamero, J. Rab11 regulates the compartmentalization of early endosomes required for efficient transport from early endosomes to the trans-golgi network. J. Cell Biol. 2000, 151, 1207–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, T.A.; Blizinsky, K.D.; Cobia, D.J.; Cahill, M.E.; Xie, Z.; Sweet, R.A.; Duan, J.; Gejman, P.V.; Wang, L.; Csernansky, J.G.; et al. A sequence variant in human KALRN impairs protein function and coincides with reduced cortical thickness. Nat. Commun. 2014, 5, 4858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchet, J.; Del Rio-Iniguez, I.; Lasserre, R.; Aguera-Gonzalez, S.; Cuche, C.; Danckaert, A.; McCaffrey, M.W.; Di Bartolo, V.; Alcover, A. Rac1-Rab11-FIP3 regulatory hub coordinates vesicle traffic with actin remodeling and T-cell activation. EMBO J. 2016, 35, 1160–1174. [Google Scholar] [CrossRef] [PubMed]
- Derivery, E.; Sousa, C.; Gautier, J.J.; Lombard, B.; Loew, D.; Gautreau, A. The Arp2/3 activator WASH controls the fission of endosomes through a large multiprotein complex. Dev. Cell 2009, 17, 712–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, S.S.; Bassani, S.; Brown, T.C.; Lise, M.F.; Backos, D.S.; El-Husseini, A.; Passafaro, M.; Esteban, J.A. Motor protein-dependent transport of AMPA receptors into spines during long-term potentiation. Nat. Neurosci. 2008, 11, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Penick, E.C.; Edwards, J.G.; Kauer, J.A.; Ehlers, M.D. Recycling endosomes supply AMPA receptors for LTP. Science 2004, 305, 1972–1975. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Salgado, J.M.; Ostroff, L.; Helton, T.D.; Robinson, C.G.; Harris, K.M.; Ehlers, M.D. Plasticity-induced growth of dendritic spines by exocytic trafficking from recycling endosomes. Neuron 2006, 52, 817–830. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Srivastava, D.P.; Photowala, H.; Kai, L.; Cahill, M.E.; Woolfrey, K.M.; Shum, C.Y.; Surmeier, D.J.; Penzes, P. Kalirin-7 controls activity-dependent structural and functional plasticity of dendritic spines. Neuron 2007, 56, 640–656. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Weng, M.; Ke, Y.; Sapp, E.; DiFiglia, M.; Li, X. Kalirin Interacts with TRAPP and Regulates Rab11 and Endosomal Recycling. Cells 2020, 9, 1132. https://doi.org/10.3390/cells9051132
Wang X, Weng M, Ke Y, Sapp E, DiFiglia M, Li X. Kalirin Interacts with TRAPP and Regulates Rab11 and Endosomal Recycling. Cells. 2020; 9(5):1132. https://doi.org/10.3390/cells9051132
Chicago/Turabian StyleWang, Xiaolong, Meiqian Weng, Yuting Ke, Ellen Sapp, Marian DiFiglia, and Xueyi Li. 2020. "Kalirin Interacts with TRAPP and Regulates Rab11 and Endosomal Recycling" Cells 9, no. 5: 1132. https://doi.org/10.3390/cells9051132
APA StyleWang, X., Weng, M., Ke, Y., Sapp, E., DiFiglia, M., & Li, X. (2020). Kalirin Interacts with TRAPP and Regulates Rab11 and Endosomal Recycling. Cells, 9(5), 1132. https://doi.org/10.3390/cells9051132