Homing and Engraftment of Intravenously Administered Equine Cord Blood-Derived Multipotent Mesenchymal Stromal Cells to Surgically Created Cutaneous Wound in Horses: A Pilot Project

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
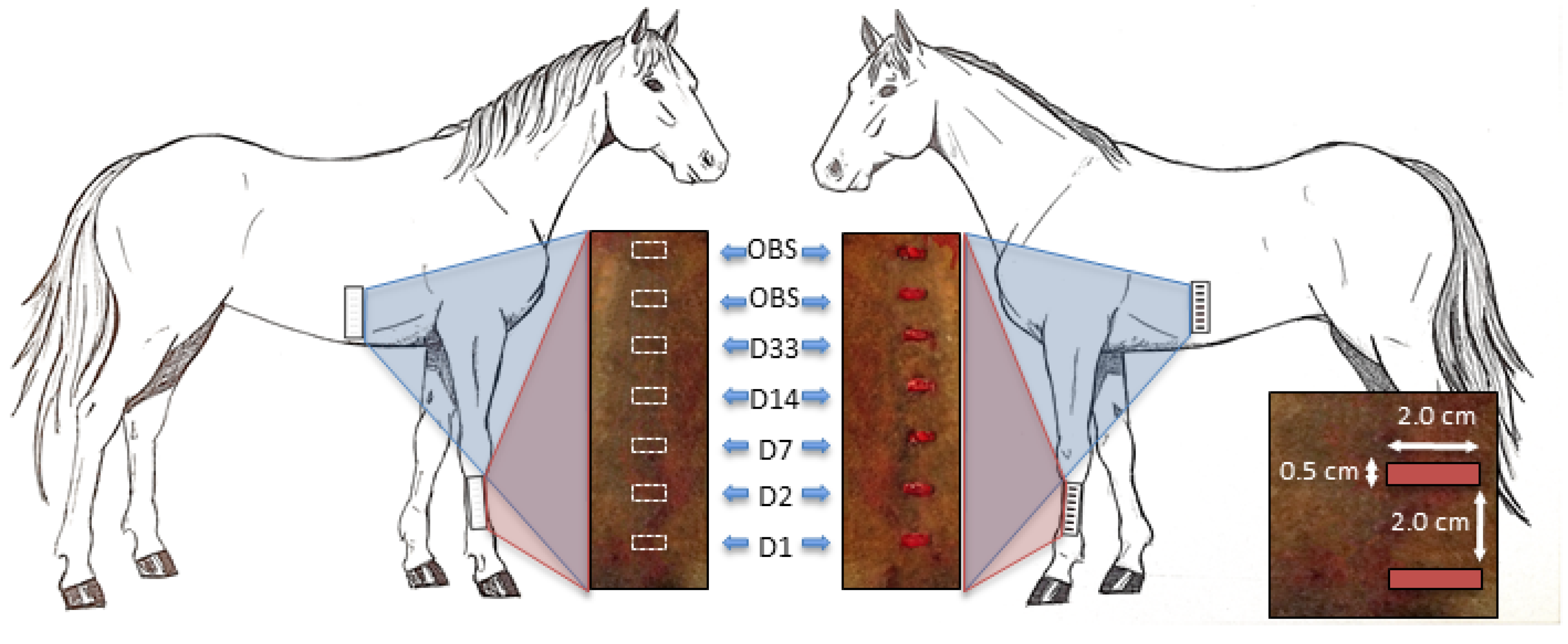
2.1. Overview of the Study Design
2.2. Recipient Animals
2.3. Transducing and Labeling Trial
2.4. Source and Transportation of CB-MSCs
2.5. Preparation of CB-MSCs
2.6. Wound Creation
2.7. Prepared CB-MSC Administration and Monitoring
2.8. Biopsy Collection
2.9. Fluorescence Evaluation of Biopsies
3. Results
3.1. Adverse Reactions
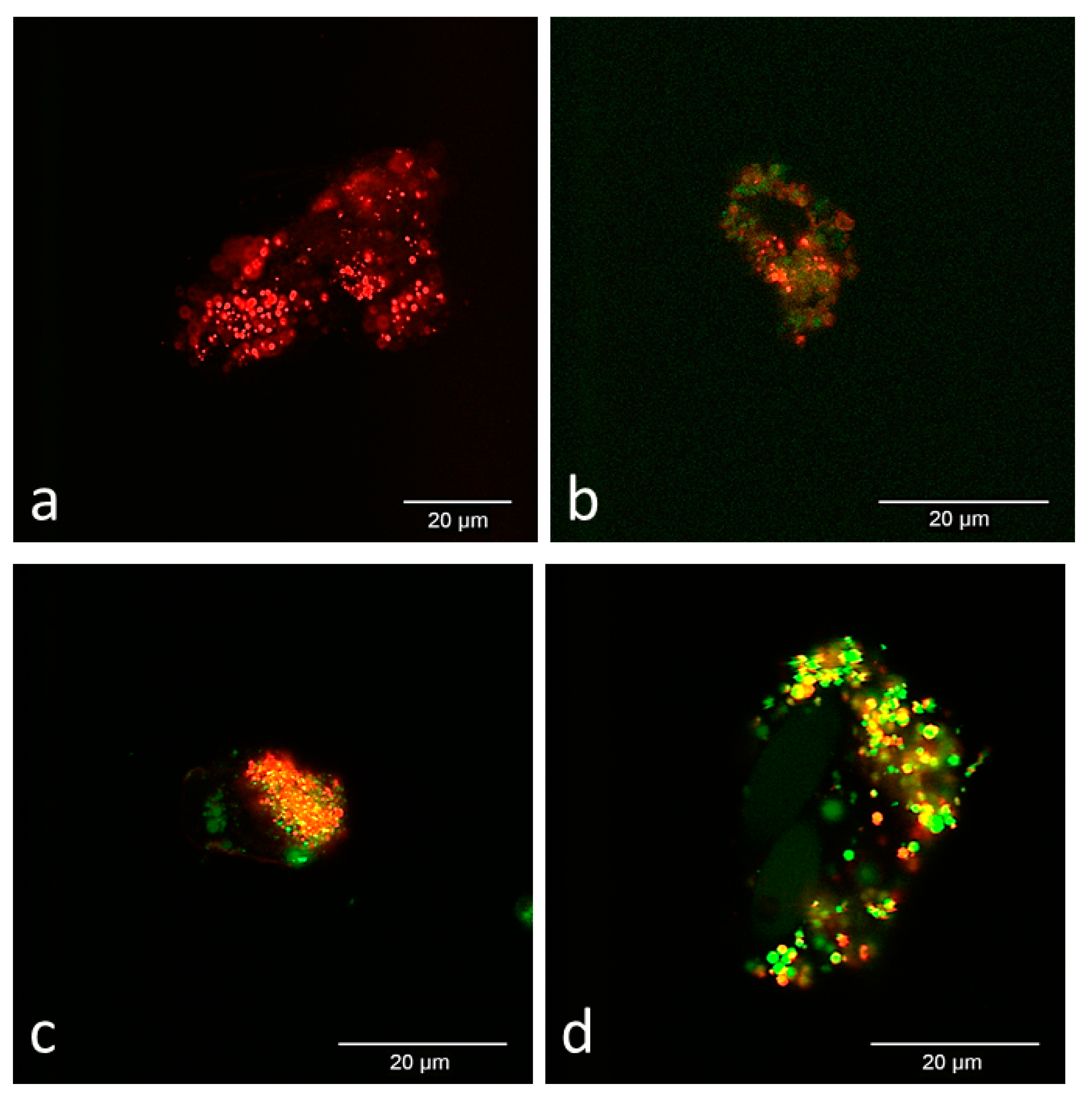
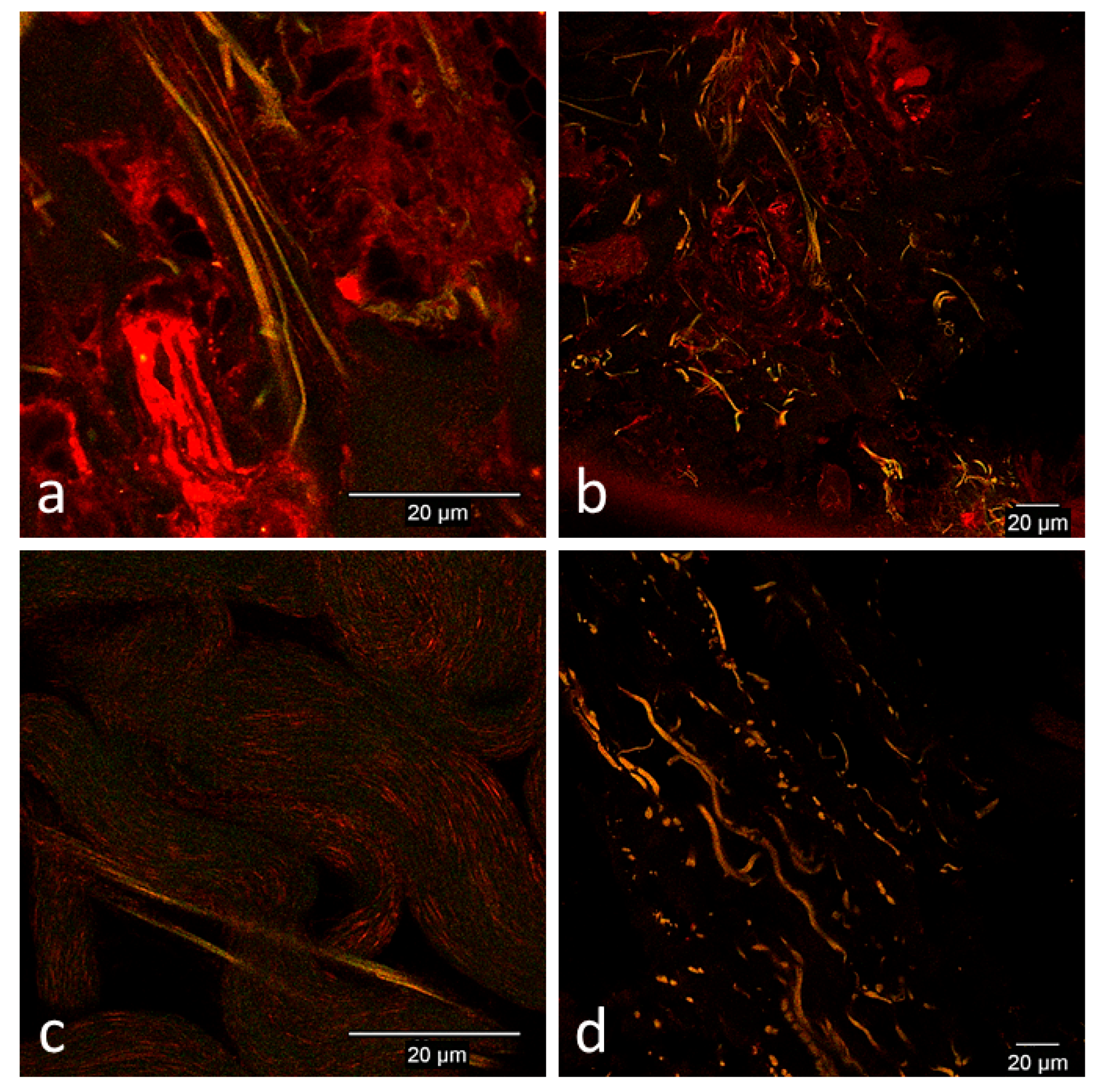
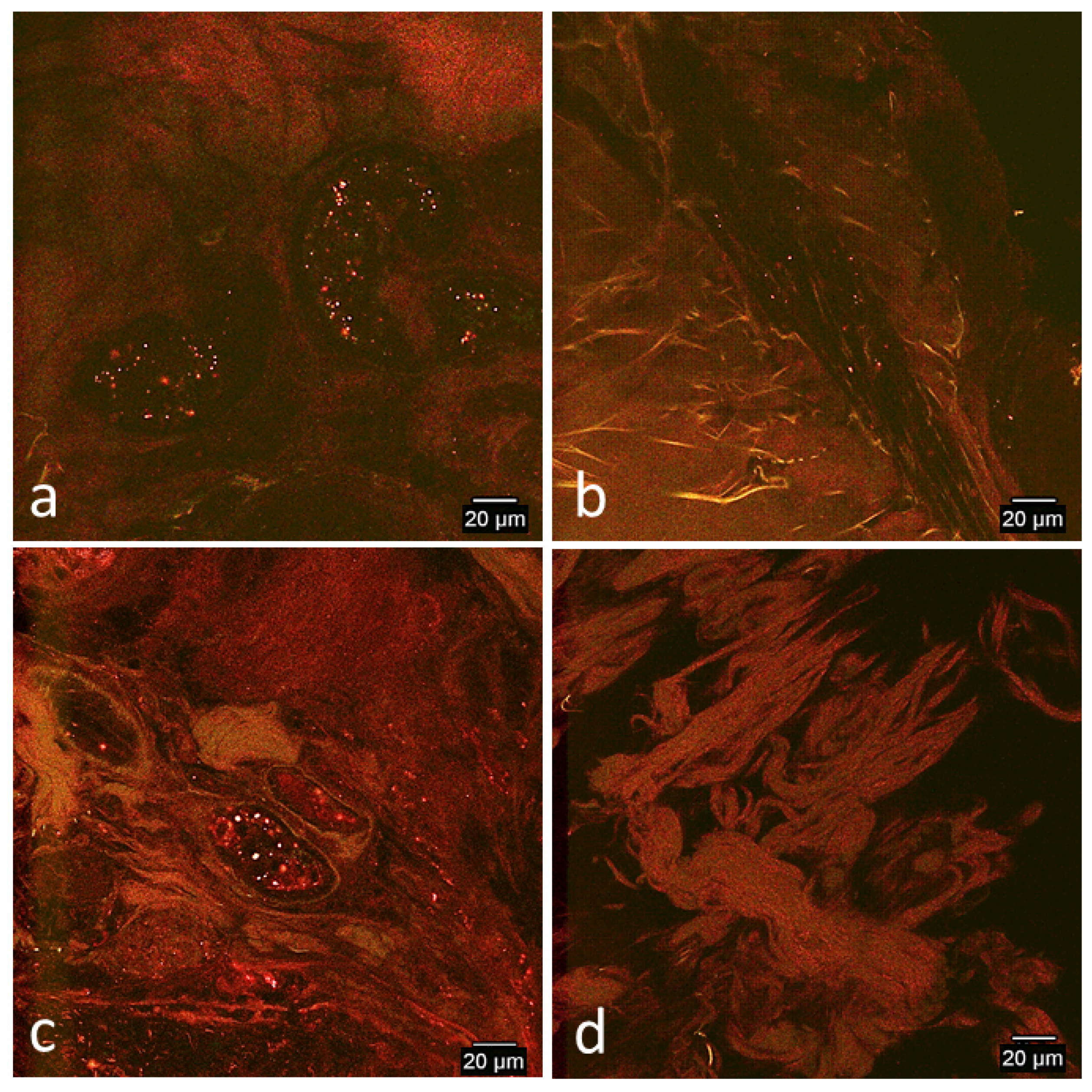
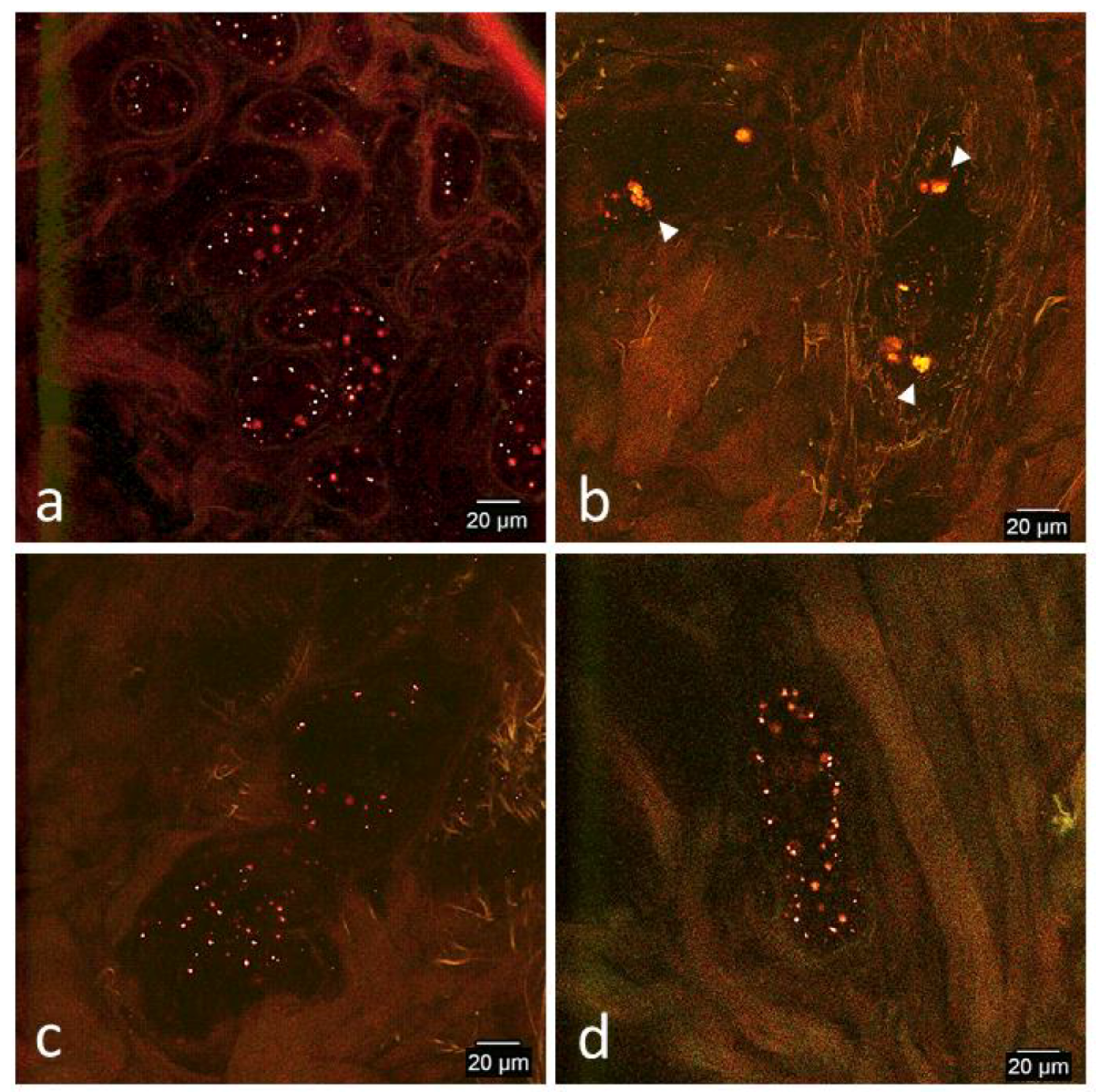
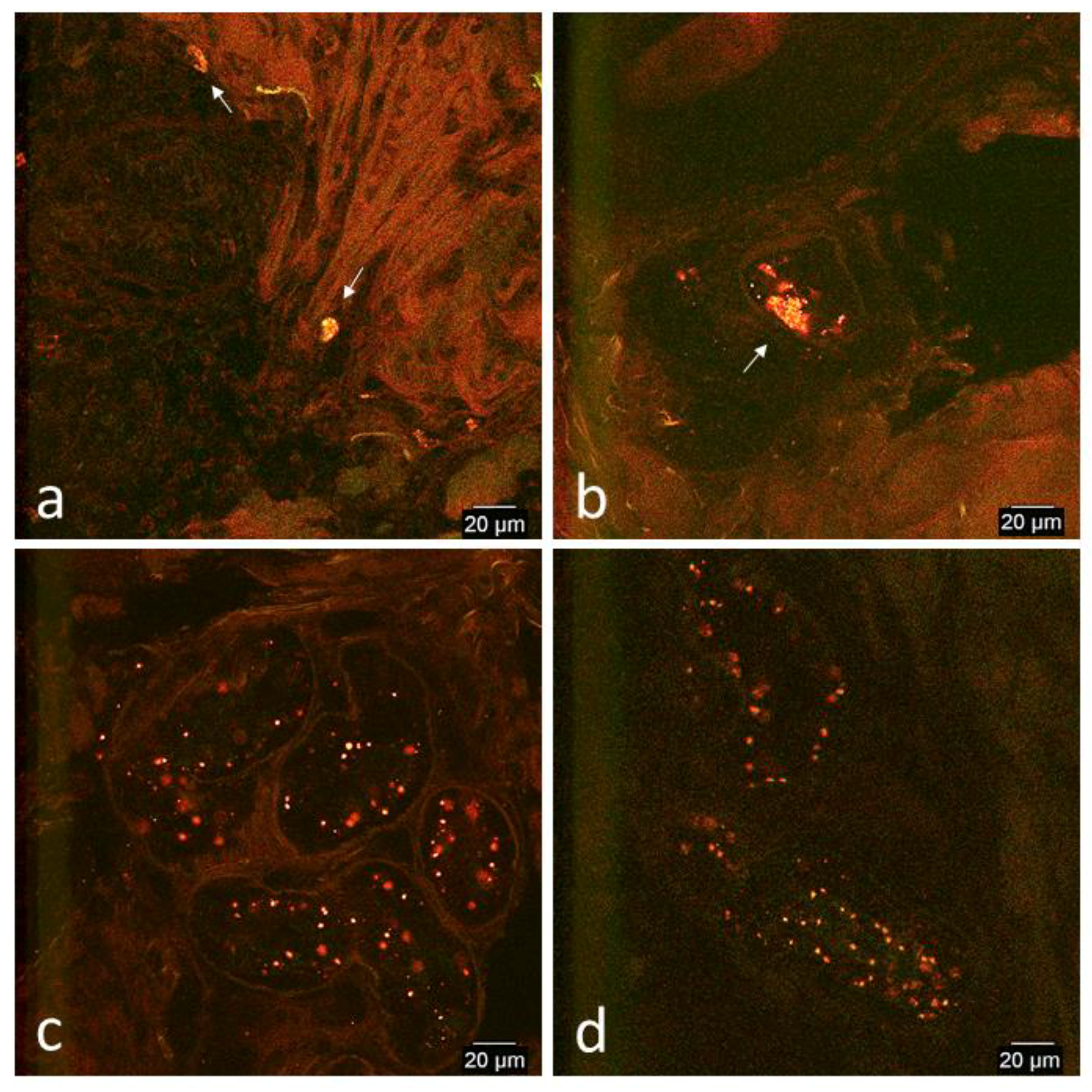
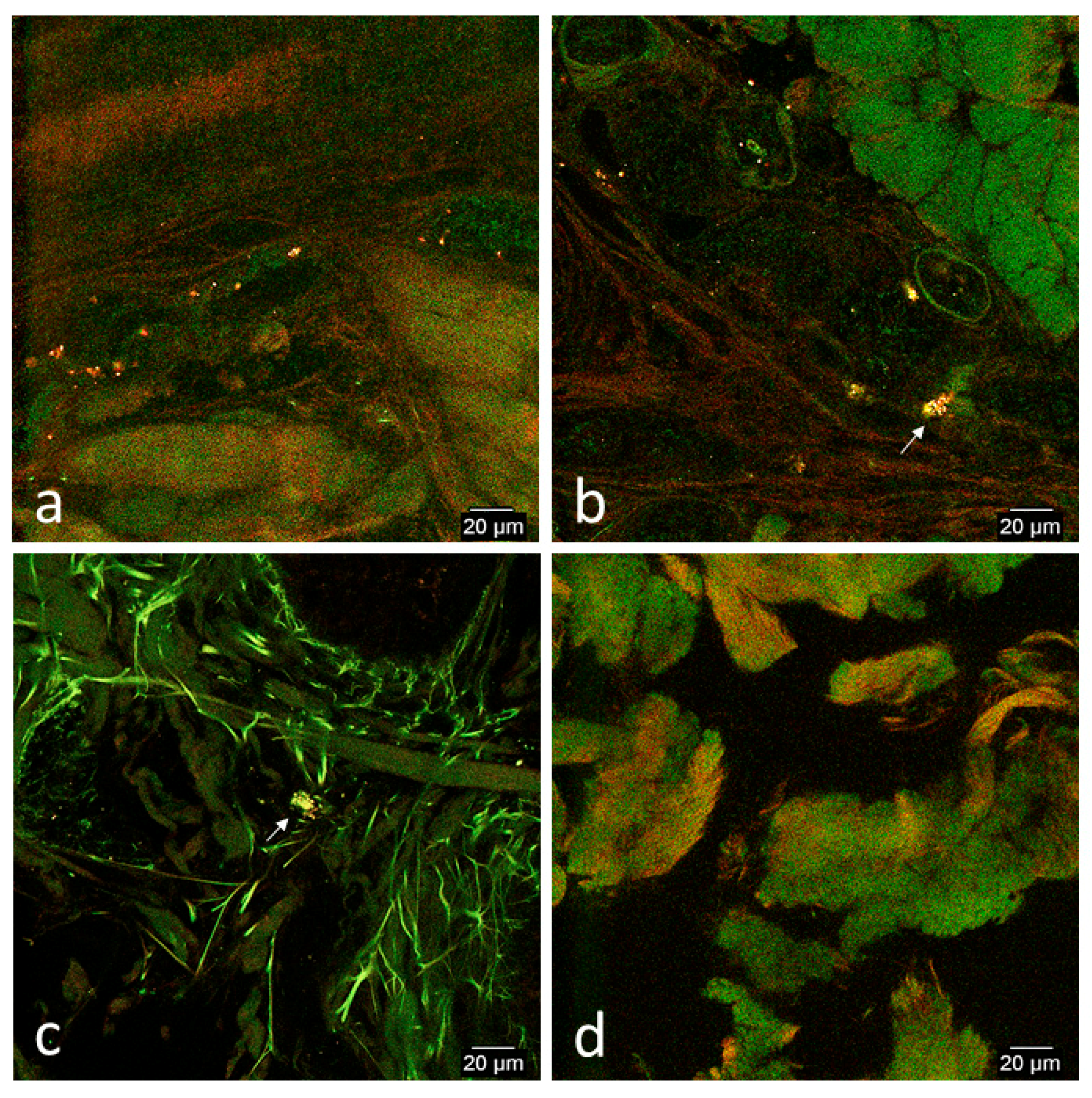
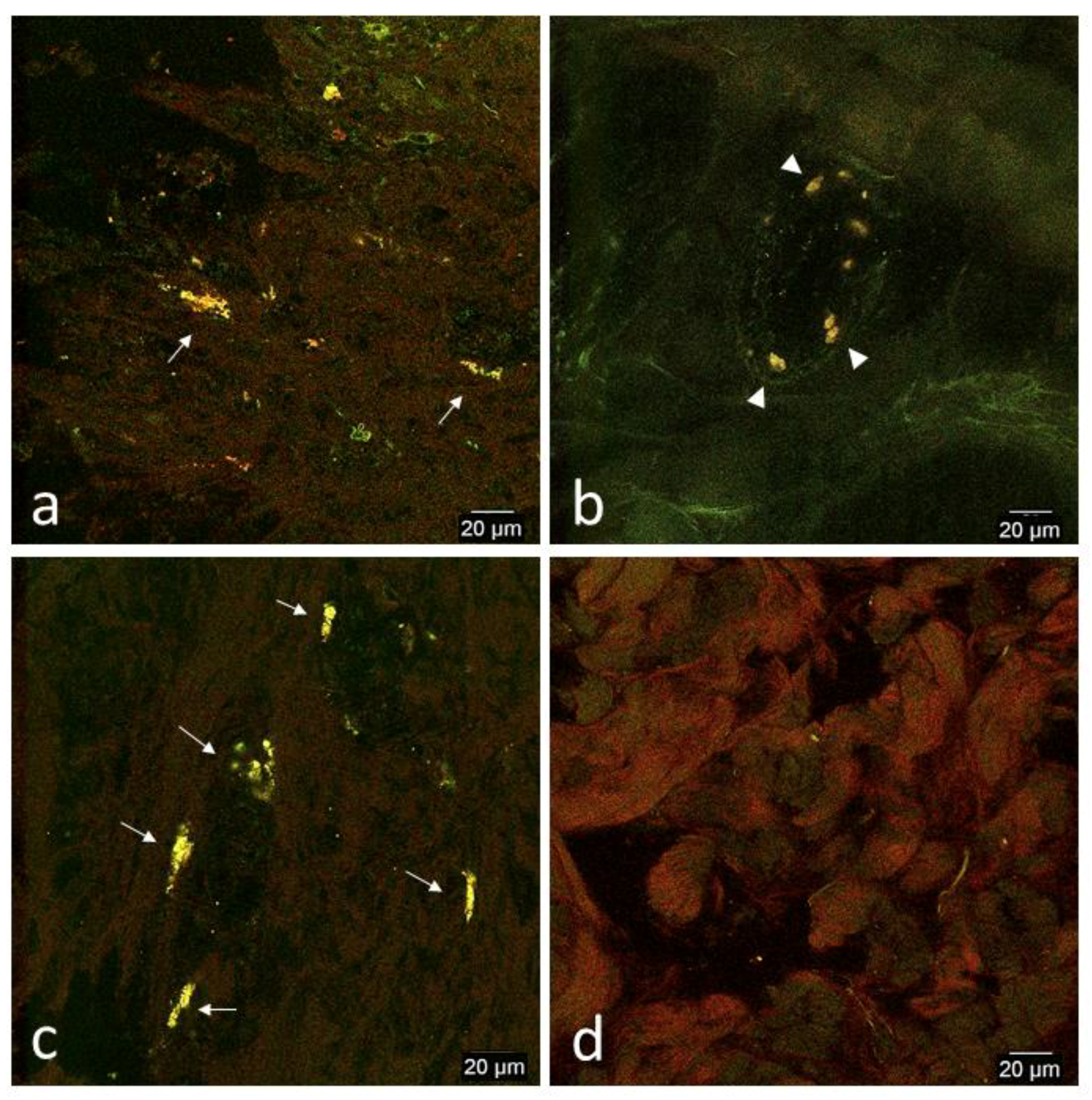
3.2. Confocal Microscopy Observations
3.2.1. In Vitro Culture
3.2.2. In Vivo Tissue Biopsies
Day 0
Day 1
Day 2
Day 7
Day 14
Day 33
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Theoret, C.; Wilmink, J.M. Exuberant granulation tissue. In Equine Wound Management; Theoret, C., Schumacher, J., Eds.; John Wiley & Sons, Inc.: Ames, IA, USA, 2017; pp. 369–384. [Google Scholar]
- Wilmink, J.M. Differences in wound healing between horses and ponies. In Equine Wound Management; Theoret, C., Schumacher, J., Eds.; John Wiley & Sons, Inc.: Ames, IA, USA, 2017; pp. 14–29. [Google Scholar]
- Theoret, C.L.; Wilmink, J.M. Aberrant wound healing in the horse: Naturally occurring conditions reminiscent of those observed in man. Wound Rep. Regener. 2013, 21, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Watt, S.M.; Pleat, J.M. Stem cells, niches and scaffolds: Applications to burns and wound care. Adv. Drug Deliv. Rev. 2017, 123, 82–106. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jang, Y.J. Recent understandings of biology, prophylaxis and treatment strategies for hypertrophic scars and keloids. Int. J. Mol. Sci. 2018, 19, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, C.-Y.; Biswas, A.; Subramanian, A.; Srinivasan, A.; Choolani, M.; Bongso, A. Human keloid cell characterization and inhibition of growth with human Wharton’s jelly stem cell extracts. J. Cell. Biochem. 2014, 115, 826–838. [Google Scholar] [CrossRef] [PubMed]
- Maxson, S.; Lopez, E.A.; Yoo, D.; Danilkovitch-Miagkova, A.; LeRoux, M.A. Concise review: Role of mesenchymal stem cells in wound repair. Stem Cells Transl. Med. 2012, 1, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Przyborowski, M.; Berthiaume, F. Stem cells for skin tissue engineering and wound healing. Crit. Rev. Biomed. Eng. 2009, 37, 399–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustad, K.C.; Gurtner, G.C. Mesenchymal stem cells home to sites of injury and inflammation. Adv. Wound Care 2012, 1, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Kean, T.J.; Lin, P.; Caplan, A.I.; Dennis, J.E.; Kean, J. MSCs: Delivery routes and engraftment, cell-targeting strategies, and immune modulation. Stem Cells Int. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ezquer, F.E.; Ezquer, M.E.; Vicencio, J.M.; Calligaris, S.D. Two complementary strategies to improve cell engraftment in mesenchymal stem cell-based therapy: Increasing transplanted cell resistance and increasing tissue receptivity. Cell Adhes. Migr. 2017, 11, 110–119. [Google Scholar] [CrossRef]
- Textor, J.A.; Clark, K.C.; Walker, N.J.; Aristizobal, F.A.; Kol, A.; LeJeune, S.S.; Bledsoe, A.; Davidyan, A.; Gray, S.N.; Bohannon-Worsley, L.K.; et al. Allogeneic stem cells alter gene expression and improve healing of distal limb wounds in horses. Stem Cells Transl. Med. 2018, 7, 98–108. [Google Scholar] [CrossRef]
- Spaas, J.H.; Gomiero, C.; Broeckx, S.Y.; Van Hecke, L.; Maccatrozzo, L.; Martens, A.; Martinello, T.; Patruno, M. Wound-healing markers after autologous and allogeneic epithelial-like stem cell treatment. Cytotherapy 2016, 18, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Broeckx, S.Y.; Borena, B.M.; Van Hecke, L.; Chiers, K.; Maes, S.; Guest, D.J.; Meyer, E.; Duchateau, L.; Martens, A.; Spaas, J.H. Comparison of autologous versus allogeneic epithelial-like stem cell treatment in an in vivo equine skin wound model. J. Cytother. 2015, 17, 1434–1446. [Google Scholar] [CrossRef] [PubMed]
- Ortved, K.F. Regenerative medicine and rehabilitation for tendinous and ligamentous injuries in sport horses. Vet. Clin. North Am. - Equine Pract. 2018, 34, 359–373. [Google Scholar] [CrossRef] [PubMed]
- De Schauwer, C. Stem cell therapy in the horse: From laboratory to clinic. Vet. J. 2015, 203, 137. [Google Scholar] [CrossRef] [PubMed]
- White, S.V.; Czisch, C.E.; Han, M.H.; Plant, C.D.; Harvey, A.R.; Plant, G.W. Intravenous transplantation of mesenchymal progenitors distribute solely to the lungs and improve outcomes in cervical spinal cord injury. Stem Cells 2016, 34, 1812–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawata, Y.; Tsuchiya, A.; Seino, S.; Watanabe, Y.; Kojima, Y.; Ikarashi, S.; Tominaga, K.; Yokoyama, J.; Yamagiwa, S.; Terai, S. Early injection of human adipose tissue-derived mesenchymal stem cell after inflammation ameliorates dextran sulfate sodium-induced colitis in mice through the induction of M2 macrophages and regulatory T cells. Cell Tissue Res. 2019, 376, 257–271. [Google Scholar] [CrossRef]
- Macrin, D.; Joseph, J.P.; Pillai, A.A.; Devi, A. Eminent sources of adult mesenchymal stem cells and their therapeutic imminence. Stem Cell Rev. Rep. 2017, 13, 741–756. [Google Scholar] [CrossRef]
- Koch, T.G.; Thomsen, P.D.; Betts, D.H. Improved isolation protocol for equine cord blood-derived mesenchymal stromal cells. Cytotherapy 2009, 11, 443–447. [Google Scholar] [CrossRef]
- Williams, L.B.; Koenig, J.B.; Black, B.; Gibson, T.W.G.; Sharif, S.; Koch, T.G. Equine allogeneic umbilical cord blood derived mesenchymal stromal cells reduce synovial fluid nucleated cell count and induce mild self-limiting inflammation when evaluated in an lipopolysaccharide induced synovitis model. Equine Vet. J. 2016, 48, 619–625. [Google Scholar] [CrossRef]
- Tessier, L.; Bienzle, D.; Williams, L.B.; Koch, T.G. Phenotypic and immunomodulatory properties of equine cord blood-derived mesenchymal stromal cells. PLoS ONE 2015, 10, e0122954. [Google Scholar] [CrossRef]
- Millán-Rivero, J.E.; Martínez, C.M.; Romecín, P.A.; Aznar-Cervantes, S.D.; Carpes-Ruiz, M.; Cenis, J.L.; Moraleda, J.M.; Atucha, N.M.; García-Bernal, D. Silk fibroin scaffolds seeded with Wharton’s jelly mesenchymal stem cells enhance re-epithelialization and reduce formation of scar tissue after cutaneous wound healing. Stem Cell Res. Ther. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, M.; Abe, R.; Fujita, Y.; Ando, S.; Inokuma, D.; Shimizu, H. Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J. Immunol. 2008, 180, 2581–2587. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, J.; Scott, P.G.; Tredget, E.E. Bone marrow-derived stem cells in wound healing: A review. Wound Repair Regen. 2007, 15, S18–S26. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yu, Y.; Hou, Y.; Chai, J.; Duan, H.; Chu, W.; Zhang, H.; Hu, Q.; Du, J. Human umbilical cord mesenchymal stem cells transplantation promotes cutaneous wound healing of severe burned rats. PLoS ONE 2014, 9, e88348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, D.D.; Singer, A.J.; Gordon, C.; Brink, P. The effects of rat mesenchymal stem cells on injury progression in a rat model. Acad. Emerg. Med. 2013, 20, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Trela, J.M.; Spriet, M.; Padgett, K.A.; Galuppo, L.D.; Vaughan, B.; Vidal, M.A. Scintigraphic comparison of intra-arterial injection and distal intravenous regional limb perfusion for administration of mesenchymal stem cells to the equine foot. Equine Vet. J. 2014, 46, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, P.; Spriet, M.; Sole, A.; Walker, N.J.; Vaughan, B.; Galuppo, L.D. Scintigraphic Tracking of Allogeneic Mesenchymal Stem Cells in the Distal Limb After Intra-Arterial Injection in Standing Horses. Vet. Surg. 2016, 45, 619–624. [Google Scholar] [CrossRef]
- Spriet, M.; Trela, J.M.; Galuppo, L.D. Ultrasound-guided injection of the median artery in the standing sedated horse. Equine Vet. J. 2014, 1–4. [Google Scholar] [CrossRef]
- Furlani, D.; Ugurlucan, M.; Ong, L.; Bieback, K.; Pittermann, E.; Westien, I.; Wang, W.; Yerebakan, C.; Li, W.; Gaebel, R.; et al. Is the intravascular administration of mesenchymal stem cells safe? Mesenchymal stem cells and intravital microscopy. Microvasc. Res. 2009, 77, 370–376. [Google Scholar] [CrossRef]
- Kang, M.H.; Park, H.M. Evaluation of adverse reactions in dogs following intravenous mesenchymal stem cell transplantation. Acta Vet. Scand. 2014, 56, 16. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.L.; Kerkelä, E.; Bakreen, A.; Nitzsche, F.; Andrzejewska, A.; Nowakowski, A.; Janowski, M.; Walczak, P.; Boltze, J.; Lukomska, B.; et al. The cerebral embolism evoked by intra-arterial delivery of allogeneic bone marrow mesenchymal stem cells in rats is related to cell dose and infusion velocity. Stem Cell Res. Ther. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzanite, L.M.; Fortier, L.A.; Antczak, D.F.; Cassano, J.M.; Brosnahan, M.M.; Miller, D.; Schnabel, L.V. Equine allogeneic bone marrow-derived mesenchymal stromal cells elicit antibody responses in vivo. Stem Cell Res. Ther. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broeckx, S.; Borena, B.M.; Zimmerman, M.; Marien, T.; Serys, B.; Suls, M.; Duchateau, L.; Spaas, J.H. Intravenous application of allogeneic peripheral blood-derived mesenchymal stem cells: A safety assessment in 291 equine recipients. Curr. Stem Cell Res. Ther. 2014, 9, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Kol, A.; Wood, J.A.; Carrade Holt, D.D.; Gillette, J.A.; Bohannon-Worsley, L.K.; Puchalski, S.M.; Walker, N.J.; Clark, K.C.; Watson, J.L.; Borjesson, D.L. Multiple intravenous injections of allogeneic equine mesenchymal stem cells do not induce a systemic inflammatory response but do alter lymphocyte subsets in healthy horses. Stem Cell Res. Ther. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, S.D.; Kol, A.; Walker, N.J.; Borjesson, D.L. Allogeneic mesenchymal stem cell treatment induces specific alloantibodies in horses. Stem Cells Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.B.; Co, C.; Koenig, J.B.; Tse, C.; Lindsay, E.; Koch, T.G. Response to intravenous allogeneic equine cord blood-derived mesenchymal stromal cells administered from chilled or frozen state in serum and protein-free media. Front. Vet. Sci. 2016, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Honaramooz, A.; Megee, S.; Zeng, W.; Destrempes, M.M.; Overton, S.A.; Luo, J.; Galantino-Homer, H.; Modelski, M.; Chen, F.; Blash, S.; et al. Adeno-associated virus (AAV)-mediated transduction of male germ line stem cells results in transgene transmission after germ cell transplantation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 374–382. [Google Scholar] [CrossRef]
- Zeng, W.; Tang, L.; Bondareva, A.; Honaramooz, A.; Tanco, V.; Dores, C.; Megee, S.; Modelski, M.; Rodriguez-sosa, J.R.; Paczkowski, M.; et al. Viral transduction of male germline stem cells results in transgene transmission after germ cell transplantation in pigs viral transduction of male germline stem cells results in transgene transmission after germ cell transplantation in pigs. Biol. Reprod. 2013, 88, 1–9. [Google Scholar] [CrossRef]
- Honaramooz, A.; Megee, S.O.; Dobrinski, I. Germ Cell Transplantation in Pigs. Biol. Reprod. 2002, 66, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Honaramooz, A.; Behboodi, E.; Blash, S.; Megee, S.O.; Dobrinski, I. Germ cell transplantation in goats. Mol. Reprod. Dev. 2003, 64, 422–428. [Google Scholar] [CrossRef]
- Theoret, C.L.; Barber, S.M.; Gordon, J.R. Temporal localization of immunoreactive transforming growth factor B1 in normal equine skin and full-thickness dermal wounds. Vet. Surg. 2002, 31, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Theoret, C.L.; Moyana, T.N.; Barber, S.M.; Gordon, J.R. Preliminary observations on expression of transforming growth factors B1 and B3 in equine full-thickness skin wounds healing normally or with exuberant granulation tissue. Vet. Surg. 2002, 31, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Theoret, C.L.; Barber, S.M.; Moyana, T.N.; Gordon, J.R. Expression of transforming growth factor B1, B3, and basic fibroblast growth factor in full-thickness skin wounds of equine limbs and thorax. Vet. Surg. 2001, 30, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Wilmink, J.M.; Stolk, P.W.T.; Van Weeran, P.R.; Barneveld, A. Differences in second-intention wound healing between horses and ponies: Histological aspects. Equine Vet. J. 1999, 31, 61–67. [Google Scholar] [CrossRef]
- Wilmink, J.M.; Stolk, P.W.T.; Van Weeran, P.R.; Barneveld, A. Differences in second-intention wound healing between horses and ponies: Macroscopic aspects. Equine Vet. J. 1999, 31, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Wilmink, J.M.; Veenman, J.N.; Van den Boom, R.; Rutten, V.P.M.G.; Niewold, T.A.; Broekhuisen-Davies, J.M.; Lees, P.; Armstrong, S.; Van Weeren, P.R.; Barneveld, A. Differences in polymorphonucleocyte function and local inflammatory response between horses and ponies. Equine Vet. J. 2003, 35, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Van den Boom, R.; Wilmink, J.M.; O’Kane, S.; Wood, J.; Ferguson, M.W.J. Transforming growth factor-beta levels during second- intention healing are related to the different course of wound contraction in horses and ponies. Wound Repair Regen. 2002, 10, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Celeste, C.J.; Deschene, K.; Riley, C.B.; Theoret, C.L. Regional differences in wound oxygenation during normal healing in an equine model of cutaneous fibroproliferative disorder. Wound Repair Regen. 2011, 19, 89–97. [Google Scholar] [CrossRef]
- Theoret, C.L.; Olutoye, O.O.; Parnell, L.K.S.; Hicks, J. Equine exuberant granulation tissue and human keloids: A comparative histopathologic study. Vet. Surg. 2013, 42, 783–789. [Google Scholar] [CrossRef]
- Huang, C.; Murphy, G.F.; Akaishi, S.; Ogawa, R. Keloids and hypertrophic scars. Plast. Reconstr. Surg. Glob. Open 2013, 1, e25. [Google Scholar] [CrossRef]
- Shaker, S.A.; Ayuob, N.N.; Hajrah, N.H. Cell talk: A phenomenon observed in the keloid scar by immunohistochemical study. Appl. Immunohistochem. Mol. Morphol. 2011, 19, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Kanji, S.; Das, H. Advances of stem cell therapeutics in cutaneous wound healing and regeneration. Mediat. Inflamm. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanci, A.; Merlo, B.; Mariella, J.; Castagnetti, C.; Iacono, E. Heterologous Wharton’s jelly derived mesenchymal stem cells application on a large chronic skin wound in a 6-month-old filly. Front. Vet. Sci. 2019, 6, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaas, J.H.; De Schauwer, C.; Cornillie, P.; Meyer, E.; Van Soom, A.; Van de Walle, G.R. Culture and characterisation of equine peripheral blood mesenchymal stromal cells. Vet. J. 2013, 195, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Iacono, E.; Merlo, B.; Pirrone, A.; Antonelli, C.; Brunori, L.; Romagnoli, N. Effects of mesenchymal stem cells isolated from amniotic fluid and platelet- rich plasma gel on severe decubitus ulcers in a septic neonatal foal. Res. Vet. Sci. 2012, 93, 1439–1440. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.J.; Jarazo, J. State of the art: Stem cells in equine regenerative medicine. Equine Vet. J. 2015, 47, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, Y.Z.; Rash, N.; Garvican, E.R.; Paillot, R.; Guest, D.J. Equine mesenchymal stromal cells and embryo-derived stem cells are immune privileged in vitro. Stem Cell Res. Ther. 2014, 5, 90. [Google Scholar] [CrossRef] [Green Version]
- Ranera, B.; Lyahyai, J.; Romero, A.; Vázquez, F.J.; Remacha, A.R.; Bernal, M.L.; Zaragoza, P.; Rodellar, C.; Martín-Burriel, I. Immunophenotype and gene expression profiles of cell surface markers of mesenchymal stem cells derived from equine bone marrow and adipose tissue. Vet. Immunol. Immunopathol. 2011, 144, 147–154. [Google Scholar] [CrossRef]
- Williams, L.B.; Tessier, L.; Koenig, J.B.; Koch, T.G. Post-thaw non-cultured and post-thaw cultured equine cord blood mesenchymal stromal cells equally suppress lymphocyte proliferation in vitro. PLoS ONE 2014, 9, e113615. [Google Scholar] [CrossRef]
- Colbath, A.C.; Dow, S.W.; Phillips, J.N.; McIlwraith, W.C.; Goodrich, L.R. Autologous and allogeneic equine mesenchymal stem cells exhibit equivalent immunomodulatory properties in vitro. Stem Cells Dev. 2017, 26, 503–511. [Google Scholar] [CrossRef]
- Barrachina, L.; Remacha, A.R.; Romero, A.; Vázquez, F.J.; Albareda, J.; Prades, M.; Ranera, B.; Zaragoza, P.; Martin-Burriel, I.; Rodellar, C. Effect of inflammatory environment on equine bone marrow derived mesenchymal stem cells immunogenicity and immunomodulatory properties. Vet. Immunol. Immunopathol. 2016, 171, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Remacha, A.R.; Barrachina, L.; Álvarez-Arguedas, S.; Ranera, B.; Romero, A.; Vazquez, F.J.; Zaragoza, P.; Yanez, R.; Martín-Burriel, I.; Rodellar, C. Expression of genes involved in immune response and in vitro immunosuppressive effect of equine MSCs. Vet. Immunol. Immunopathol. 2015, 165, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Berglund, A.K.; Schnabel, L.V. Allogeneic major histocompatibility complex-mismatched equine bone marrow-derived mesenchymal stem cells are targeted for death by cytotoxic anti-major histocompatibility complex antibodies. Equine Vet. J. 2017, 49, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, C.; Theoret, C.; Smith, O.; Segura, M.; Lemire, P.; Smith, L.C. Immune potential of allogeneic equine induced pluripotent stem cells. Equine Vet. J. 2015, 47, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Joswig, A.J.; Mitchell, A.; Cummings, K.J.; Levine, G.J.; Gregory, C.A.; Smith, R.; Watts, A.E. Repeated intra-articular injection of allogeneic mesenchymal stem cells causes an adverse response compared to autologous cells in the equine model. Stem Cell Res. Ther. 2017, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigg, A.; Gibson, R.; Bardy, P.; Szer, J. Acute portal vein thrombosis after autologous stem cell transplantation. Bone Marrow Transplant. 1996, 18, 949–953. [Google Scholar]
- Ra, J.C.; Shin, I.S.; Kim, S.H.; Kang, S.K.; Kang, B.C.; Lee, H.Y.; Kim, Y.J.; Jo, J.Y.; Yoon, E.J.; Choi, H.J.; et al. Safety of intravenous infusion of human adipose tissue-derived mesenchymal stem cells in animals and humans. Stem Cells Dev. 2011, 20, 1297–1308. [Google Scholar] [CrossRef]
- Kim, Y.; Jo, S.; Kim, W.H.; Kweon, O.-K. Antioxidant and anti-inflammatory effects of intravenously injected adipose derived mesenchymal stem cells in dogs with acute spinal cord injury. Stem Cell Res. Ther. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.-H.; Zhang, T.-T.; Li, Y.; Yan, H.-J.; Qi, H.; Li, F.-R. Immunogenicity of allogeneic mesenchymal stem cells transplanted via different routes in diabetic rats. Cell. Mol. Immunol. 2015, 444–455. [Google Scholar] [CrossRef] [Green Version]
- Kabat, M.; Bobkov, I.; Kumar, S.; Grumet, M. Trends in mesenchymal stem cell clinical trials 2004–2018: Is efficacy optimal in a narrow dose range? Stem Cells Transl. Med. 2020, 9, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Conway, A.; Vazin, T.; Spelke, D.P.; Rode, N.A.; Healy, K.E.; Kane, R.S.; Schaffer, D.V. Multivalent ligands control stem cell behaviour in vitro and in vivo. Nat. Nanotechnol. 2013, 8, 831–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Nitzsche, F.; Müller, C.; Lukomska, B.; Jolkkonen, J.; Deten, A.; Boltze, J. The MSC adhesion cascade—Insights into homing and transendothelial migration. Stem Cells 2017, 35, 1446–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.; Xu, L.; Zwingenberger, S.; Gibon, E.; Goodman, S.B.; Li, G. Mesenchymal stem cells homing to improve bone healing. J. Orthop. Transl. 2017, 9, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.É.S.; Sousa, M.R.R.; Alencar-Silva, T.; Carvalho, J.L.; Saldanha-Araujo, F. Mesenchymal stem cells immunomodulation: The road to IFN-γ licensing and the path ahead. Cytokine Growth Factor Rev. 2019, 47, 32–42. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, L.; Scott, P.G.; Tredget, E.E. Mesenchymal stem cells enhance wound healing through differentiation and angiogenesis. Stem Cells (Dayt. Ohio) 2007, 25, 2648–2659. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Luo, X.; Lv, X.; Liu, V.; Zhao, G.; Zhang, X.; Cao, W.; Wang, R.; Wang, W. In vivo human adipose-derived mesenchymal stem cell tracking after intra-articular delivery in a rat osteoarthritis model. Stem Cell Res. Ther. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Guest, D.J.; Smith, M.R.W.; Allen, W.R. Equine embryonic stem-like cells and mesenchymal stromal cells have different survival rates and migration patterns following their injection into damaged superficial digital flexor tendon. Equine Vet. J. 2010, 42, 636–642. [Google Scholar] [CrossRef]
- Shin, L.; Peterson, D.A. Human mesenchymal stem cell grafts enhance normal and impaired wound healing by recruiting existing endogenous tissue stem/progenitor cells. Stem Cells Transl. Med. 2013, 2, 33–42. [Google Scholar] [CrossRef]
- Othman, O.E.; Mahrous, K.F.; Shafey, H.I. Mitochondrial DNA genetic variations among four horse populations in Egypt. J. Genet. Eng. Biotechnol. 2017, 15, 469–474. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biopsy Collection Day | Day 1 | Day 2 | Day 7 | Day 14 | Day 33 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Subject | Biopsy Site | Color | Homing | Color | Homing | Color | Homing | Color | Homing | Color | Homing |
| Horse 1 | Limb Wounded | R, G | 1+ | R, G | 1+ | R, G | = | R, G | = | R, G | 3+ |
| Limb Non-Wounded | R, G | R, G | R, G | R, G | R, G | ||||||
| Thorax Wounded | R, G | 1+ | R, G | = | R, G | = | R, G | = | R, G | 3+ | |
| Thorax Non-Wounded | none | R, G | R, G | R, G | R, G | ||||||
| Horse 2 | Limb Wounded | R, G | 1+ | R, G | 1+ | R, G | = | R, G | = | R, G | 3+ |
| Limb Non-Wounded | R, G | R, G | R, G | R, G | R, G | ||||||
| Thorax Wounded | R, G | 1+ | R, G | = | R, G | = | R, G | = | R, G | 3+ | |
| Thorax Non-Wounded | R, G | R, G | R, G | R, G | R, G | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mund, S.J.K.; Kawamura, E.; Awang-Junaidi, A.H.; Campbell, J.; Wobeser, B.; MacPhee, D.J.; Honaramooz, A.; Barber, S. Homing and Engraftment of Intravenously Administered Equine Cord Blood-Derived Multipotent Mesenchymal Stromal Cells to Surgically Created Cutaneous Wound in Horses: A Pilot Project. Cells 2020, 9, 1162. https://doi.org/10.3390/cells9051162
Mund SJK, Kawamura E, Awang-Junaidi AH, Campbell J, Wobeser B, MacPhee DJ, Honaramooz A, Barber S. Homing and Engraftment of Intravenously Administered Equine Cord Blood-Derived Multipotent Mesenchymal Stromal Cells to Surgically Created Cutaneous Wound in Horses: A Pilot Project. Cells. 2020; 9(5):1162. https://doi.org/10.3390/cells9051162
Chicago/Turabian StyleMund, Suzanne J. K., Eiko Kawamura, Awang Hazmi Awang-Junaidi, John Campbell, Bruce Wobeser, Daniel J. MacPhee, Ali Honaramooz, and Spencer Barber. 2020. "Homing and Engraftment of Intravenously Administered Equine Cord Blood-Derived Multipotent Mesenchymal Stromal Cells to Surgically Created Cutaneous Wound in Horses: A Pilot Project" Cells 9, no. 5: 1162. https://doi.org/10.3390/cells9051162
APA StyleMund, S. J. K., Kawamura, E., Awang-Junaidi, A. H., Campbell, J., Wobeser, B., MacPhee, D. J., Honaramooz, A., & Barber, S. (2020). Homing and Engraftment of Intravenously Administered Equine Cord Blood-Derived Multipotent Mesenchymal Stromal Cells to Surgically Created Cutaneous Wound in Horses: A Pilot Project. Cells, 9(5), 1162. https://doi.org/10.3390/cells9051162