The Paracrine Role of Endothelial Cells in Bone Formation via CXCR4/SDF-1 Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation, Expansion and Characterization of EPC
2.2. HNDF Culture
2.3. MSCs Culture
2.4. DII Staining
2.5. In Vivo Ectopic Bone Model
2.6. Scaffold Preparation
2.7. Subcutaneous Cell Transplantation
2.8. Dextran Preparation and Injection
2.9. Histological Preparation
2.10. Immunohistochemistry
2.11. Double Staining Immunohistochemistry
2.12. EPC Conditioned Medium (EPC-CM) Preparation
2.13. RT-PCR
2.14. Migration Assay
2.15. Statistical Analysis
3. Results
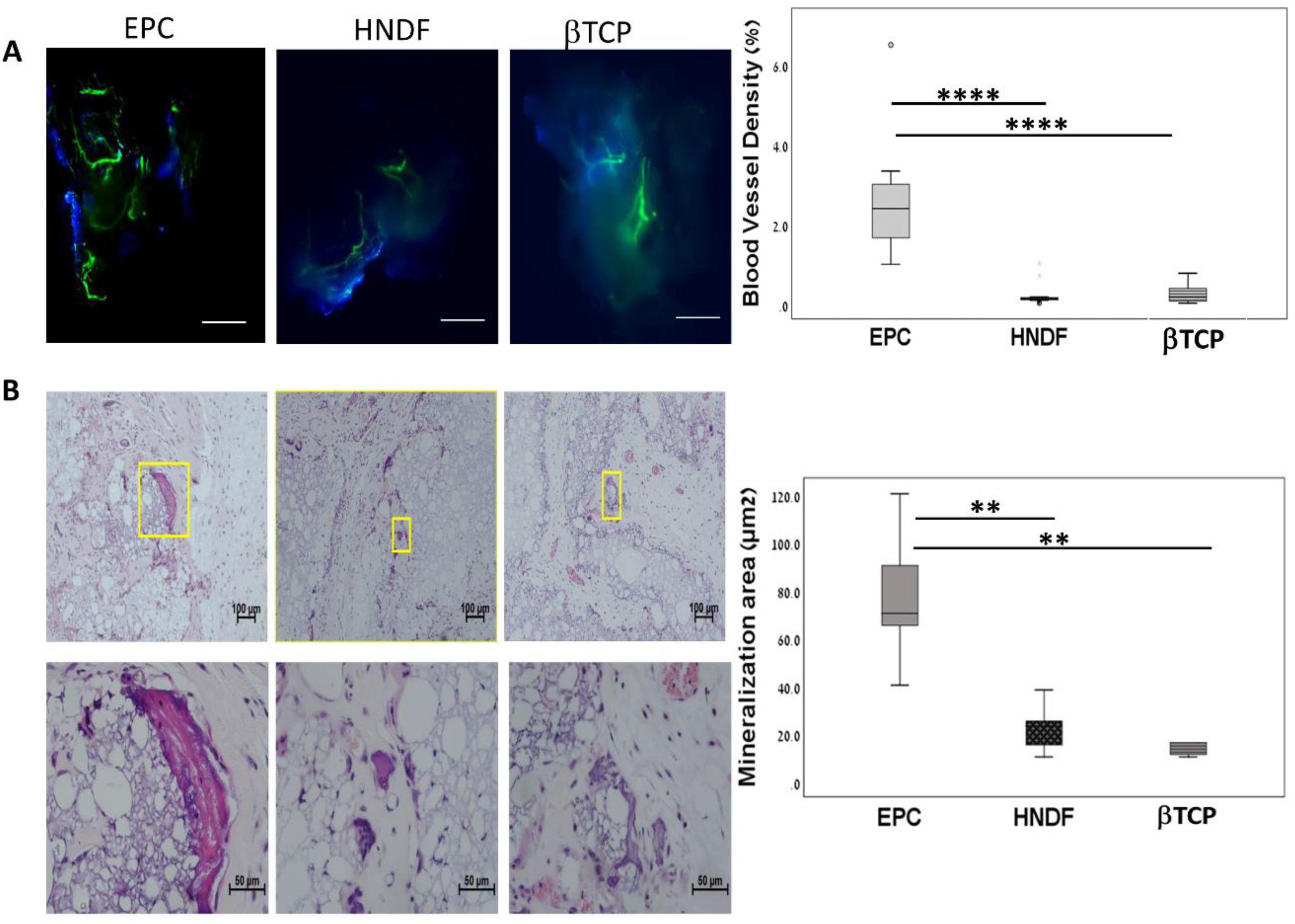
3.1. Human EPCs Enhance Angiogenesis and Mineralization in An Ectopic Bone Model
3.2. EPCs into Newly Formed Vessels in the Ectopic Transplants
3.3. EPCs Recruit Endogenous CD31 and CD73 Cells in Vivo
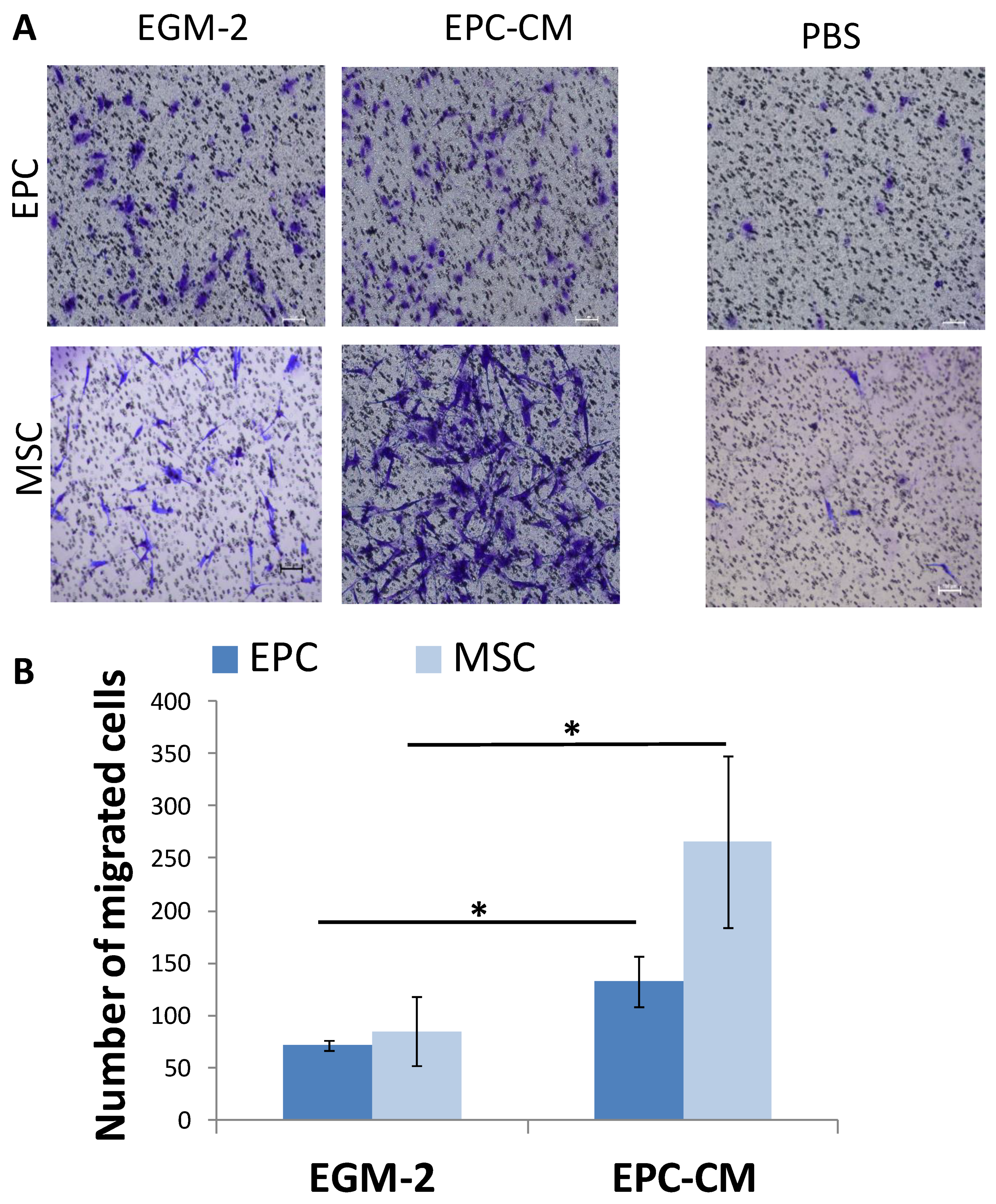
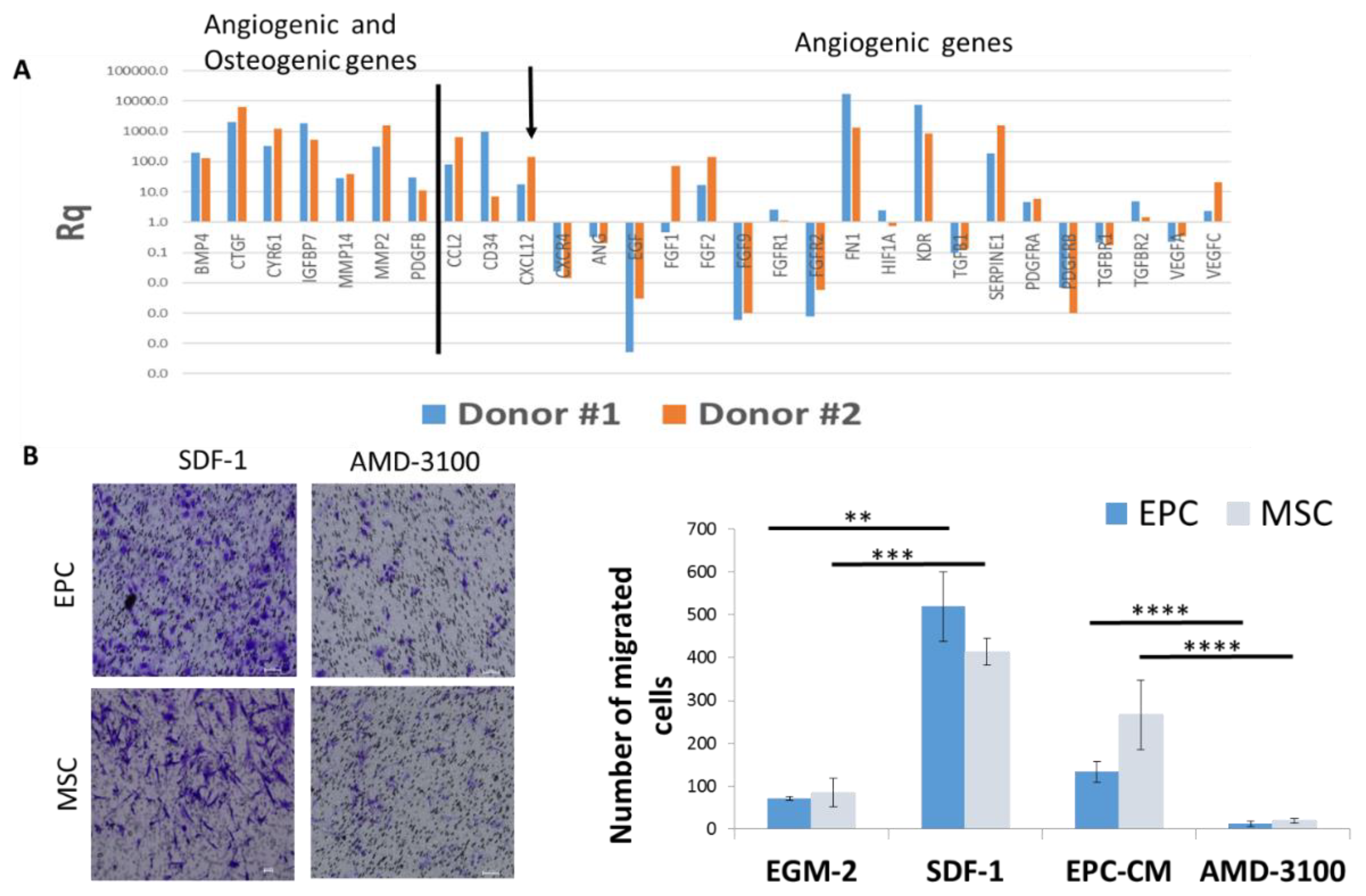
3.4. SDF-1 in EPC Conditioned Medium (EPC-CM) Stimulate Migration of EPCs and MSCs In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Streeten, E.A.; Brandi, M.L. Biology of bone endothelial cells. Bone Miner. 1990, 10, 85–94. [Google Scholar] [CrossRef]
- Collin-Osdoby, P. Role of vascular endothelial cells in bone biology. J. Cell. Biochem. 1994, 55, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Sivaraj, K.K.; Adams, R.H. Blood vessel formation and function in bone. Development 2016, 143, 2706–2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calori, G.M.; Giannoudis, P.V. Enhancement of fracture healing with the diamond concept: The role of the biological chamber. Injury 2011, 42, 1191–1193. [Google Scholar] [CrossRef] [PubMed]
- Motamedian, S.R.; Khojaste, M.; Khojasteh, A. Success rate of implants placed in autogenous bone blocks versus allogenic bone blocks: A systematic literature review. Ann. Maxillofac. Surg. 2016, 6, 78–90. [Google Scholar] [PubMed] [Green Version]
- Pandit, N.; Pandit, I.K.; Malik, R.; Bali, D.; Jindal, S. Autogenous bone block in the treatment of teeth with hopeless prognosis. Contemp. Clin. Dent. 2012, 3, 437–442. [Google Scholar] [CrossRef]
- Janicki, P.; Schmidmaier, G. What should be the characteristics of the ideal bone graft substitute? Combining scaffolds with growth factors and/or stem cells. Injury 2011, 42 (Suppl. 2), S77–S81. [Google Scholar] [CrossRef]
- Giannoudis, P.V.; Dinopoulos, H.; Tsiridis, E. Bone substitutes: An update. Injury 2005, 36 (Suppl. 3), S20–S27. [Google Scholar] [CrossRef]
- Marquez-Curtis, L.A.; Janowska-Wieczorek, A. Enhancing the migration ability of mesenchymal stromal cells by targeting the SDF-1/CXCR4 axis. Biomed. Res. Int. 2013, 2013, 561098. [Google Scholar] [CrossRef] [Green Version]
- Najar, M.; Bouhtit, F.; Melki, R.; Afif, H.; Hamal, A.; Fahmi, H.; Merimi, M.; Lagneaux, L. Mesenchymal Stromal Cell-Based Therapy: New Perspectives and Challenges. J. Clin. Med. 2019, 8, 626. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.Y.; Cho, T.-J.; Kim, J.A.; Lee, H.R.; Yoo, W.J.; Chung, C.Y.; Choi, I.H. Mobilization of endothelial progenitor cells in fracture healing and distraction osteogenesis. Bone 2008, 42, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Schipani, E.; Maes, C.; Carmeliet, G.; Semenza, G.L. Regulation of Osteogenesis-Angiogenesis Coupling by HIFs and VEGF. J. Bone Miner. Res. 2009, 24, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Kalka, C.; Masuda, H.; Chen, D.; Silver, M.; Kearney, M.; Magner, M.; Isner, J.M.; Asahara, T. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat. Med. 1999, 5, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; Van Der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of Putative Progenitor Endothelial Cells for Angiogenesis. Science 1997, 275, 964–966. [Google Scholar] [CrossRef]
- Urbich, C.; Heeschen, C.; Aicher, A.; Dernbach, E.; Zeiher, A.M.; Dimmeler, S. Relevance of Monocytic Features for Neovascularization Capacity of Circulating Endothelial Progenitor Cells. Circulation 2003, 108, 2511–2516. [Google Scholar] [CrossRef] [Green Version]
- Kalka, C.; Masuda, H.; Takahashi, T.; Kalka-Moll, W.M.; Silver, M.; Kearney, M.; Li, T.; Isner, J.M.; Asahara, T. Transplantation of ex vivo expanded endothelial progenitor cells for therapeutic neovascularization. Proc. Natl. Acad. Sci. USA 2000, 97, 3422–3427. [Google Scholar]
- Hur, J.; Yoon, C.H.; Kim, H.S.; Choi, J.H.; Kang, H.J.; Hwang, K.K.; Oh, B.H.; Lee, M.M.; Park, Y.B. Characterization of Two Types of Endothelial Progenitor Cells and Their Different Contributions to Neovasculogenesis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 288–293. [Google Scholar] [CrossRef]
- Medina, R.J.; O’Neill, C.L.; Sweeney, M.; Guduric-Fuchs, J.; Gardiner, T.A.; Simpson, D.; Stitt, A.W. Molecular analysis of endothelial progenitor cell (EPC) subtypes reveals two distinct cell populations with different identities. BMC Med Genom. 2010, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Balaji, S.; King, A.; Crombleholme, T.M.; Keswani, S.G. The Role of Endothelial Progenitor Cells in Postnatal Vasculogenesis: Implications for Therapeutic Neovascularization and Wound Healing. Adv. Wound Care 2013, 2, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Rozen, N.; Bick, T.; Bajayo, A.; Shamian, B.; Schrift-Tzadok, M.; Gabet, Y.; Yayon, A.; Bab, I.; Soudry, M.; Lewinson, D. Transplanted blood-derived endothelial progenitor cells (EPC) enhance bridging of sheep tibia critical size defects. Bone 2009, 45, 918–924. [Google Scholar] [CrossRef]
- Zigdon-Giladi, H.; Michaeli-Geller, G.; Bick, T.; Lewinson, D.; Machtei, E.E. Human blood-derived endothelial progenitor cells augment vasculogenesis and osteogenesis. J. Clin. Periodontol. 2015, 42, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Zigdon-Giladi, H.; Elimelech, R.; Michaeli-Geller, G.; Rudich, U.; Machtei, E.E. Safety profile and long-term engraftment of human CD31 + blood progenitors in bone tissue engineering. Cytotherapy 2017, 19, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Sabbah, N.; Tamari, T.; Elimelech, R.; Doppelt, O.; Rudich, U.; Zigdon-Giladi, H. Predicting Angiogenesis by Endothelial Progenitor Cells Relying on In-Vitro Function Assays and VEGFR-2 Expression Levels. Biomolecules 2019, 9, 717. [Google Scholar] [CrossRef] [PubMed]
- Costa-Pinto, A.R.; Correlo, V.M.; Sol, P.; Bhattacharya, M.; Srouji, S.; Livne, E.; Reis, R.L.; Neves, N.M. Chitosan-poly (butylene succinate) scaffolds and human bone marrow stromal cells induce bone repair in a mouse calvaria model. J. Tissue Eng. Regen Med. 2012, 6, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zigdon-Giladi, H.; Lewinson, D.; Bick, T.; Machtei, E.E. Vertical bone augmentation using different osteoconductive scaffolds combined with barrier domes in the rat calvarium. Clin. Implant. Dent. Relat. Res. 2014, 16, 138–144. [Google Scholar] [CrossRef]
- Miller, G.E.; Chen, E. Life stress and diminished expression of genes encoding glucocorticoid receptor and β2-adrenergic receptor in children with asthma. Proc. Natl. Acad. Sci. USA 2006, 103, 5496–5501. [Google Scholar] [CrossRef] [Green Version]
- Kawamoto, A.; Asahara, T.; Losordo, D.W. Transplantation of endothelial progenitor cells for therapeutic neovascularization. Cardiovasc. Radiat. Med. 2002, 3, 221–225. [Google Scholar] [CrossRef]
- Asahara, T.; Kawamoto, A. Endothelial progenitor cells for postnatal vasculogenesis. Am. J. Physiol. Cell Physiol. 2004, 287, C572–C579. [Google Scholar] [CrossRef] [Green Version]
- Zigdon-Giladi, H.; Bick, T.; Morgan, E.F.; Lewinson, D.; Machtei, E.E. Peripheral Blood-Derived Endothelial Progenitor Cells Enhance Vertical Bone Formation. Clin. Implant. Dent. Relat. Res. 2015, 17, 83–92. [Google Scholar] [CrossRef]
- Machtei, E.E.; Kim, D.M.; Karimbux, N.; Giladi, H.Z. The use of endothelial progenitor cells combined with barrier membrane for the reconstruction of peri-implant osseous defects: An animal experimental study. J. Clin. Periodontol. 2016, 43, 289–297. [Google Scholar] [CrossRef]
- Woodrow, D. Endothelial Cells 3 Volumes; Ryan Una, S., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1988; Volume 1, p. 191; Volume 2, p. 282; Volume 3, p. 251. [Google Scholar]
- Mantovani, A.; Dejana, E. Cytokines as communication signals between leukocytes and endothelial cells. Immunol. Today 1989, 10, 370–375. [Google Scholar] [CrossRef]
- Seelentag, W.K.; Mermod, J.J.; Montesano, R.; Vassalli, P. Additive effects of interleukin 1 and tumour necrosis factor-alpha on the accumulation of the three granulocyte and macrophage colony-stimulating factor mRNAs in human endothelial cells. EMBO J. 1987, 6, 2261–2265. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.M.J.; Ferrige, A.G.; Moncada, S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 1987, 327, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, M.; Alam, A.; Bax, B.E.; Shankar, V.; Bax, C.; Gill, J.; Pazianas, M.; Huang, C.-H.; Sahinoglu, T.; Moonga, B.; et al. Role of the endothelial cell in osteoclast control: New perspectives. Bone 1993, 14, 97–102. [Google Scholar] [CrossRef]
- Falasca, G.F.; Ramachandrula, A.; Kelley, K.A.; O’Connor, C.R.; Reginato, A.J. Superoxide anion production and phagocytosis of crystals by cultured endothelial cells. Arthritis Rheum. 1993, 36, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Dauwe, D.; Pelacho, B.; Wibowo, A.; Walravens, A.; Verdonck, K.; Gillijns, H.; Caluwe, E.; Pokreisz, P.; Van Gastel, N.; Carmeliet, G.; et al. Neovascularization Potential of Blood Outgrowth Endothelial Cells from Patients with Stable Ischemic Heart Failure Is Preserved. J. Am. Heart Assoc. 2016, 5, 002288. [Google Scholar] [CrossRef] [Green Version]
- Laing, A.J.; Dillon, J.P.; Condon, E.T.; Street, J.; Wang, J.; McGuinness, A.; Redmond, H. Mobilization of endothelial precursor cells: Systemic vascular response to musculoskeletal trauma. J. Orthop. Res. 2007, 25, 44–50. [Google Scholar] [CrossRef]
- Scott, M.A.; Levi, B.; Askarinam, A.; Nguyen, A.; Rackohn, T.; Ting, K.; Soo, C.; James, A.W. Brief review of models of ectopic bone formation. Stem Cells Dev. 2012, 21, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Schmid, G.J.; Kobayashi, C.; Sandell, L.J.; Ornitz, D.M. Fibroblast growth factor expression during skeletal fracture healing in mice. Dev. Dyn. 2009, 238, 766–774. [Google Scholar] [CrossRef] [Green Version]
- Ohgushi, H.; Okumura, M.; Tamai, S.; Shors, E.C.; Caplan, A. Marrow cell induced osteogenesis in porous hydroxyapatite and tricalcium phosphate: A comparative histomorphometric study of ectopic bone formation. J. Biomed. Mater. Res. 1990, 24, 1563–1570. [Google Scholar] [CrossRef]
- Sheikh, Z.; Sima, C.; Glogauer, M. Bone Replacement Materials and Techniques Used for Achieving Vertical Alveolar Bone Augmentation. Materials 2015, 8, 2953–2993. [Google Scholar] [CrossRef]
- Erbe, E.; Marx, J.; Clineff, T.; Bellincampi, L. Potential of an ultraporous β-tricalcium phosphate synthetic cancellous bone void filler and bone marrow aspirate composite graft. Eur. Spine J. 2001, 10, S141–S146. [Google Scholar] [PubMed] [Green Version]
- Seebach, C.; Henrich, D.; Kähling, C.; Wilhelm, K.; Tami, A.E.; Alini, M.; Marzi, I. Endothelial progenitor cells and mesenchymal stem cells seeded onto beta-TCP granules enhance early vascularization and bone healing in a critical-sized bone defect in rats. Tissue Eng. Part A 2010, 16, 1961–1970. [Google Scholar]
- Szpalski, M.; Gunzburg, R. Applications of calcium phosphate-based cancellous bone void fillers in trauma surgery. Orthopedics 2002, 25, 601–609. [Google Scholar]
- De Groot, K. Effect of porosity and physicochemical properties on the stability, resorption, and strength of calcium phosphate ceramics. Ann. New York Acad. Sci. 1988, 523, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Zerbo, I.R.; Zijderveld, S.A.; De Boer, A.; Bronckers, A.L.J.J.; De Lange, G.; Bruggenkate, C.M.T.; Burger, E.H. Histomorphometry of human sinus floor augmentation using a porous beta-tricalcium phosphate: A prospective study. Clin. Oral Implant. Res. 2004, 15, 724–732. [Google Scholar] [CrossRef]
- Horowitz, R.A.; Mazor, Z.; Foitzik, C.; Prasad, H.; Rohrer, M.; Palti, A. β-tricalcium phosphate as bone substitute material: Properties and clinical applications. J. Osseointegration 2010, 2, 61–68. [Google Scholar]
- Wang, J.; Chen, W.; Li, Y.; Fan, S.; Weng, J.; Zhang, X. Biological evaluation of biphasic calcium phosphate ceramic vertebral laminae. Biomaterials 1998, 19, 1387–1392. [Google Scholar] [CrossRef]
- Henrich, D.; Seebach, C.; Kaehling, C.; Scherzed, A.; Wilhelm, K.; Tewksbury, R.; Powerski, M.; Marzi, I. Simultaneous cultivation of human endothelial-like differentiated precursor cells and human marrow stromal cells on beta-tricalcium phosphate. Tissue Eng. Part C Methods 2009, 15, 551–560. [Google Scholar] [CrossRef]
- Jensen, E.C. Use of fluorescent probes: Their effect on cell biology and limitations. Anat. Rec. Adv. Integr. Anat. Evol. Boil. 2012, 295, 2031–2036. [Google Scholar] [CrossRef]
- Camp, D.M.; Loeffler, D.A.; Farrah, D.M.; Borneman, J.N.; LeWitt, P.A. Cellular immune response to intrastriatally implanted allogeneic bone marrow stromal cells in a rat model of Parkinson’s disease. J. Neuroinflamm. 2009, 6, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galleu, A.; Riffo-Vasquez, Y.; Trento, C.; Lomas, C.; Dolcetti, L.; Cheung, T.S.; Von Bonin, M.; Barbieri, L.; Halai, K.; Ward, S.; et al. Apoptosis in mesenchymal stromal cells induces in vivo recipient-mediated immunomodulation. Sci. Transl. Med. 2017, 9, eaam7828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, W.; Kim, K.L.; Kim, J.-M.; Shin, I.-S.; Lee, Y.-S.; Lee, J.-Y.; Jang, H.-S.; Lee, J.-S.; Byun, J.; Choi, J.-H.; et al. Transplantation of endothelial progenitor cells accelerates dermal wound healing with increased recruitment of monocytes/macrophages and neovascularization. Stem Cells 2005, 23, 1571–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamoto, A.; Gwon, H.-C.; Iwaguro, H.; Yamaguchi, J.-I.; Uchida, S.; Masuda, H.; Silver, M.; Ma, H.; Kearney, M.; Isner, J.M.; et al. Therapeutic Potential of Ex Vivo Expanded Endothelial Progenitor Cells for Myocardial Ischemia. Circulation 2001, 103, 634–637. [Google Scholar] [CrossRef]
- Schuster, M.D.; Kocher, A.A.; Seki, T.; Martens, T.P.; Xiang, G.; Homma, S.; Itescu, S. Myocardial neovascularization by bone marrow angioblasts results in cardiomyocyte regeneration. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H525–H532. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-J.; Lee, N.; Lee, J.Y.; Choi, Y.J.; Ii, M.; Wecker, A.; Jeong, J.-O.; Curry, C.; Qin, G.; Yoon, Y. Role of host tissues for sustained humoral effects after endothelial progenitor cell transplantation into the ischemic heart. J. Exp. Med. 2007, 204, 3257–3269. [Google Scholar] [CrossRef]
- He, T.; Peterson, T.E.; Katusic, Z.S. Paracrine mitogenic effect of human endothelial progenitor cells: Role of interleukin-8. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H968–H972. [Google Scholar] [CrossRef]
- Urbich, C.; Aicher, A.; Heeschen, C.; Dernbach, E.; Hofmann, W.K.; Zeiher, A.M.; Dimmeler, S. Soluble factors released by endothelial progenitor cells promote migration of endothelial cells and cardiac resident progenitor cells. J. Mol. Cell. Cardiol. 2005, 39, 733–742. [Google Scholar] [CrossRef]
- Henrich, D.; Wilhelm, K.; Warzecha, J.; Frank, J.; Barker, J.; Marzi, I.; Seebach, C. Human Endothelial-Like Differentiated Precursor Cells Maintain Their Endothelial Characteristics When Cocultured with Mesenchymal Stem Cell and Seeded onto Human Cancellous Bone. Mediat. Inflamm. 2013, 2013, e364591. Available online: https://www.hindawi.com/journals/mi/2013/364591/ (accessed on 3 January 2013). [CrossRef]
- Han, Y.; Hsieh, F.H. Osteogenic Differentiation of Late-Outgrowth CD45-Negative Endothelial Progenitor Cells. J. Vasc. Res. 2014, 51, 369–375. [Google Scholar] [CrossRef]
- Pittenger, M.F. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakitani, S.; Saito, T.; Caplan, A. Myogenic cells derived from rat bone marrow mesenchymal stem cells exposed to 5-azacytidine. Muscle Nerve 1995, 18, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Fedorovich, N.E.; Haverslag, R.T.; Dhert, W.; Alblas, J. The role of endothelial progenitor cells in prevascularized bone tissue engineering: Development of heterogeneous constructs. Tissue Eng. Part A 2010, 16, 2355–2367. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.; Planell, J.A.; Engel, E. Dynamics of bone marrow-derived endothelial progenitor cell/mesenchymal stem cell interaction in co-culture and its implications in angiogenesis. Biochem. Biophys. Res. Commun. 2010, 400, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Möhle, R.; Bautz, F.; Rafii, S.; Moore, M.A.; Brugger, W.; Kanz, L. The chemokine receptor CXCR-4 is expressed on CD34+ hematopoietic progenitors and leukemic cells and mediates transendothelial migration induced by stromal cell-derived factor-1. Blood 1998, 91, 4523–4530. [Google Scholar] [CrossRef]
- Lataillade, J.-J.; Clay, D.; Dupuy, C.; Rigal, S.; Jasmin, C.; Bourin, P.; Le Bousse-Kerdilès, M.-C. Chemokine SDF-1 enhances circulating CD34 (+) cell proliferation in synergy with cytokines: Possible role in progenitor survival. Blood 2000, 95, 756–768. [Google Scholar] [CrossRef]
- Yamaguchi, J.-I.; Kusano, K.F.; Masuo, O.; Kawamoto, A.; Silver, M.; Murasawa, S.; Bosch-Marce, M.; Masuda, H.; Losordo, D.W.; Isner, J.M.; et al. Stromal cell-derived factor-1 effects on ex vivo expanded endothelial progenitor cell recruitment for ischemic neovascularization. Circulation 2003, 107, 1322–1328. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Liu, X.; Ou, L.; Zhou, X.; Liu, Y.; Jia, X.; Zhang, J.; Li, Y.; Kong, D. Mobilization of mesenchymal stem cells by granulocyte colony-stimulating factor in rats with acute myocardial infarction. Cardiovasc. Drugs Ther. 2008, 22, 363–371. [Google Scholar] [CrossRef]
- Cheng, Z.; Ou, L.; Zhou, X.; Li, F.; Jia, X.; Zhang, Y.; Liu, X.; Li, Y.; Ward, C.A.; Melo, L.G.; et al. Targeted migration of mesenchymal stem cells modified with CXCR4 gene to infarcted myocardium improves cardiac performance. Mol. Ther. 2008, 16, 571–579. [Google Scholar] [CrossRef]
- Liu, H.; Xue, W.; Ge, G.; Luo, X.; Li, Y.; Xiang, H.; Ding, X.; Tian, P.-X.; Tian, X. Hypoxic preconditioning advances CXCR4 and CXCR7 expression by activating HIF-1α in MSCs. Biochem. Biophys. Res. Commun. 2010, 401, 509–515. [Google Scholar] [CrossRef]
- Otsuru, S.; Tamai, K.; Yamazaki, T.; Yoshikawa, H.; Kaneda, Y. Circulating bone marrow-derived osteoblast progenitor cells are recruited to the bone-forming site by the CXCR4/stromal cell-derived factor-1 pathway. Stem Cells 2008, 26, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, R.; Wasserman, K.; Young, H.A.; Grimm, M.; Howard, O.; Anver, M.R.; Kleinman, H.K.; Murphy, W.J.; Oppenheim, J.J. Vascular endothelial growth factor and basic fibroblast growth factor induce expression of CXCR4 on human endothelial cells: In vivo neovascularization induced by stromal-derived factor-1alpha. Am. J. Pathol. 1999, 154, 1125–1135. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, C.; Li, Y.; Ji, Y.; Xu, G.; Wang, X.; Yan, J. SDF-1 promotes endochondral bone repair during fracture healing at the traumatic brain injury condition. PLoS ONE 2013, 8, e54077. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamari, T.; Kawar-Jaraisy, R.; Doppelt, O.; Giladi, B.; Sabbah, N.; Zigdon-Giladi, H. The Paracrine Role of Endothelial Cells in Bone Formation via CXCR4/SDF-1 Pathway. Cells 2020, 9, 1325. https://doi.org/10.3390/cells9061325
Tamari T, Kawar-Jaraisy R, Doppelt O, Giladi B, Sabbah N, Zigdon-Giladi H. The Paracrine Role of Endothelial Cells in Bone Formation via CXCR4/SDF-1 Pathway. Cells. 2020; 9(6):1325. https://doi.org/10.3390/cells9061325
Chicago/Turabian StyleTamari, Tal, Rawan Kawar-Jaraisy, Ofri Doppelt, Ben Giladi, Nadin Sabbah, and Hadar Zigdon-Giladi. 2020. "The Paracrine Role of Endothelial Cells in Bone Formation via CXCR4/SDF-1 Pathway" Cells 9, no. 6: 1325. https://doi.org/10.3390/cells9061325
APA StyleTamari, T., Kawar-Jaraisy, R., Doppelt, O., Giladi, B., Sabbah, N., & Zigdon-Giladi, H. (2020). The Paracrine Role of Endothelial Cells in Bone Formation via CXCR4/SDF-1 Pathway. Cells, 9(6), 1325. https://doi.org/10.3390/cells9061325