Potential Prognostic Role of SPARC Methylation in Non-Small-Cell Lung Cancer

 , ,
, ,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Patients and Tumor Tissue Samples
2.3. DNA and RNA Extraction
2.4. Cell Culture and 5-Aza-2′-Deoxycytidine (5-Aza-CdR) Treatment
2.5. Proliferation, Viability, Migration and Invasion Assays
2.6. Immunoistochemistry (IHC)
2.7. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.8. Sodium Bisulfite Conversion and Quantitative Methylation Specific PCR Analysis (QMSP)
2.9. Mutation Screening of Epidermal Growth Factor Receptor Tyrosine Kinase (EGFR) and Kirsten Rat Sarcoma Viral Oncogene Homolog (KRAS) Genes by Sanger Sequencing
2.10. TCGA Data Analysis
2.11. Statistical Analysis
3. Results
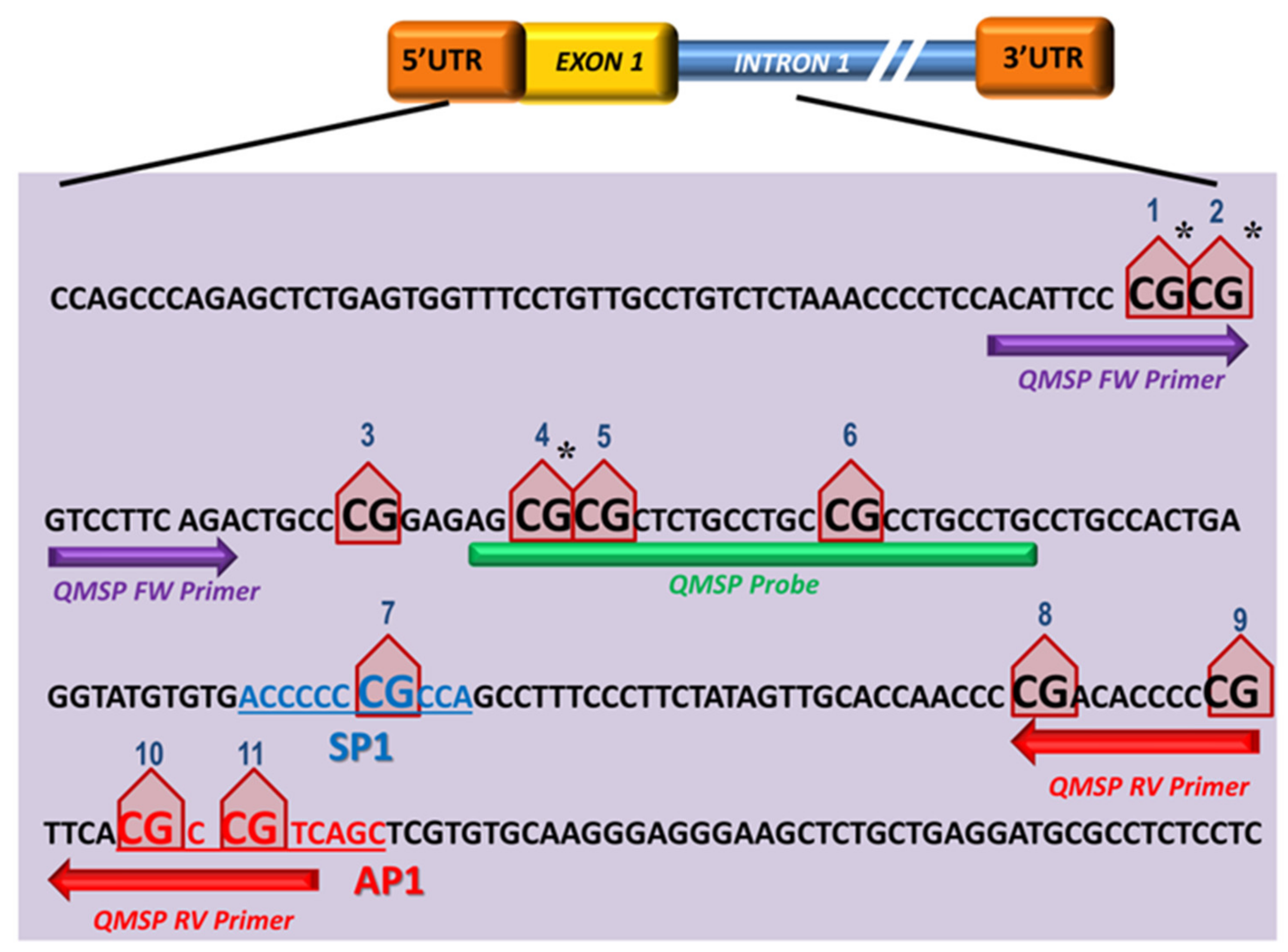
3.1. SPARC CpG Island Prediction and QMSP Assay Optimization
3.2. Aberrant SPARC Methylation Is a Frequent Event in Primary NSCLCs
3.3. Hypermethylation of SPARC Gene in NSCLC Cell Lines and Association with Reduced SPARC mRNA Level
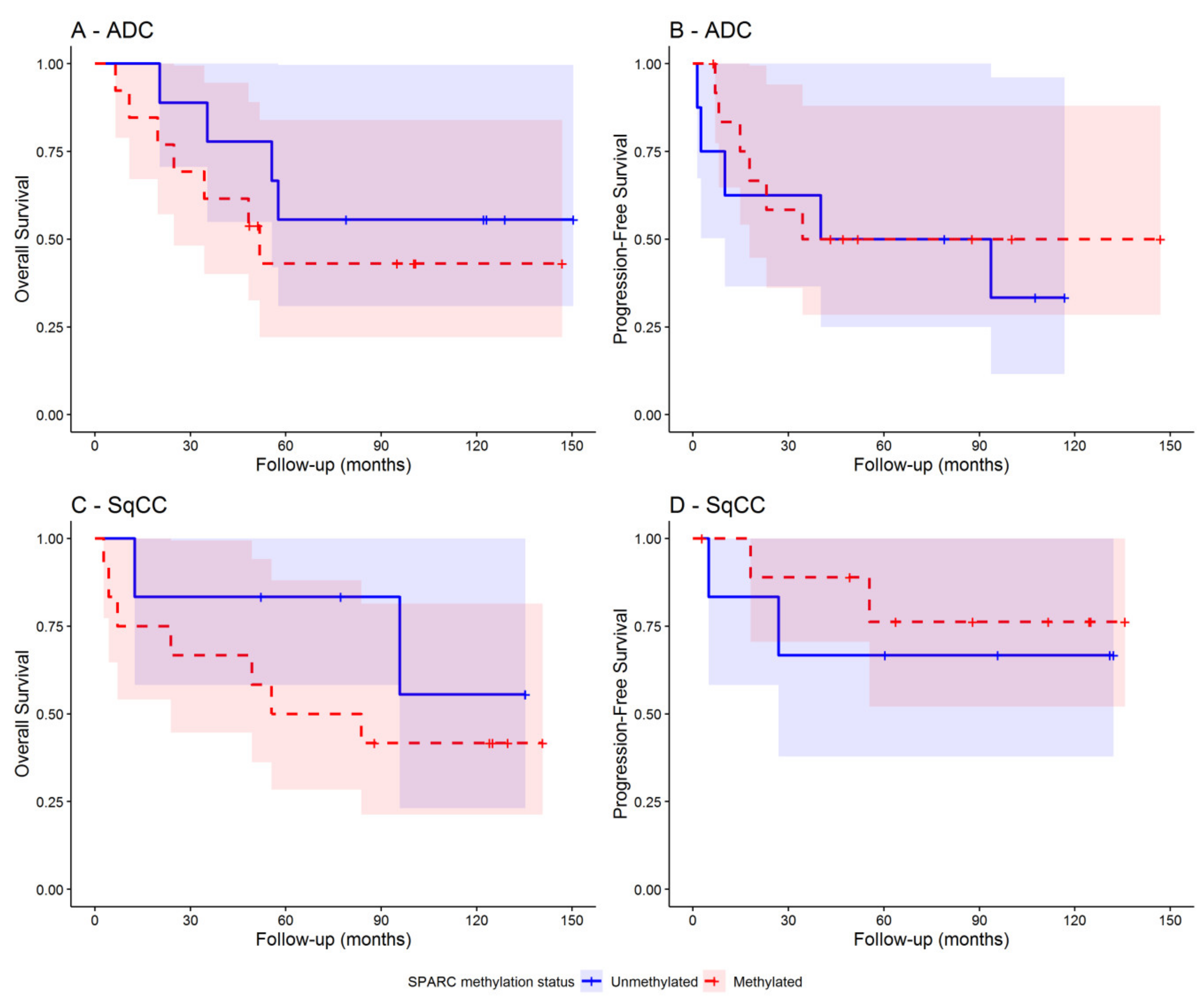
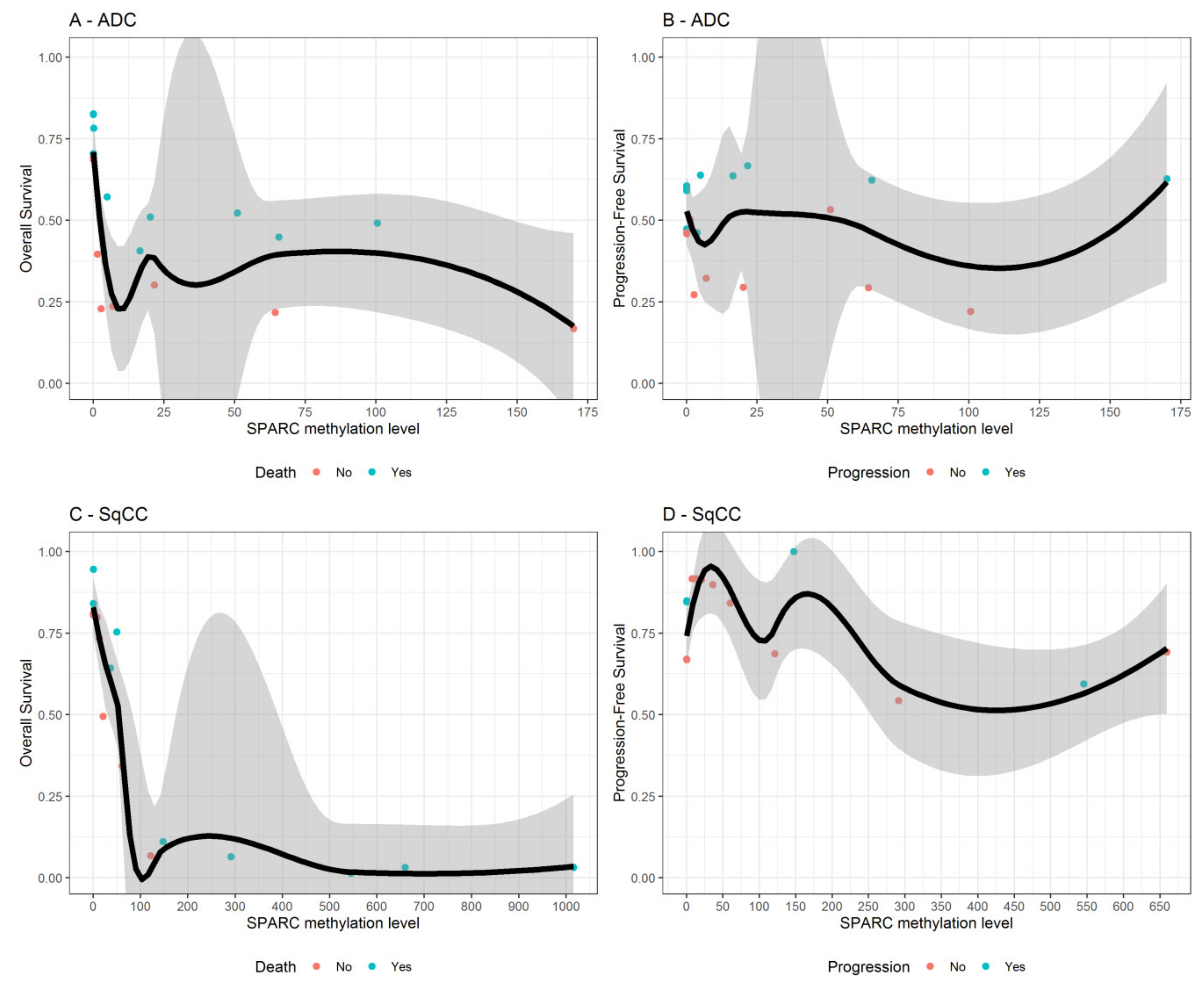
3.4. SPARC Hypermethylation Is Associated with Higher Mortality Risk in SqCC Ratients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dela Cruz, C.S.; Tanoue, L.T.; Matthay, R.A. Lung cancer: Epidemiology, etiology, and prevention. Clin. Chest. Med. 2011, 32, 605–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thun, M.; Linet, M.S.; Cerhan, J.R.; Haiman, C.A.; Schottenfeld, D. Cancer Epidemiology and Prevention, 4th ed.; Thun, M., Linet, M.S., Cerhan, J.R., Haiman, C.A., Schottenfeld, D., Eds.; O.U. Press: New York, NY, USA, 2018; pp. 519–542. [Google Scholar]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.A.; Brekken, R.A. SPARC: A matricellular regulator of tumorigenesis. J. Cell Commun. Signal 2009, 3, 255–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.L.; Sukkar, M.B. The SPARC protein: An overview of its role in lung cancer and pulmonary fibrosis and its potential role in chronic airways disease. Br. J. Pharmacol. 2017, 174, 3–14. [Google Scholar] [CrossRef]
- Brekken, R.A.; Sage, E.H. SPARC, a matricellular protein: At the crossroads of cell-matrix communication. Matrix Biol. 2001, 19, 816–827. [Google Scholar] [CrossRef]
- Bradshaw, A.D. Diverse biological functions of the SPARC family of proteins. Int. J. Biochem. Cell Biol. 2012, 44, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Gagliardi, F.; Narayanan, A.; Mortini, P. SPARCL1 a novel player in cancer biology. Crit. Rev. Oncol. Hematol. 2016, 109, 63–68. [Google Scholar] [CrossRef]
- Xu, J.; Yang, S.; Gu, X.; Shen, H.; Wang, L.; Xu, X.; Fang, L.; Mao, Y.; Xu, L.; Chen, Y. SPARC correlates with unfavorable outcome and promotes tumor growth in lung squamous cell carcinoma. Exp. Mol. Pathol. 2019, 110, 104276. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, J.; Zhao, Y.Y.; Jiang, W.; Xue, C.; Xu, F.; Zhao, H.Y.; Zhang, Y.; Zhao, L.P.; Hu, Z.H.; et al. SPARC expression and prognostic value in non-small cell lung cancer. Chin. J. Cancer 2012, 31, 541–548. [Google Scholar] [CrossRef]
- Bertino, E.M.; Williams, T.M.; Nana-Sinkam, S.P.; Shilo, K.; Chatterjee, M.; Mo, X.; Rahmani, M.; Phillips, G.S.; Villalona-Calero, M.A.; Otterson, G.A. Stromal Caveolin-1 Is Associated With Response and Survival in a phase II trial of nab-paclitaxel with carboplatin for advanced NSCLC Patients. Clin. Lung Cancer 2015, 16, 466–474. [Google Scholar] [CrossRef] [Green Version]
- Komiya, K.; Nakamura, T.; Nakashima, C.; Takahashi, K.; Umeguchi, H.; Watanabe, N.; Sato, A.; Takeda, Y.; Kimura, S.; Sueoka-Aragane, N. SPARC is a possible predictive marker for albumin-bound paclitaxel in non-small-cell lung cancer. Onco. Targets Ther. 2016, 9, 6663–6668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isler, S.G.; Schenk, S.; Bendik, I.; Schraml, P.; Novotna, H.; Moch, H.; Sauter, G.; Ludwig, C.U. Genomic organization and chromosomal mapping of SPARC-like 1, a gene down regulated in cancers. Int. J. Oncol. 2001, 18, 521–526. [Google Scholar] [CrossRef]
- Sato, N.; Fukushima, N.; Maehara, N.; Matsubayashi, H.; Koopmann, J.; Su, G.H.; Hruban, R.H.; Goggins, M. SPARC/osteonectin is a frequent target for aberrant methylation in pancreatic adenocarcinoma and a mediator of tumor-stromal interactions. Oncogene 2003, 22, 5021–5030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, E.; Kang, H.J.; Koh, K.H.; Rhee, H.; Kim, N.K.; Kim, H. Frequent inactivation of SPARC by promoter hypermethylation in colon cancers. Int. J. Cancer 2007, 121, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Cheetham, S.; Tang, M.J.; Mesak, F.; Kennecke, H.; Owen, D.; Tai, I.T. SPARC promoter hypermethylation in colorectal cancers can be reversed by 5-Aza-2’deoxycytidine to increase SPARC expression and improve therapy response. Br. J. Cancer 2008, 98, 1810–1819. [Google Scholar] [CrossRef] [PubMed]
- Socha, M.J.; Said, N.; Dai, Y.; Kwong, J.; Ramalingam, P.; Trieu, V.; Desai, N.; Mok, S.C.; Motamed, K. Aberrant promoter methylation of SPARC in ovarian cancer. Neoplasia 2009, 11, 126–135. [Google Scholar]
- Gao, J.; Song, J.; Huang, H.; Li, Z.; Du, Y.; Cao, J.; Li, M.; Lv, S.; Lin, H.; Gong, Y. Methylation of the SPARC gene promoter and its clinical implication in pancreatic cancer. J. Exp. Clin. Cancer Res. 2010, 29, 28. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, B.; Du, Z.; Bai, T.; Gao, Y.T.; Wang, Y.J.; Lou, C.; Wang, F.M.; Bai, Y. Aberrant methylation of SPARC in human hepatocellular carcinoma and its clinical implication. World J. Gastroenterol. 2012, 18, 2043–2052. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Wei, J.; Zhang, J.; Jiang, H.; Wang, S.; Zhou, X.; Zhang, Z.; Huang, G.; Watanabe, H.; Su, J. Aberrant methylation of secreted protein, acidic and rich in cysteine in human laryngeal and hypopharyngeal carcinoma. Oncol. Lett. 2011, 2, 725–729. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Zhang, J.L.; Yao, H.X.; Wang, P.Y.; Zhu, J.; Wang, W.; Wang, X.; Wan, Y.L.; Chen, S.W.; Chen, G.W.; et al. Aberrant methylation of the SPARC gene promoter and its clinical implication in gastric cancer. Sci. Rep. 2014, 4, 7035. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Qiu, X.; Zhao, X.; Yang, R.; Lian, H.; Qu, F.; Li, X.; Guo, H. Hypermethylation of the SPARC promoter and its prognostic value for prostate cancer. Oncol. Rep. 2018, 39, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Rigoutsos, I.; Lee, S.K.; Nam, S.Y.; Anfossi, S.; Pasculli, B.; Pichler, M.; Jing, Y.; Rodriguez-Aguayo, C.; Telonis, A.G.; Rossi, S.; et al. N-BLR, a primate-specific non-coding transcript leads to colorectal cancer invasion and migration. Genome Biol. 2017, 18, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.Y.L.; Wei, J.; Ying, Z. Checking the cox model with cumulative sums of martingale-based residuals. Biometrika. 1993, 80, 557–572. [Google Scholar] [CrossRef]
- Ishwaran, H.; Kogalur, U.B.; Blackstone, E.H.; Lauer, M.S. Random survival forests. Ann. Appl. Stat. 2008, 80, 841–860. [Google Scholar] [CrossRef]
- Breiman, L. Random Forest. Mach. Learn 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Koukourakis, M.I.; Giatromanolaki, A.; Brekken, R.A.; Sivridis, E.; Gatter, K.C.; Harris, A.L.; Sage, E.H. Enhanced expression of SPARC/osteonectin in the tumor-associated stroma of non-small cell lung cancer is correlated with markers of hypoxia/acidity and with poor prognosis of patients. Cancer Res. 2003, 63, 5376–5380. [Google Scholar] [PubMed]
- Schneider, S.; Yochim, J.; Brabender, J.; Uchida, K.; Danenberg, K.D.; Metzger, R.; Schneider, P.M.; Salonga, D.; Hölscher, A.H.; Danenberg, P.V. Osteopontin but not osteonectin messenger RNA expression is a prognostic marker in curatively resected non-small cell lung cancer. Clin. Cancer Res. 2004, 10, 1588–1596. [Google Scholar] [CrossRef] [Green Version]
- Isler, S.G.; Ludwig, C.U.; Chiquet-Ehrismann, R.; Schenk, S. Evidence for transcriptional repression of SPARC-like 1, a gene downregulated in human lung tumors. Int. J. Oncol. 2004, 25, 1073–1079. [Google Scholar]
- Sato, T.; Arai, E.; Kohno, T.; Takahashi, Y.; Miyata, S.; Tsuta, K.; Watanabe, S.; Soejima, K.; Betsuyaku, T.; Kanai, Y. Epigenetic clustering of lung adenocarcinomas based on DNA methylation profiles in adjacent lung tissue: Its correlation with smoking history and chronic obstructive pulmonary disease. Int. J. Cancer 2014, 135, 319–334. [Google Scholar] [CrossRef]
- Suddiq, F.; Sarkar, F.H.; Wali, A.; Pass, H.I.; Lonardo, F. Increased osteonectin expression is associated with malignant transformation and tumor associated fibrosis in the lung. Lung Cancer 2004, 45, 197–205. [Google Scholar] [CrossRef]
- Torres-Nunez, E.; Cal, L.; Suárez-Bregua, P.; Gómez-Marin, C.; Moran, P.; Gómez-Skarmeta, J.L.; Rotllant, J. Matricellular protein SPARC/osteonectin expression is regulated by DNA methylation in its core promoter region. Dev. Dyn. 2015, 244, 693–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.L.; Chiang, P.H.; Huang, C.C.; Su, Y.L.; Sung, M.T.; Tsai, E.M.; Lin, C.S.; Chiang, P.H. Methylation of SPARCL1 Is Associated with oncologic outcome of advanced upper urinary tract urothelial carcinoma. Int. J. Mol. Sci. 2019, 20, 1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, S.; Zhou, N.M.; Dai, D.Q. Aberrant methylation of secreted protein acidic and rich in cysteine gene and its significance in gastric cancer. World J. Gastroenterol. 2019, 25, 6713–6727. [Google Scholar] [CrossRef]
- Leal, A.; Sidransky, D.; Brait, M. Tissue and cell-Free DNA-based epigenomic approaches for cancer detection. Clin. Chem. 2019, 66, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Dobersch, S.; Romero-Olmedo, A.J.; Barreto, G. Epigenetics in lung cancer diagnosis and therapy. Cancer Metastasis Rev. 2015, 34, 229–241. [Google Scholar] [CrossRef]
- Ooki, A.; Maleki, Z.; Tsay, J.J.; Goparaju, C.; Brait, M.; Turaga, N.; Nam, H.S.; Rom, W.N.; Pass, H.I.; Sidransky, D.; et al. A panel of novel detection and prognostic methylated DNA markers in primary non-small cell lung cancer and serum DNA. Clin. Cancer Res. 2017, 23, 7141–7152. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | TOTAL (n = 59) | ADC (n = 34) | SqCC (n = 25) |
|---|---|---|---|
| Age at Diagnosis (mean yy ± SD) | 67.7 ± 8.4 | 67.5 ± 10.2 | 68.0 ± 5.2 |
| Gender (n, %) | |||
| Male | 50 (84.8) | 25 (73.5) | 25 (100.0) |
| Female | 9 (15.2) | 9 (26.5) | 0 (0.0) |
| Smoking status (n, %) | |||
| Smoker | 28 (47.5) | 13 (38.2) | 15 (60.0) |
| Never smoker | 6 (10.1%) | 6 (17.7) | 0 (0.0) |
| Former smoker | 25 (42.4%) | 15 (44.1) | 10 (40.0) |
| Disease Stage | |||
| IA/B | 28 (47.5) | 18 (52.9) | 10 (40.0) |
| IIA/B | 14 (23.7) | 6 (17.7) | 8 (32.0) |
| IIIA/B | 11 (18.6) | 4 (11.7) | 7 (28.0) |
| IV | 6 (10.2) | 6 (17.7) | 0 (0.0) |
| pT § (n, %) | |||
| T1 | 20 (33.9) | 14 (41.2) | 6 (24.0) |
| T2 | 27 (45.7) | 14 (41.2) | 13 (52.0) |
| T3 | 6 (10.2) | 4 (11.7) | 2 (8.0) |
| T4 | 6 (10.2) | 2 (5.9) | 4 (16.0) |
| pN † (n, %) | |||
| N0 | 42 (71.2) | 27 (79.4) | 15 (60.0) |
| N1 | 10 (17.0) | 3 (8.8) | 7 (28.0) |
| N2 | 7 (11.8) | 4 (11.8) | 3 (12.0) |
| pM (n, %) | |||
| M0 | 54 (91.5) | 29 (85.3) | 25 (100.0) |
| M1 | 5 (8.5) | 5 (14.7) | 0 (0.0) |
| Characteristics | SPARC Unmethylated | SPARC Methylated | p-Value * |
|---|---|---|---|
| n | 23 | 36 | |
| Age at Diagnosis (mean yy ± SD) | 67.5 ± 8.7 | 67.8 ± 8.3 | 0.704 |
| Gender (n, %) | 0.726 | ||
| Male | 19 (82.6) | 31 (86.1) | |
| Female | 4 (17.4) | 5 (13.9) | |
| Smoking Status (n, %) | 0.741 | ||
| Smoker | 10 (43.5) | 18 (50.0) | |
| Never smoker | 3 (13.0) | 3 (8.3) | |
| Former smoker | 10 (43.5) | 15 (41.7) | |
| Disease Stage | 0.936 | ||
| IA/B | 11 (47.8) | 17 (47.2) | |
| IIA/B | 5 (21.7) | 9 (25.0) | |
| IIIA/B | 4 (17.4) | 7 (19.5) | |
| IV | 3 (13.1) | 3 (8.3) | |
| pT § (n, %) | 0.604 | ||
| T1 | 7 (30.4) | 13 (36.1) | |
| T2 | 12 (52.2) | 15 (41.7) | |
| T3 | 1 (4.4) | 5 (13.9) | |
| T4 | 3 (13.0) | 3 (8.3) | |
| pN (n, %) | 0.701 | ||
| N0 | 16 (69.6) | 26 (72.2) | |
| N1 | 5 (21.7) | 5 (13.9) | |
| N2 | 2 (8.7) | 5 (13.9) | |
| pM (n, %) | 0.369 | ||
| M0 | 20 (87.0) | 34 (94.4) | |
| M1 | 3 (13.0) | 2 (5.6) | |
| Histology | 0.423 | ||
| ADC | 15 (65.2) | 19 (52.8) | |
| SqCC | 8 (34.8) | 17 (47.2) | |
| KRAS Status § | 1.000 | ||
| Wild-type | 18 (90.0) | 29 (90.6) | |
| Mutated | 2 (10.0) | 3 (9.4) | |
| Missing data | 3 | 4 |
| Characteristics | n | SPARC Methylation Level ^ | p-Value # |
|---|---|---|---|
| Age at Diagnosis | 59 | r = 0.10 | 0.454 |
| Gender | 0.270 | ||
| Male | 50 | 7.25 (0.00–65.60) | |
| Female | 9 | 1.40 (0.00–4.30) | |
| Smoking Status | 0.658 | ||
| Smoker | 28 | 4.64 (0.00–62.89) | |
| Former smoker | 25 | 6.90 (0.00–64.40) | |
| Never smoked | 6 | 0.70 (0.00–26.02) | |
| pT § | 0.279 | ||
| T1 | 20 | 5.64 (0.00–35.75) | |
| T2 | 27 | 3.70 (0.00–65.60) | |
| T3 | 6 | 73.95 (21.60–173.20) | |
| T4 | 6 | 1.28 (0.00–7.60) | |
| pN † | 0.701 | ||
| N0 | 42 | 4.04 (0.00–50.90) | |
| N1 | 10 | 6.23 (0.00–545.20) | |
| N2 | 7 | 7.60 (0.00–173.20) | |
| pM | 0.110 | ||
| M0 | 54 | 7.25 (0.00–65.60) | |
| M1 | 5 | 0.00 (0.00–3.53) | |
| Histology | 0.053 | ||
| ADC | 34 | 3.12 (0.00–21.60) | |
| SqCC | 25 | 20.60 (0.00–173.20) | |
| Disease Stage | 0.282 | ||
| IA/B | 28 | 4.30 (0.00–42.68) | |
| IIA/B | 14 | 47.03 (0.00–291.25) | |
| IIIA/B | 11 | 4.30 (0.00–173.20) | |
| IV | 6 | 1.77 (0.00–4.38) | |
| KRAS Status § | 0.634 | ||
| Wild-type | 47 | 4.38 (0.00–64.40) | |
| Mutated | 5 | 6.90 (0.00–20.10) |
| Sample | Histology | Outcome | SPARC Methylation | SD | N. Events | N. Total | Group | HR (95%CI) | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| All NSCLC | All | OS | Levels | 194.59 | 31 | 54 | - | 1.46 (1.07–2.00) * | 0.018 |
| Status | - | 31 | 54 | Met vs. Unmet | 1.39 (0.66–2.90) | 0.385 | |||
| PFS | Levels | 194.59 | 28 | 51 | - | 0.98 (0.61–1.57) * | 0.931 | ||
| Status | - | 28 | 51 | Met vs. Unmet | 0.73 (0.35–1.53) | 0.400 | |||
| ADC | OS | Levels | 40.84 | 17 | 30 | - | 0.80 (0.47–1.37) * | 0.420 | |
| Status | - | 17 | 30 | Met vs. Unmet | 1.38 (0.52–3.64) | 0.518 | |||
| PFS | Levels | 40.84 | 19 | 29 | - | 0.99 (0.64–1.54) * | 0.968 | ||
| Status | - | 19 | 29 | Met vs. Unmet | 0.84 (0.34–2.08) | 0.704 | |||
| SqCC | OS | Levels | 277.08 | 14 | 24 | - | 2.04 (1.21–3.45) * | 0.008 | |
| Status | - | 14 | 24 | Met vs. Unmet | 1.41 (0.44–4.49) | 0.566 | |||
| PFS | Levels | 277.08 | 9 | 22 | - | 1.30 (0.61–2.75) * | 0.498 | ||
| Status | - | 9 | 22 | Met vs. Unmet | 0.64 (0.17–2.38) | 0.503 | |||
| Early tumor stage (I-II) NSCLC | All | OS | Levels | 200.40 | 40 | 20 | - | 1.97 (1.32–2.93) * | 0.001 |
| Status | - | 40 | 20 | Met vs. Unmet | 1.83 (0.70–4.78) | 0.217 | |||
| PFS | Levels | 200.40 | 37 | 15 | - | 1.12 (0.51–2.44) * | 0.781 | ||
| Status | - | 37 | 15 | Met vs. Unmet | 0.70 (0.25–1.95) | 0.499 | |||
| ADC | OS | Levels | 42.45 | 22 | 11 | --- | 1.00 (0.58–1.72) * | 0.994 | |
| Status | - | 22 | 11 | Met vs. Unmet | 1.75 (0.50–6.04) | 0.379 | |||
| PFS | Levels | 42.45 | 21 | 11 | - | 0.99 (0.56–1.75) * | 0.971 | ||
| Status | - | 21 | 11 | Met vs. Unmet | 0.78 (0.23–2.58) | 0.681 | |||
| SqCC | OS | Levels | 287.42 | 18 | 9 | - | 2.96 (1.43–6.12) * | 0.003 | |
| Status | - | 18 | 9 | Met vs. Unmet | 2.06 (0.43–9.93) | 0.368 | |||
| PFS | Levels | 287.42 | 16 | 4 | - | 2.03 (0.55–7.58) * | 0.290 | ||
| Status | - | 16 | 4 | Met vs. Unmet | 0.60 (0.08–4.29) | 0.613 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabrizio, F.P.; Sparaneo, A.; Fontana, A.; Mazza, T.; Graziano, P.; Pantalone, A.; Parente, P.; Centra, F.; Orlando, N.; Trombetta, D.; et al. Potential Prognostic Role of SPARC Methylation in Non-Small-Cell Lung Cancer. Cells 2020, 9, 1523. https://doi.org/10.3390/cells9061523
Fabrizio FP, Sparaneo A, Fontana A, Mazza T, Graziano P, Pantalone A, Parente P, Centra F, Orlando N, Trombetta D, et al. Potential Prognostic Role of SPARC Methylation in Non-Small-Cell Lung Cancer. Cells. 2020; 9(6):1523. https://doi.org/10.3390/cells9061523
Chicago/Turabian StyleFabrizio, Federico Pio, Angelo Sparaneo, Andrea Fontana, Tommaso Mazza, Paolo Graziano, Angela Pantalone, Paola Parente, Flavia Centra, Natalizia Orlando, Domenico Trombetta, and et al. 2020. "Potential Prognostic Role of SPARC Methylation in Non-Small-Cell Lung Cancer" Cells 9, no. 6: 1523. https://doi.org/10.3390/cells9061523
APA StyleFabrizio, F. P., Sparaneo, A., Fontana, A., Mazza, T., Graziano, P., Pantalone, A., Parente, P., Centra, F., Orlando, N., Trombetta, D., la Torre, A., Ferretti, G. M., Taurchini, M., Di Micco, C. M., Maiello, E., Fazio, V. M., Rossi, A., & Muscarella, L. A. (2020). Potential Prognostic Role of SPARC Methylation in Non-Small-Cell Lung Cancer. Cells, 9(6), 1523. https://doi.org/10.3390/cells9061523