Mechanistic Models of Signaling Pathways Reveal the Drug Action Mechanisms behind Gender-Specific Gene Expression for Cancer Treatments


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Source, Selection Criteria, and Data Preprocessing
2.2. Differential Gene Expression
2.3. Rationale of the Signaling Circuit Activity Mechanistic Model
2.4. Cell Functional Output Triggered by the Signaling Circuit
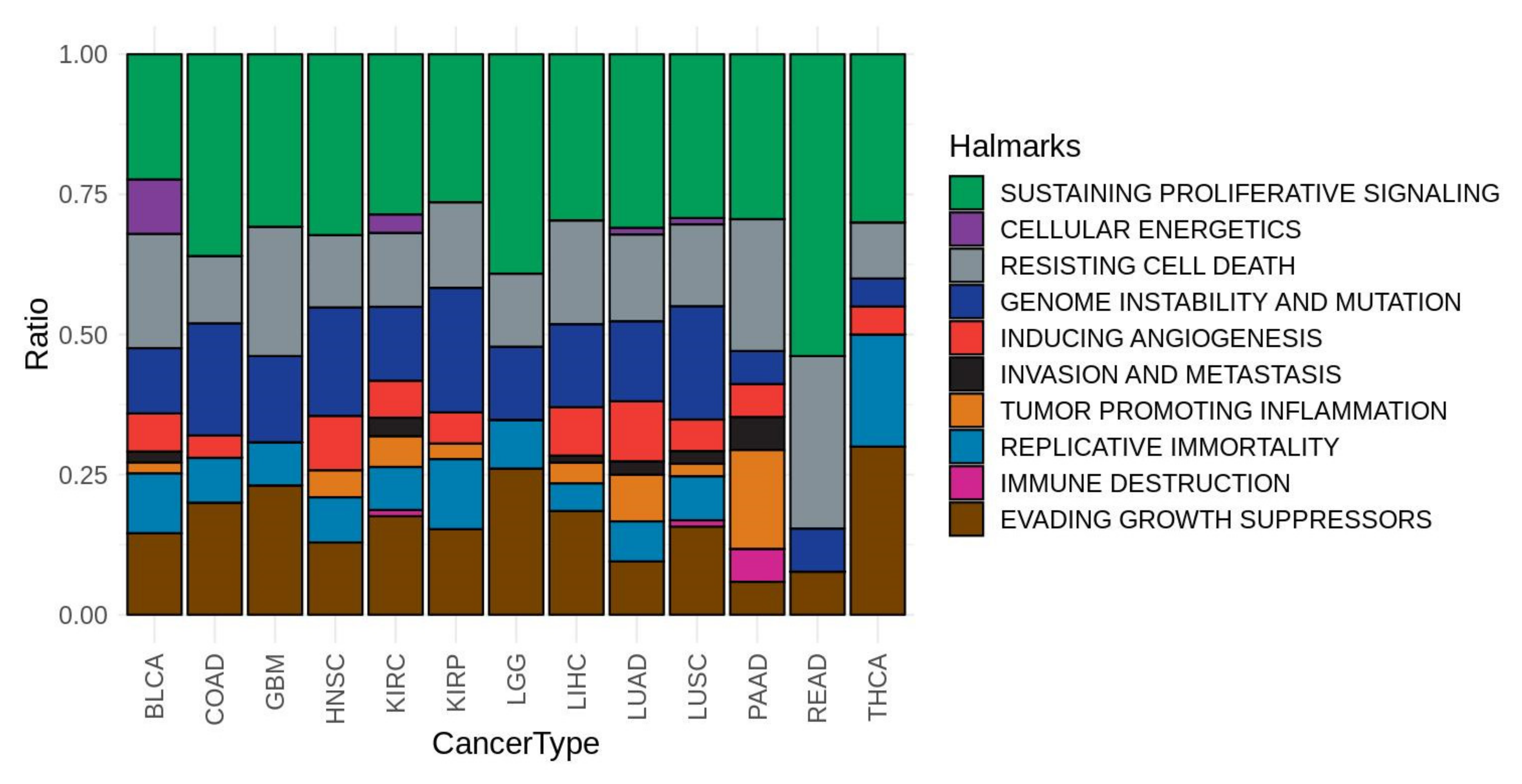
2.5. Association of Signaling Circuits Activities to Cancer Hallmarks
2.6. Estimation of the Differential Signaling Activity
2.7. Drug Effect Simulation
2.8. Differential Drug Effect between Male and Female Patients
3. Results
3.1. Data Processing
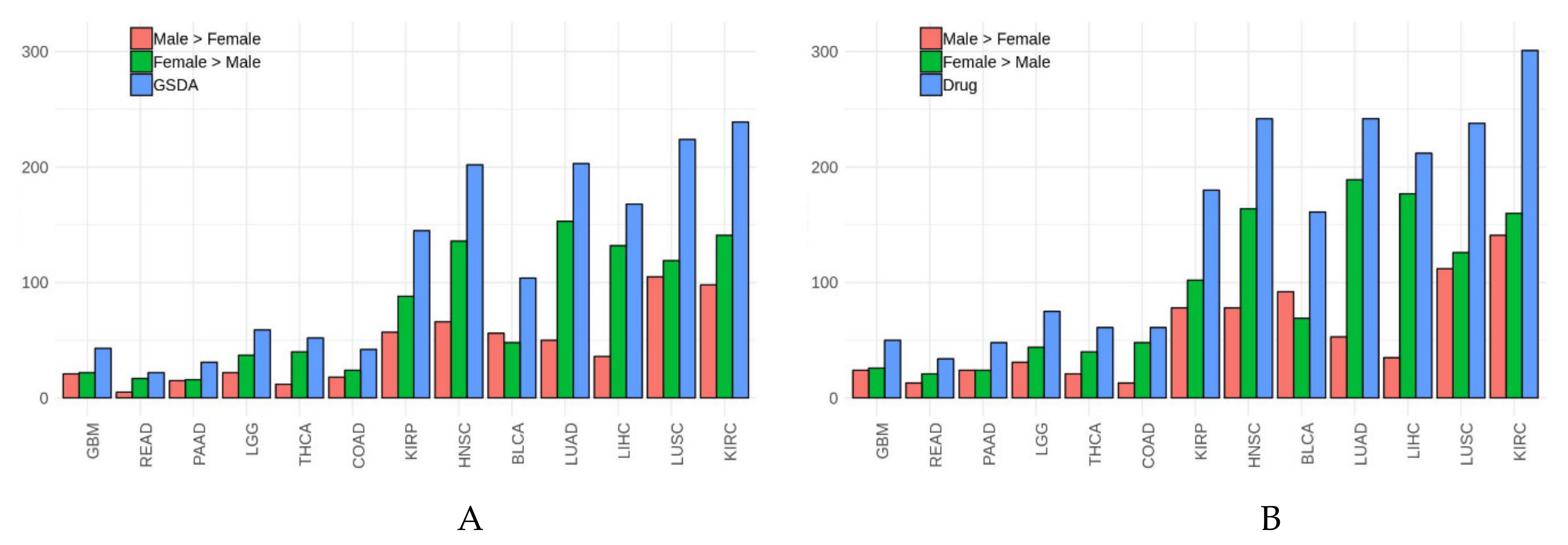
3.2. Gender-Specific Functional Differences in Cancer
3.3. Potential Differences in Drug Effects Due to Gender-Specific Functional Differences
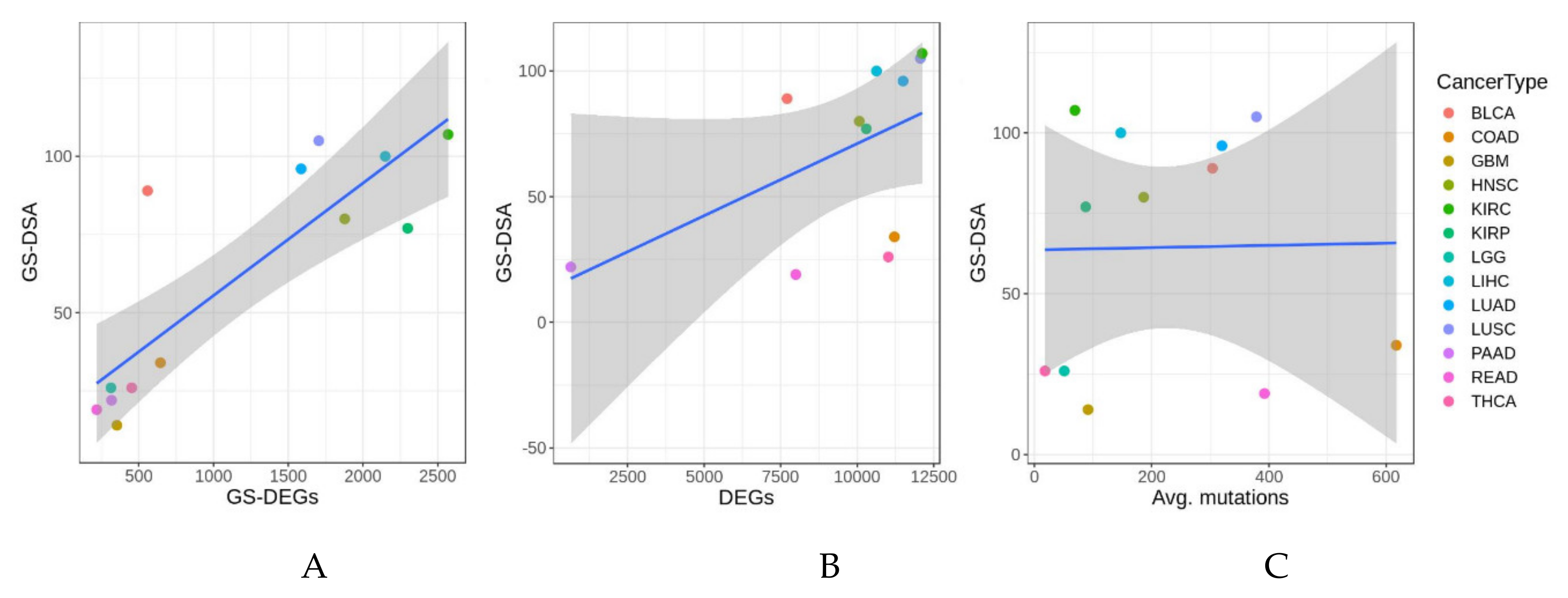
3.4. Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations (Cancer Abbreviations are Listed in Table 1):
| Abbreviation | Meaning |
| CHAT | Cancer Hallmarks Analytics Tool |
| DEG | Differentially Expressed Genes |
| FDR | False Discovery Rate |
| GS-DEG | Gender-Specific Differential Expressed Genes |
| GS-DSA | Gender-Specific Differential Signaling Activity |
| ICGC | International Cancer Genome Consortium |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| PSM | Propensity Score Matching |
| TMM | Trimmed Mean of M-values |
References
- Flanagan, K.L. Sexual dimorphism in biomedical research: A call to analyse by sex. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 385–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodruff, T. Sex, equity, and science. Proc. Natl. Acad. Sci. USA 2014, 111, 5063–5064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.L.; Schiebinger, L.; Stefanick, M.L.; Cahill, L.; Danska, J.; De Vries, G.J.; Kibbe, M.R.; McCarthy, M.M.; Mogil, J.S.; Woodruff, T.; et al. Opinion: Sex inclusion in basic research drives discovery. Proc. Natl. Acad. Sci. USA 2015, 112, 5257–5258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinn, J.L.; Snyder, M. Sexual dimorphism in mammalian gene expression. Trends Genet. 2005, 21, 298–305. [Google Scholar] [CrossRef]
- Ober, C.; Loisel, D.A.; Gilad, Y. Sex-specific genetic architecture of human disease. Nat. Rev. Genet. 2008, 9, 911–922. [Google Scholar] [CrossRef] [Green Version]
- Karp, N.A.; Mason, J.; Beaudet, A.L.; Benjamini, Y.; Bower, L.; Braun, R.E.; Brown, S.D.; Chesler, E.J.; Dickinson, M.E.; Flenniken, A.M.; et al. Prevalence of sexual dimorphism in mammalian phenotypic traits. Nat. Commun. 2017, 8, 15475. [Google Scholar] [CrossRef]
- Yoon, D.Y.; Mansukhani, N.A.; Stubbs, V.C.; Helenowski, I.B.; Woodruff, T.; Kibbe, M.R. Sex bias exists in basic science and translational surgical research. Surgery 2014, 156, 508–516. [Google Scholar] [CrossRef]
- Beery, A.K.; Zucker, I. Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2010, 35, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.S.; Reiches, M.; Shattuck-Heidorn, H.; LaBonte, M.L.; Consoli, T. Opinion: Focus on preclinical sex differences will not address women’s and men’s health disparities. Proc. Natl. Acad. Sci. USA 2015, 112, 13419–13420. [Google Scholar] [CrossRef] [Green Version]
- Kolhe, R.; Hunter, M.; Liu, S.; Jadeja, R.N.; Pundkar, C.; Mondal, A.K.; Mendhe, B.; Drewry, M.; Rojiani, M.V.; Liu, Y.; et al. Gender-specific differential expression of exosomal miRNA in synovial fluid of patients with osteoarthritis. Sci. Rep. 2017, 7, 2029. [Google Scholar] [CrossRef]
- Grissom, N.M.; E McKee, S.; Schoch, H.; Bowman, N.; Havekes, R.; O’Brien, W.T.; Mahrt, E.; Siegel, S.; Commons, K.G.; Portfors, C.; et al. Male-specific deficits in natural reward learning in a mouse model of neurodevelopmental disorders. Mol. Psychiatry 2017, 23, 544–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amadoz, A.; Hidalgo, M.; Cubuk, C.; Caballero, J.C.; Dopazo, J. A comparison of mechanistic signaling pathway activity analysis methods. Briefings Bioinform. 2019, 20, 1655–1668. [Google Scholar] [CrossRef]
- Cubuk, C.; Hidalgo, M.R.; Amadoz, A.; Rian, K.; Salavert, F.; Pujana, M.A.; Mateo, F.; Herranz, C.; Caballero, J.C.; Dopazo, J. Differential metabolic activity and discovery of therapeutic targets using summarized metabolic pathway models. bioRxiv 2018, 367334. [Google Scholar]
- Hidalgo, M.R.; Cubuk, C.; Amadoz, A.; Salavert, F.; Caballero, J.C.; Dopazo, J. High throughput estimation of functional cell activities reveals disease mechanisms and predicts relevant clinical outcomes. Oncotarget 2016, 8, 5160–5178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubuk, C.; Hidalgo, M.; Amadoz, A.; Pujana, M.A.; Mateo, F.; Herranz, C.; Caballero, J.C.; Dopazo, J. Gene Expression Integration into Pathway Modules Reveals a Pan-Cancer Metabolic Landscape. Cancer Res. 2018, 78, 6059–6072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fey, D.; Halász, M.; Dreidax, D.; Kennedy, S.P.; Hastings, J.F.; Rauch, N.; Munoz, A.G.; Pilkington, R.; Fischer, M.; Westermann, F.; et al. Signaling pathway models as biomarkers: Patient-specific simulations of JNK activity predict the survival of neuroblastoma patients. Sci. Signal. 2015, 8, ra130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, M.R.; Amadoz, A.; Cubuk, C.; Carbonell-Caballero, J.; Dopazo, J. Models of cell signaling uncover molecular mechanisms of high-risk neuroblastoma and predict disease outcome. Biol. Direct 2018, 13, 16. [Google Scholar] [CrossRef]
- Jiao, Y.; Hidalgo, M.; Cubuk, C.; Amadoz, A.; Caballero, J.C.; Vert, J.-P.; Dopazo, J. Signaling Pathway Activities Improve Prognosis for Breast Cancer. bioRxiv 2017, 132357. [Google Scholar] [CrossRef] [Green Version]
- Chacón-Solano, E.; León, C.; Díaz, F.; García-García, F.; García, M.; Escámez, M.; Guerrero-Aspizua, S.; Conti, C.; Mencía, Á.; Martínez-Santamaría, L.; et al. Fibroblast activation and abnormal extracellular matrix remodelling as common hallmarks in three cancer-prone genodermatoses. Br. J. Dermatol. 2019, 181, 512–522. [Google Scholar] [CrossRef]
- Peña-Chilet, M.; Esteban-Medina, M.; Falco, M.M.; Rian, K.; Hidalgo, M.R.; Loucera, C.; Dopazo, J. Using mechanistic models for the clinical interpretation of complex genomic variation. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Amadoz, A.; Sebastian-Leon, P.; Vidal, E.; Salavert, F.; Dopazo, J. Using activation status of signaling pathways as mechanism-based biomarkers to predict drug sensitivity. Sci. Rep. 2015, 5, 18494. [Google Scholar] [CrossRef] [Green Version]
- Esteban-Medina, M.; Peña-Chilet, M.; Loucera, C.; Dopazo, J. Exploring the druggable space around the Fanconi anemia pathway using machine learning and mechanistic models. BMC Bioinform. 2019, 20, 370. [Google Scholar] [CrossRef] [Green Version]
- Razzoli, M.; Frontini, A.; Gurney, A.; Mondini, E.; Cubuk, C.; Katz, L.S.; Cero, C.; Bolan, P.J.; Dopazo, J.; Vidal-Puig, A.; et al. Stress-induced activation of brown adipose tissue prevents obesity in conditions of low adaptive thermogenesis. Mol. Metab. 2015, 5, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.; Muñoz-Aguirre, M.; Reverter, F.; Godinho, C.P.S.; Sousa, A.; Amadoz, A.; Sodaei, R.; Hidalgo, M.R.; Pervouchine, D.; Carbonell-Caballero, J.; et al. The effects of death and post-mortem cold ischemia on human tissue transcriptomes. Nat. Commun. 2018, 9, 490. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; A Weinberg, R. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salavert, F.; Hidago, M.R.; Amadoz, A.; Cubuk, C.; Medina, I.; Crespo, D.; Carbonell-Caballero, J.; Dopazo, J. Actionable pathways: Interactive discovery of therapeutic targets using signaling pathway models. Nucleic Acids Res. 2016, 44, W212–W216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falco, M.M.; Pena-Chilet, M.; Loucera, C.; Hidalgo, M.; Dopazo, J. Mechanistic models of signaling pathways deconvolute the functional landscape of glioblastoma at single cell resolution. bioRxiv 2019, 858811. [Google Scholar] [CrossRef] [Green Version]
- Ho, D.E.; Imai, K.; King, G.; Stuart, E.A. Matching as Nonparametric Preprocessing for Reducing Model Dependence in Parametric Causal Inference. Polit. Anal. 2007, 15, 199–236. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 2006, 8, 118–127. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: Back to metabolism in KEGG. Nucleic Acids Res. 2013, 42, D199–D205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastian-Leon, P.; Vidal, E.; Minguez, P.; Conesa, A.; Tarazona, S.; Amadoz, A.; Armero, C.; Salavert, F.; Vidal-Puig, A.; Montaner, D.; et al. Understanding disease mechanisms with models of signaling pathway activities. BMC Syst. Biol. 2014, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Efroni, S.; Schaefer, C.F.; Buetow, K.H. Identification of Key Processes Underlying Cancer Phenotypes Using Biologic Pathway Analysis. PLoS ONE 2007, 2, e425. [Google Scholar] [CrossRef] [Green Version]
- Montaner, D.; Minguez, P.; Al-Shahrour, F.; Dopazo, J. Gene set internal coherence in the context of functional profiling. BMC Genom. 2009, 10, 197. [Google Scholar] [CrossRef] [Green Version]
- UniProt Consortium. The UniProt Consortium UniProt: A hub for protein information. Nucleic Acids Res. 2014, 43, D204–D212. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium; Carbon, S.; Douglass, E.; Dunn, N.; Good, B.; Harris, N.L.; Lewis, S.E.; Mungall, C.J.; Basu, S.; Chisholm, R.L.; et al. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2018, 47, D330–D338. [Google Scholar] [CrossRef] [Green Version]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1–33. [Google Scholar] [CrossRef]
- Baker, S.; Ali, I.; Silins, I.; Pyysalo, S.; Guo, Y.; Högberg, J.; Stenius, U.; Korhonen, A. Cancer Hallmarks Analytics Tool (CHAT): A text mining approach to organize and evaluate scientific literature on cancer. Bioinformatics 2017, 33, 3973–3981. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, L.; Chen, H.; Wang, Y.; Xu, Y.; Mao, H.; Li, J.; Mills, G.B.; Shu, Y.; Li, L.; et al. Comprehensive Characterization of Molecular Differences in Cancer between Male and Female Patients. Cancer Cell 2016, 29, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Assessment Report MVASI. Available online: https://www.ema.europa.eu/en/documents/assessment-report/mvasi-epar-public-assessment-report_en.pdf (accessed on 20 June 2020).
- CHPM Assessment Report cabometyx. Available online: https://www.ema.europa.eu/en/documents/assessment-report/cabometyx-epar-public-assessment-report_en.pdf (accessed on 20 June 2020).
- Assessment Report for iressa. Available online: https://www.ema.europa.eu/en/documents/assessment-report/iressa-epar-public-assessment-report_en.pdf (accessed on 20 June 2020).
- Assessment Report for tyverb. Available online: https://www.ema.europa.eu/en/documents/assessment-report/tyverb-epar-public-assessment-report_en.pdf (accessed on 20 June 2020).
- Flagg, P.J. Scientific Discussions. Anesthesiology 1942, 3, 336–337. [Google Scholar] [CrossRef]
- CHMP Assessment Report ruxolitinib. Available online: https://www.ema.europa.eu/en/documents/assessment-report/jakavi-epar-public-assessment-report_en.pdf (accessed on 20 June 2020).
- CHMP Extension of Indication Variation Assessment Report. Available online: https://www.ema.europa.eu/en/documents/variation-report/nexavar-h-c-690-ii-35-epar-assessment-report-variation_en.pdf (accessed on 20 June 2020).
- Segarra, I.; Modamio, P.; Fernández, C.; Mariño, E.L. Sex-Divergent Clinical Outcomes and Precision Medicine: An Important New Role for Institutional Review Boards and Research Ethics Committees. Front. Pharmacol. 2017, 8, 488. [Google Scholar] [CrossRef] [PubMed]
- CHMP Assessment Report trametinib. Available online: https://www.ema.europa.eu/en/documents/assessment-report/mekinist-epar-public-assessment-report_en.pdf (accessed on 20 June 2020).
- Assessment Report Zelboraf. Available online: https://www.ema.europa.eu/en/documents/assessment-report/zelboraf-epar-public-assessment-report_en.pdf (accessed on 20 June 2020).
- Assessment Report Odomzo. Available online: https://www.ema.europa.eu/en/documents/assessment-report/odomzo-epar-public-assessment-report_en.pdf (accessed on 20 June 2020).
- Ye, Y.; Jing, Y.; Li, L.; Mills, G.B.; Diao, L.; Liu, H.; Han, L. Sex-associated molecular differences for cancer immunotherapy. Nat. Commun. 2020, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cubuk, C.; Hidalgo, M.R.; Amadoz, A.; Rian, K.; Salavert, F.; Pujana, M.A.; Mateo, F.; Herranz, C.; Carbonell-Caballero, J.; Dopazo, J. Differential metabolic activity and discovery of therapeutic targets using summarized metabolic pathway models. NPJ Syst. Biol. Appl. 2019, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-I.; Lim, H.; Moon, A. Sex Differences in Cancer: Epidemiology, Genetics and Therapy. Biomol. Ther. 2018, 26, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Dorak, M.T.; Karpuzoglu, E. Gender Differences in Cancer Susceptibility: An Inadequately Addressed Issue. Front. Genet. 2012, 3, 268. [Google Scholar] [CrossRef] [Green Version]
- Özdemir, B.C.; Csajka, C.; Dotto, G.-P.; Wagner, A.D. Sex Differences in Efficacy and Toxicity of Systemic Treatments: An Undervalued Issue in the Era of Precision Oncology. J. Clin. Oncol. 2018, 36, 2680–2683. [Google Scholar] [CrossRef]
- A Pinto, J.; Vallejos, C.S.; E Raez, L.; A Mas, L.; Ruiz, R.; Torres-Roman, J.S.; Morante, Z.; Araujo, J.M.; Gomez, H.L.; Aguilar, A.; et al. Gender and outcomes in non-small cell lung cancer: An old prognostic variable comes back for targeted therapy and immunotherapy? ESMO Open 2018, 3, e000344. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Cowley, L.A.; Liu, X.-S. Sex Differences in Cancer Immunotherapy Efficacy, Biomarkers, and Therapeutic Strategy. Molecules 2019, 24, 3214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niedernhofer, L.J.; Lalai, A.S.; Hoeijmakers, J.H. Fanconi Anemia (Cross)linked to DNA Repair. Cell 2005, 123, 1191–1198. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Barzilla, J.E.; Mendez, J.S.; Stephens, E.H.; Lee, E.L.; Collard, C.D.; Laucirica, R.; Weigel, P.H.; Grande-Allen, K.J. Abundance and location of proteoglycans and hyaluronan within normal and myxomatous mitral valves. Cardiovasc. Pathol. 2008, 18, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-H.; Kim, Y.K.; Jung, J.-Y.; Shin, J.-E.; Chung, J.H. Changes in glycosaminoglycans and related proteoglycans in intrinsically aged human skin in vivo. Exp. Dermatol. 2011, 20, 454–456. [Google Scholar] [CrossRef]
- Levin, E.R. Bidirectional Signaling between the Estrogen Receptor and the Epidermal Growth Factor Receptor. Mol. Endocrinol. 2003, 17, 309–317. [Google Scholar] [CrossRef]
- Bonaccorsi, L. The androgen receptor associates with the epidermal growth factor receptor in androgen-sensitive prostate cancer cells. Steroids 2004, 69, 549–552. [Google Scholar] [CrossRef]
- Carter, C.S. The Oxytocin–Vasopressin Pathway in the Context of Love and Fear. Front. Endocrinol. 2017, 8, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Fitzpatrick, L.; Ruan, M.; Anderson, J.; Moraghan, T.; Miller, V. Gender-related differences in vascular smooth muscle cell proliferation: Implications for prevention of arteriosclerosis. Lupus 1999, 8, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Kerksick, C.M.; Taylor, L.; Harvey, A.; Willoughby, D. Gender-Related Differences in Muscle Injury, Oxidative Stress, and Apoptosis. Med. Sci. Sports Exerc. 2008, 40, 1772–1780. [Google Scholar] [CrossRef] [PubMed]
- Fossett, E.; Khan, W.S.; Longo, U.G.; Smitham, P. Effect of age and gender on cell proliferation and cell surface characterization of synovial fat pad derived mesenchymal stem cells. J. Orthop. Res. 2012, 30, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Mallat, Z.; Fornes, P.; Costagliola, R.; Esposito, B.; Belmin, J.; LeComte, D.; Tedgui, A. Age and gender effects on cardiomyocyte apoptosis in the normal human heart. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2001, 56, M719–M723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yan, S.; Liu, X.; Gan, L.; Wu, Z.; Gong, Y.; Huang, M.; Zhang, X.; Guo, W. Gender-related prognostic value and genomic pattern of intra-tumor heterogeneity in colorectal cancer. Carcinogenesis 2017, 38, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Köglsberger, S.; Cordero-Maldonado, M.L.; Antony, P.; Forster, J.I.; Garcia, P.; Buttini, M.; Crawford, A.; Glaab, E. Gender-Specific Expression of Ubiquitin-Specific Peptidase 9 Modulates Tau Expression and Phosphorylation: Possible Implications for Tauopathies. Mol. Neurobiol. 2016, 54, 7979–7993. [Google Scholar] [CrossRef] [Green Version]
- Li, C.H.; Haider, S.; Shiah, Y.-J.; Thai, K.; Boutros, P.C. Sex Differences in Cancer Driver Genes and Biomarkers. Cancer Res. 2018, 78, 5527–5537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, I.; Högberg, J.; Hsieh, J.-H.; Auerbach, S.; Korhonen, A.; Stenius, U.; Silins, I. Gender differences in cancer susceptibility: Role of oxidative stress. Carcinogenesis 2016, 37, 985–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, K.E.; Riddle, N.C. Sex Differences in Aging: Genomic Instability. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2018, 73, 166–174. [Google Scholar] [CrossRef]
- Jensen, M.D. Gender differences in regional fatty acid metabolism before and after meal ingestion. J. Clin. Investig. 1995, 96, 2297–2303. [Google Scholar] [CrossRef] [Green Version]
- Childs, C.E.; Romeu-Nadal, M.; Burdge, G.C.; Calder, P.C. Gender differences in the n-3 fatty acid content of tissues. Proc. Nutr. Soc. 2008, 67, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittendorfer, B. Sexual Dimorphism in Human Lipid Metabolism. J. Nutr. 2005, 135, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.R.; Schulze, A. Lipid metabolism in cancer. FEBS J. 2012, 279, 2610–2623. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Zhang, C.-J.; Zhu, N.; Du, K.; Yin, Y.-F.; Tan, X.; Liao, D.-F.; Qin, L. Lipid metabolism and carcinogenesis, cancer development. Am. J. Cancer Res. 2018, 8, 778–791. [Google Scholar]
- Munir, R.; Lisec, J.; Swinnen, J.V.; Zaidi, N. Lipid metabolism in cancer cells under metabolic stress. Br. J. Cancer 2019, 120, 1090–1098. [Google Scholar] [CrossRef]
- Ricketts, C.J.; Linehan, W.M. Gender Specific Mutation Incidence and Survival Associations in Clear Cell Renal Cell Carcinoma (CCRCC). PLoS ONE 2015, 10, e0140257. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.-H.; Chiang, C.-J.; Tseng, J.-S.; Yang, T.-Y.; Hsu, K.-H.; Chen, K.-C.; Wang, C.-L.; Chen, C.-Y.; Yen, S.-H.; Tsai, C.-M.; et al. EGFR mutation, smoking, and gender in advanced lung adenocarcinoma. Oncotarget 2017, 8, 98384–98393. [Google Scholar] [CrossRef] [Green Version]
- Andersson, G.; Wennersten, C.; Borgquist, S.; Jirström, K. Pancreatic cancer risk in relation to sex, lifestyle factors, and pre-diagnostic anthropometry in the Malmö Diet and Cancer Study. Biol. Sex. Differ. 2016, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of Pancreatic Cancer: Global Trends, Etiology and Risk Factors. World J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Haake, S.M.; Brannon, A.R.; Hacker, K.; Pruthi, R.; Wallen, E.; Nielsen, M.E.; Rathmell, K. Use of meta-analysis of clear cell renal cell carcinoma gene expression to define a variant subgroup and identify gender influences on tumor biology. J. Clin. Oncol. 2012, 30, 412. [Google Scholar] [CrossRef]
- Lee, S.H.; Oh, S.-Y.; I Do, S.; Lee, H.J.; Kang, H.J.; Rho, Y.S.; Bae, W.J.; Lim, Y.C. SOX2 regulates self-renewal and tumorigenicity of stem-like cells of head and neck squamous cell carcinoma. Br. J. Cancer 2014, 111, 2122–2130. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Wu, J.; Lai, S.; Xu, Y.; Zhan, J.; Li, R.; Liu, X.; Wang, N.; Wei, X.; Jiang, X.; et al. Clear cell renal cell carcinoma: The value of sex-specific abdominal visceral fat measured on CT for prediction of Fuhrman nuclear grade. Eur. Radiol. 2020, 1–10. [Google Scholar] [CrossRef]
- Cui, C.; Yang, W.; Shi, J.; Zhou, Y.; Yang, J.; Cui, Q.; Zhou, Y. Identification and Analysis of Human Sex-biased MicroRNAs. Genom. Proteom. Bioinform. 2018, 16, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Morgan, M.; Hutchison, K.; Calhoun, V.D. A Study of the Influence of Sex on Genome Wide Methylation. PLoS ONE 2010, 5, e10028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boks, M.P.M.; Derks, E.M.; Weisenberger, D.J.; Strengman, E.; Janson, E.; Sommer, I.E.C.; Kahn, R.S.; Ophoff, R.A. The Relationship of DNA Methylation with Age, Gender and Genotype in Twins and Healthy Controls. PLoS ONE 2009, 4, e6767. [Google Scholar] [CrossRef] [Green Version]
- O’Brown, Z.K.; Van Nostrand, E.; Higgins, J.P.; Kim, S.K. The Inflammatory Transcription Factors NFκB, STAT1 and STAT3 Drive Age-Associated Transcriptional Changes in the Human Kidney. PLoS Genet. 2015, 11, e1005734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.; Dong, G.; Zhao, G.; Liu, Z.; Yao, G.; Zhu, Y.; Hou, Y. Gender Differences of B Cell Signature in Healthy Subjects Underlie Disparities in Incidence and Course of SLE Related to Estrogen. J. Immunol. Res. 2014, 2014, 1–17. [Google Scholar] [CrossRef]
- Atanaskova, N.; Keshamouni, V.; Krueger, J.S.; A Schwartz, J.; Miller, F.; Reddy, K. MAP kinase/estrogen receptor cross-talk enhances estrogen-mediated signaling and tumor growth but does not confer tamoxifen resistance. Oncogene 2002, 21, 4000–4008. [Google Scholar] [CrossRef] [Green Version]
- Driggers, P.H.; Segars, J. Estrogen action and cytoplasmic signaling pathways. Part II: The role of growth factors and phosphorylation in estrogen signaling. Trends Endocrinol. Metab. 2002, 13, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Guillot, R.; Muriach, B.; Rocha, A.; Rotllant, J.; Kelsh, R.N.; Cerdá-Reverter, J.M. Thyroid Hormones Regulate Zebrafish Melanogenesis in a Gender-Specific Manner. PLoS ONE 2016, 11, e0166152. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Cadenas, C.; Peña-Chilet, M.; Ibarrola-Villava, M.; Ribas, G. Gender is a major factor explaining discrepancies in eye colour prediction based on HERC2/OCA2 genotype and the IrisPlex model. Forensic Sci. Int. Genet. 2013, 7, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Pietroni, C.; Andersen, M.M.; Johansen, P.; Andersen, M.M.; Harder, S.; Paulsen, R.R.; Børsting, C.; Morling, N. The effect of gender on eye colour variation in European populations and an evaluation of the IrisPlex prediction model. Forensic Sci. Int. Genet. 2014, 11, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernando, B.; Ibarrola-Villava, M.; Fernández, L.P.; Pena-Chilet, M.; Llorca-Cardeñosa, M.; Oltra, S.S.; Alonso, S.; Boyano, M.D.; Cadenas, C.M.; Ribas, G. Sex-specific genetic effects associated with pigmentation, sensitivity to sunlight, and melanoma in a population of Spanish origin. Biol. Sex. Differ. 2016, 7, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, M.J. The biological actions of estrogens on skin. Exp. Dermatol. 2002, 11, 487–502. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.-Y. Recent progress in melasma pathogenesis. Pigment. Cell Melanoma Res. 2015, 28, 648–660. [Google Scholar] [CrossRef]
- Hohla, F.; Hopfinger, G.; Romeder, F.; Rinnerthaler, G.; Bezan, A.; Stättner, S.; Hauser-Kronberger, C.; Ulmer, H.; Greil, R. Female gender may predict response to FOLFIRINOX in patients with unresectable pancreatic cancer: A single institution retrospective review. Int. J. Oncol. 2013, 44, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Sebastian-Leon, P.; Carbonell, J.; Salavert, F.; Sanchez, R.; Medina, I.; Dopazo, J. Inferring the functional effect of gene expression changes in signaling pathways. Nucleic Acids Res. 2013, 41, W213–W217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsos, A.; Melas, I.; Siminelakis, P.; Chairakaki, A.D.; Saez-Rodriguez, J.; Alexopoulos, L.G. Identifying Drug Effects via Pathway Alterations using an Integer Linear Programming Optimization Formulation on Phosphoproteomic Data. PLoS Comput. Biol. 2009, 5, e1000591. [Google Scholar] [CrossRef] [Green Version]
- Fryburg, D.A.; Song, D.H.; Laifenfeld, D.; De Graaf, D. Systems diagnostics: Anticipating the next generation of diagnostic tests based on mechanistic insight into disease. Drug Discov. Today 2014, 19, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Dopazo, J. Genomics and transcriptomics in drug discovery. Drug Discov. Today 2014, 19, 126–132. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Code | Cancer Type | Female | Male | Sample Size | Proportion (Male/Female) |
|---|---|---|---|---|---|
| BLCA | Bladder urothelial carcinoma | 57 | 202 | 259 | 3.54 |
| COAD | Colon adenocarcinoma | 113 | 207 | 320 | 1.83 |
| GBM | Brain Glioblastoma Multiforme | 37 | 89 | 126 | 2.41 |
| HNSC | Head and Neck squamous cell carcinoma | 97 | 328 | 425 | 3.38 |
| KIRC | Kidney renal clear cell carcinoma | 124 | 314 | 438 | 2.53 |
| KIRP | Kidney renal papillary cell carcinoma | 38 | 108 | 146 | 2.84 |
| LGG | Brain Lower Grade Glioma | 104 | 205 | 309 | 1.97 |
| LIHC | Liver hepatocellular carcinoma | 44 | 118 | 162 | 2.68 |
| LUAD | Lung adenocarcinoma | 131 | 213 | 344 | 1.63 |
| LUSC | Lung squamous cell carcinoma | 81 | 299 | 380 | 3.69 |
| PAAD | Pancreatic Cancer | 30 | 77 | 107 | 2.57 |
| READ | Rectum adenocarcinoma | 41 | 77 | 118 | 1.88 |
| THCA | Thyroid Carcinoma | 66 | 127 | 193 | 1.92 |
| Total | 963 | 2364 | 3327 |
| Cancer Type | Cancer | Cancer (M > F) | Cancer (F > M) | Drug Simulation | Drug (M > F) | Drug (F > M) | Drug Diff. Cancer |
|---|---|---|---|---|---|---|---|
| GBM | 43 | 21 | 22 | 50 | 24 | 26 | 14 |
| READ | 22 | 5 | 17 | 34 | 13 | 21 | 19 |
| PAAD | 31 | 15 | 16 | 48 | 24 | 24 | 22 |
| LGG | 59 | 22 | 37 | 75 | 31 | 44 | 26 |
| THCA | 52 | 12 | 40 | 61 | 21 | 40 | 26 |
| COAD | 42 | 18 | 24 | 61 | 13 | 48 | 34 |
| KIRP | 145 | 57 | 88 | 180 | 78 | 102 | 77 |
| HNSC | 202 | 66 | 136 | 242 | 78 | 164 | 80 |
| BLCA | 104 | 56 | 48 | 161 | 92 | 69 | 89 |
| LUAD | 203 | 50 | 153 | 242 | 53 | 189 | 96 |
| LIHC | 168 | 36 | 132 | 212 | 35 | 177 | 100 |
| LUSC | 224 | 105 | 119 | 238 | 112 | 126 | 105 |
| KIRC | 239 | 98 | 141 | 301 | 141 | 160 | 107 |
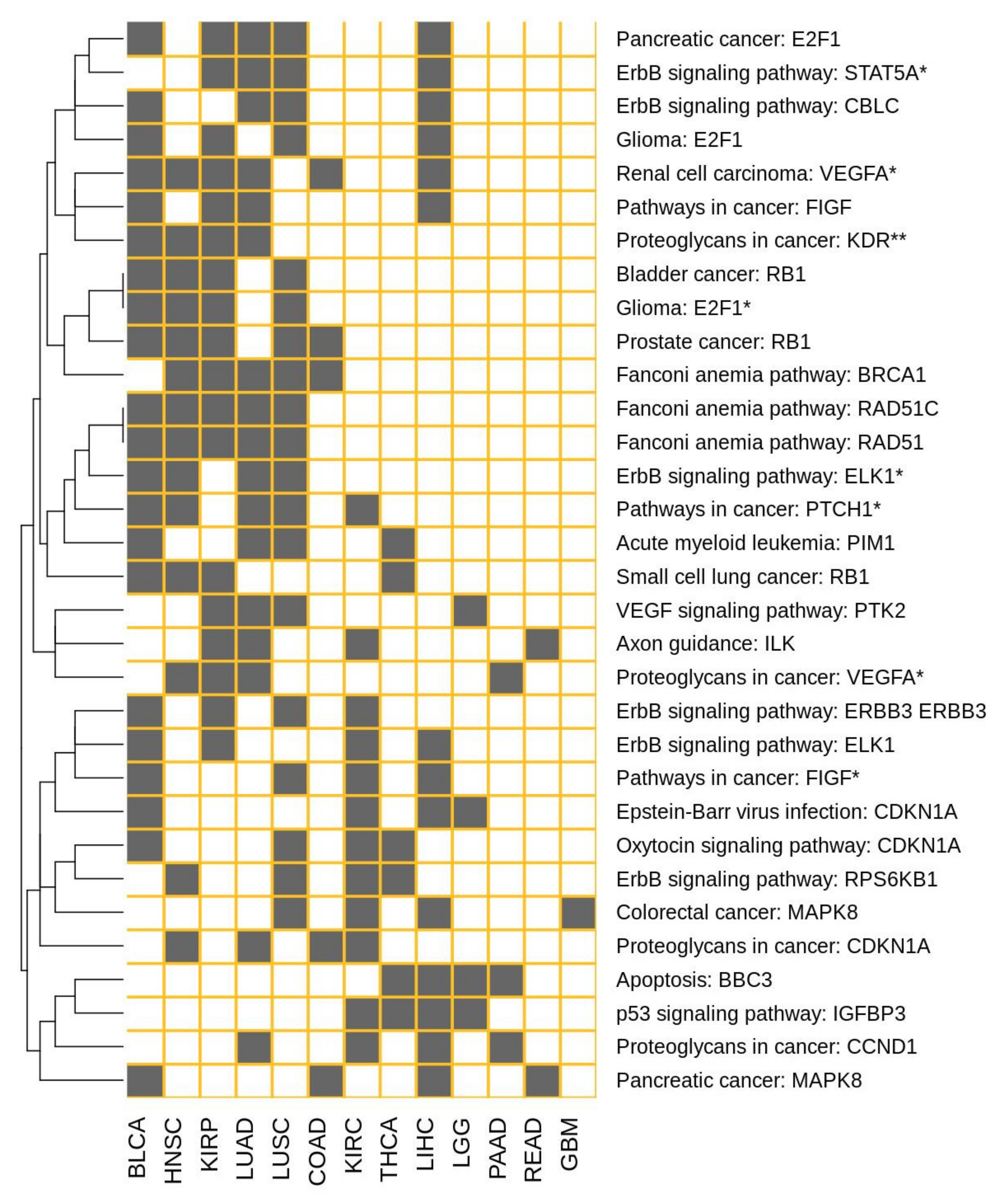
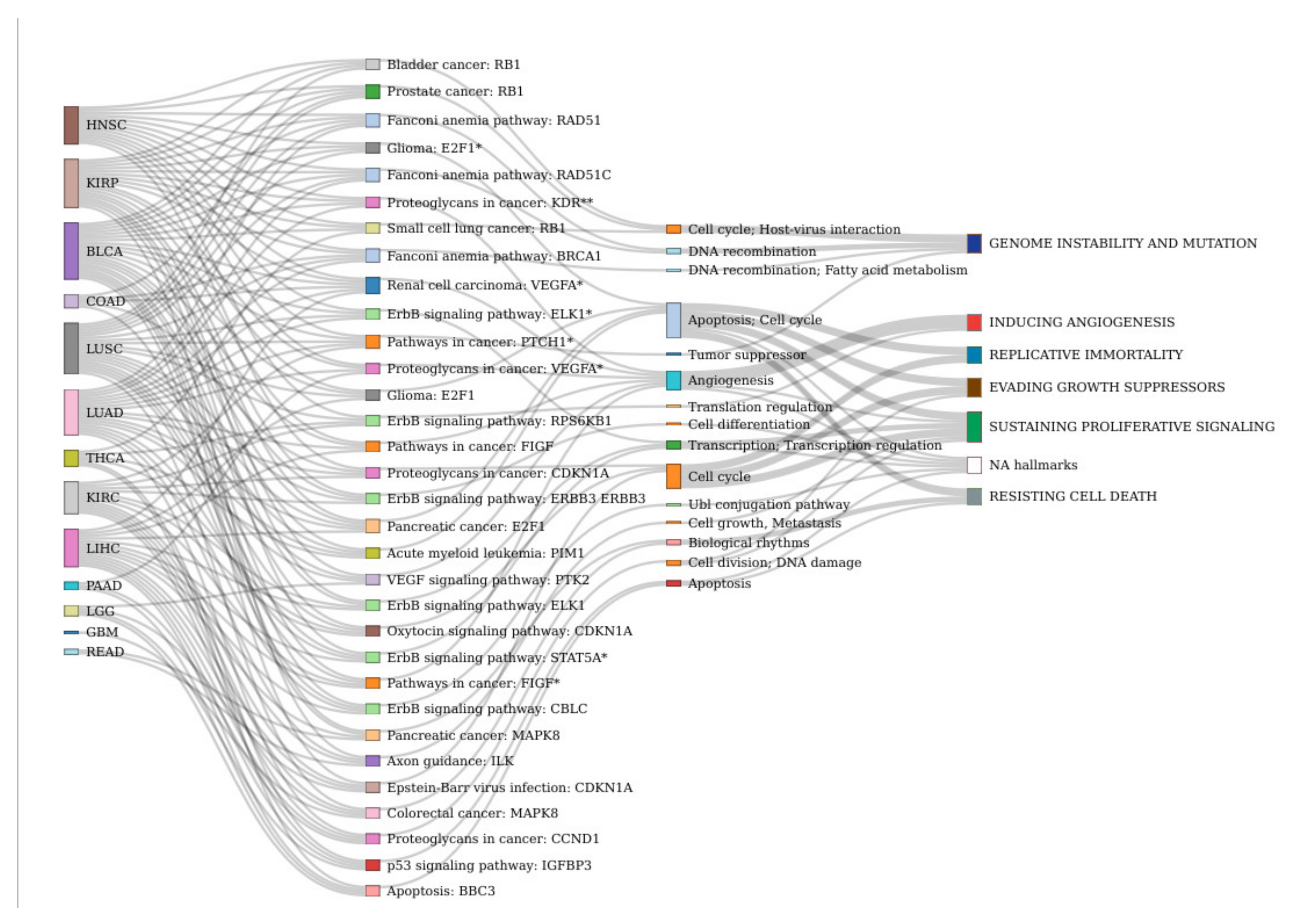
| Effector Circuit | Uniprot Annotation of Effector Circuits | Cancers with GS-DSA |
|---|---|---|
| Renal cell carcinoma: VEGFA * | Angiogenesis | BLCA, COAD, HNSC, KIRP, LIHC, LUAD |
| Fanconi anemia pathway: RAD51 | DNA recombination | BLCA, HNSC, KIRP, LUAD, LUSC |
| Fanconi anemia pathway: RAD51C | DNA recombination | BLCA, HNSC, KIRP, LUAD, LUSC |
| Fanconi anemia pathway: BRCA1 | DNA recombination; | COAD, HNSC, KIRP, LUAD, LUSC |
| Pathways in cancer: PTCH1 * | Tumor suppressor | BLCA, HNSC, KIRC, LUAD, LUSC |
| Pancreatic cancer: E2F1 | Apoptosis; Cell cycle | BLCA, KIRP, LIHC, LUAD, LUSC |
| Prostate cancer: RB1 | Cell cycle | BLCA, COAD, HNSC, KIRP, LUSC |
| ErbB signaling pathway: RPS6KB1 | Translation regulation | HNSC, KIRC, LUSC, THCA |
| ErbB signaling pathway: ELK1 | Transcription; Transcription regulation | BLCA, KIRC, KIRP, LIHC |
| ErbB signaling pathway: STAT5A * | Transcription; Transcription regulation | KIRP, LIHC, LUAD, LUSC |
| ErbB signaling pathway: ELK1 * | Transcription; Transcription regulation | BLCA, HNSC, LUAD, LUSC |
| ErbB signaling pathway: CBLC | Ubl conjugation pathway | BLCA, LIHC, LUAD, LUSC |
| ErbB signaling pathway: ERBB3 ERBB3 | Cell differentiation | BLCA, KIRC, KIRP, LUSC |
| p53 signaling pathway: IGFBP3 | Apoptosis | KIRC, LGG, LIHC, THCA |
| Apoptosis: BBC3 | Apoptosis | LGG, LIHC, PAAD, THCA |
| Axon guidance: ILK | Cell growth, Metastasis | KIRC, KIRP, LUAD, READ |
| VEGF signaling pathway: PTK2 | Angiogenesis | KIRP, LGG, LUAD, LUSC |
| Oxytocin signaling pathway: CDKN1A | Cell cycle | BLCA, KIRC, LUSC, THCA |
| Pathways in cancer: FIGF | Angiogenesis | BLCA, KIRP, LIHC, LUAD |
| Pathways in cancer: FIGF * | Angiogenesis | BLCA, KIRC, LIHC, LUSC |
| Proteoglycans in cancer: CCND1 | Cell division; DNA damage | KIRC, LIHC, LUAD, PAAD |
| Proteoglycans in cancer: CDKN1A | Cell cycle | COAD, HNSC, KIRC, LUAD |
| Proteoglycans in cancer: VEGFA * | Angiogenesis | HNSC, KIRP, LUAD, PAAD |
| Proteoglycans in cancer: KDR ** | Angiogenesis | BLCA, HNSC, KIRP, LUAD |
| Colorectal cancer: MAPK8 | Biological rhythms | GBM, KIRC, LIHC, LUSC |
| Pancreatic cancer: MAPK8 | Biological rhythms | BLCA, COAD, LIHC, READ |
| Glioma: E2F1 | Apoptosis; Cell cycle | BLCA, KIRP, LIHC, LUSC |
| Glioma: E2F1 * | Apoptosis; Cell cycle | BLCA, HNSC, KIRP, LUSC |
| Bladder cancer: RB1 | Cell cycle | BLCA, HNSC, KIRP, LUSC |
| Acute myeloid leukemia: PIM1 | Apoptosis; Cell cycle | BLCA, LUAD, LUSC, THCA |
| Small cell lung cancer: RB1 | Cell cycle | BLCA, HNSC, KIRP, THCA |
| Pathway | Effector | Bevacizumab | Cabozantinib | Gefitinib | Lapatinib | Nilotinib | Ruxolitinib | Sorafenib | Sunitinib | Trametinib | Vemurafenib | Sonidegib |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ras signaling pathway | BRAP | . | . | . | . | . | . | . | . | . | Y | . |
| cGMP-PKG signaling pathway | MAPK1 | . | . | . | . | . | . | Y | . | . | . | . |
| cAMP signaling pathway | MYL9, PTCH1, HHIP, ACOX1, F2R AMH, ORAI1, BAD, NFKBIA NFKB1, RYR2, GRIN3A, GRIA1, CFTR, SLC9A1, ATP2B1, CACNA1C, PDE3A, ATP1B4 FXYD1, RHOA, C00165, C01245, PAK1, MLLT4, C00416, MAPK8, HCN4 | . | . | . | . | . | Y | . | . | . | . | . |
| Chemokine signaling pathway | STAT1 | . | . | . | . | . | Y | . | . | . | . | . |
| Wnt signaling pathway | JUN | . | . | Y | . | . | . | . | . | . | . | . |
| Hedgehog signaling pathway | PTCH1, SMO, PTCH1, GLI1, HHIP, CCND1, BCL2, PRKACA, GLI1 SUFU, | . | . | . | . | . | . | . | . | . | . | Y |
| Axon guidance | ILK | . | . | . | . | . | . | . | . | . | . | Y |
| VEGF signaling pathway: | NOS3 | Y | . | . | . | . | . | . | . | . | . | . |
| Osteoclast differentiation: | MAPK1 | . | . | . | . | . | . | . | Y | . | . | . |
| Osteoclast differentiation: | NFKB1 | . | . | . | . | Y | . | . | . | . | . | . |
| Signaling pathways regulating pluripotency of stem cells | HNF1A | . | . | Y | . | . | . | . | . | . | . | . |
| Jak-STAT signaling pathway | BCL2, BCL2L1, MYC, AOX1, GFAP, MCL1, PIM1, CCND1 | . | . | . | . | . | Y | . | . | . | . | . |
| Natural killer cell mediated cytotoxicity | TNF | . | . | . | . | . | . | . | . | Y | . | . |
| TNF signaling pathway | CASP7, JUN, CEBPB | . | . | . | . | . | . | . | . | Y | . | . |
| Leukocyte transendothelial migration | MAPK14 | . | . | . | . | Y | . | . | . | . | . | . |
| Inflammatory mediator regulation of TRP channels: | TRPM8, TRPV4 | . | . | . | . | . | Y | . | . | . | . | . |
| Ovarian steroidogenesis | STAR, HSD3B1, PLA2G4B, ACOT2, CYP19A1, HSD17B2, CYP19A1 | . | . | . | . | . | Y | . | . | . | . | . |
| Melanogenesis | MITF | . | . | . | . | . | Y | . | . | . | . | . |
| Thyroid hormone synthesis | TG | . | . | . | . | . | Y | . | . | . | . | . |
| Thyroid hormone signaling pathway | STAT1, ESR1, THRB | . | . | . | . | . | . | . | . | Y | . | . |
| Adipocytokine signaling pathway | AGRP, NPY, POMC, PPARGC1A, PTPN11 | . | . | . | . | . | Y | . | . | . | . | . |
| Regulation of lipolysis in adipocytes | PLIN1, LIPE | . | . | . | . | . | Y | . | . | . | . | . |
| Aldosterone synthesis and secretion | CYP11B2 | . | . | . | . | . | Y | . | . | . | . | . |
| AGE-RAGE signaling pathway in diabetic complications | FOXO1, CCND1, NFATC1 | . | . | . | . | Y | . | . | . | . | . | . |
| Pathways in cancer | CCND1 | . | . | . | . | . | . | . | . | . | Y | . |
| Pathways in cancer | FIGF | Y | . | . | . | . | . | . | . | . | . | . |
| Pathways in cancer | CCNA1, CSF3R, CSF2RA, CSF1R | . | . | . | . | . | . | . | . | . | . | . |
| Pathways in cancer | CSF1R | . | . | . | . | . | . | . | Y | . | . | . |
| Pathways in cancer | BMP2, GLI1, HHIP, PTCH1 | . | . | . | . | . | . | . | . | . | . | Y |
| Proteoglycans in cancer | HSPB2 | . | . | . | . | Y | . | . | . | . | . | . |
| Proteoglycans in cancer: | AKT3 | . | Y | Y | . | . | . | . | . | . | . | . |
| Proteoglycans in cancer: | PRKCA | . | . | . | . | . | . | Y | . | . | . | |
| Colorectal cancer: | MAPK8 | . | . | . | . | . | . | . | . | Y | . | . |
| Renal cell carcinoma | VEGFA | Y | . | . | . | . | . | . | . | . | . | . |
| Renal cell carcinoma | RAP1A | . | Y | . | . | . | . | . | . | . | . | |
| Renal cell carcinoma | AKT3 | . | Y | Y | . | . | . | . | . | . | . | . |
| Pancreatic cancer | RAC1 | . | . | . | . | . | . | . | . | Y | . | . |
| Pancreatic cancer | C00416 | . | . | . | . | . | . | . | . | Y | . | . |
| Basal cell carcinoma | PTCH1 | . | . | . | . | . | . | . | . | . | Y | |
| Acute myeloid leukemia | CCNA1, SPI1 | . | . | . | . | . | . | . | . | . | . | . |
| Non-small cell lung cancer | FOXO3 | . | . | . | Y | . | . | . | . | . | . | . |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çubuk, C.; Can, F.E.; Peña-Chilet, M.; Dopazo, J. Mechanistic Models of Signaling Pathways Reveal the Drug Action Mechanisms behind Gender-Specific Gene Expression for Cancer Treatments. Cells 2020, 9, 1579. https://doi.org/10.3390/cells9071579
Çubuk C, Can FE, Peña-Chilet M, Dopazo J. Mechanistic Models of Signaling Pathways Reveal the Drug Action Mechanisms behind Gender-Specific Gene Expression for Cancer Treatments. Cells. 2020; 9(7):1579. https://doi.org/10.3390/cells9071579
Chicago/Turabian StyleÇubuk, Cankut, Fatma E. Can, María Peña-Chilet, and Joaquín Dopazo. 2020. "Mechanistic Models of Signaling Pathways Reveal the Drug Action Mechanisms behind Gender-Specific Gene Expression for Cancer Treatments" Cells 9, no. 7: 1579. https://doi.org/10.3390/cells9071579
APA StyleÇubuk, C., Can, F. E., Peña-Chilet, M., & Dopazo, J. (2020). Mechanistic Models of Signaling Pathways Reveal the Drug Action Mechanisms behind Gender-Specific Gene Expression for Cancer Treatments. Cells, 9(7), 1579. https://doi.org/10.3390/cells9071579