The WAVE Regulatory Complex Is Required to Balance Protrusion and Adhesion in Migration


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Transgenic Mice and Isolation of Nckap1fl/fl Mouse Embryonic Fibroblasts
2.2. Generation of Nckap1 KO B16-F1 Cells
2.3. Mammalian Cell Culture Conditions
2.4. Transfection of Mammalian Cells Lines
2.5. Genetic Knockouts
2.6. Analytical PCR
2.7. Cell Proliferation and Viability
2.8. SDS-PAGE and Western Blotting
2.9. Immunofluorescence Analysis
2.10. Lamellipodia Formation
2.11. xCELLigence
2.12. Focal Adhesions
2.13. Focal Adhesions Turnover
2.14. Actin Photoactivation—Retrograde Flow
2.15. Stress Fiber Quantification
- run(“Z Project...”,“projection = [Max Intensity]”);
- run(“Difference of Gaussians”, “ sigma1 = 4 sigma2 = 0.50 enhance”);
- setOption(“ScaleConversions”, true);
- run(“8-bit”);
- setAutoThreshold(“Default dark”);
- setThreshold(40, 255);
- setThreshold(40, 255);
- //setThreshold(40, 255);
- setOption(“BlackBackground”, true);
- run(“Convert to Mask”);
- run(“Ridge Detection”, “line_width = 3.5 high_contrast = 230 low_contrast = 87 estimate_width extend_line displayresults add_to_manager method_for_overlap_resolution = NONE sigma = 1.51 lower_threshold = 3.06 upper_threshold = 7.99 minimum_line_length = 15 maximum = 0”);
2.16. Rac1 Photoactivation
2.17. Random Migration Assay
2.18. Pseudopodia Quantifications
2.19. Statistics and Reproducibility
3. Results
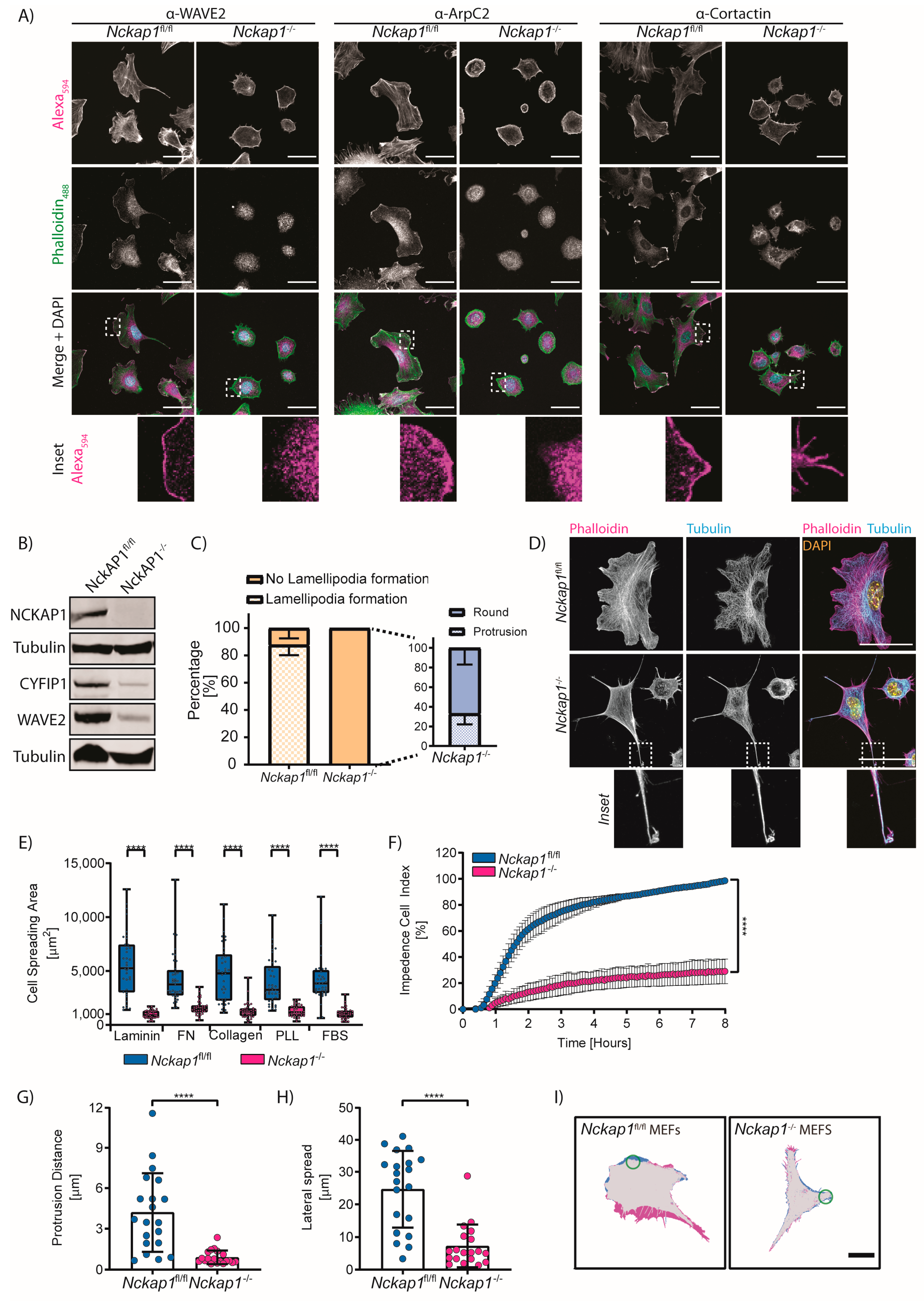
3.1. Generation of Inducible Nckap1 Knockout Mouse Embryonic Fibroblasts
3.2. NCKAP1 Is Required for Fibroblast Spreading
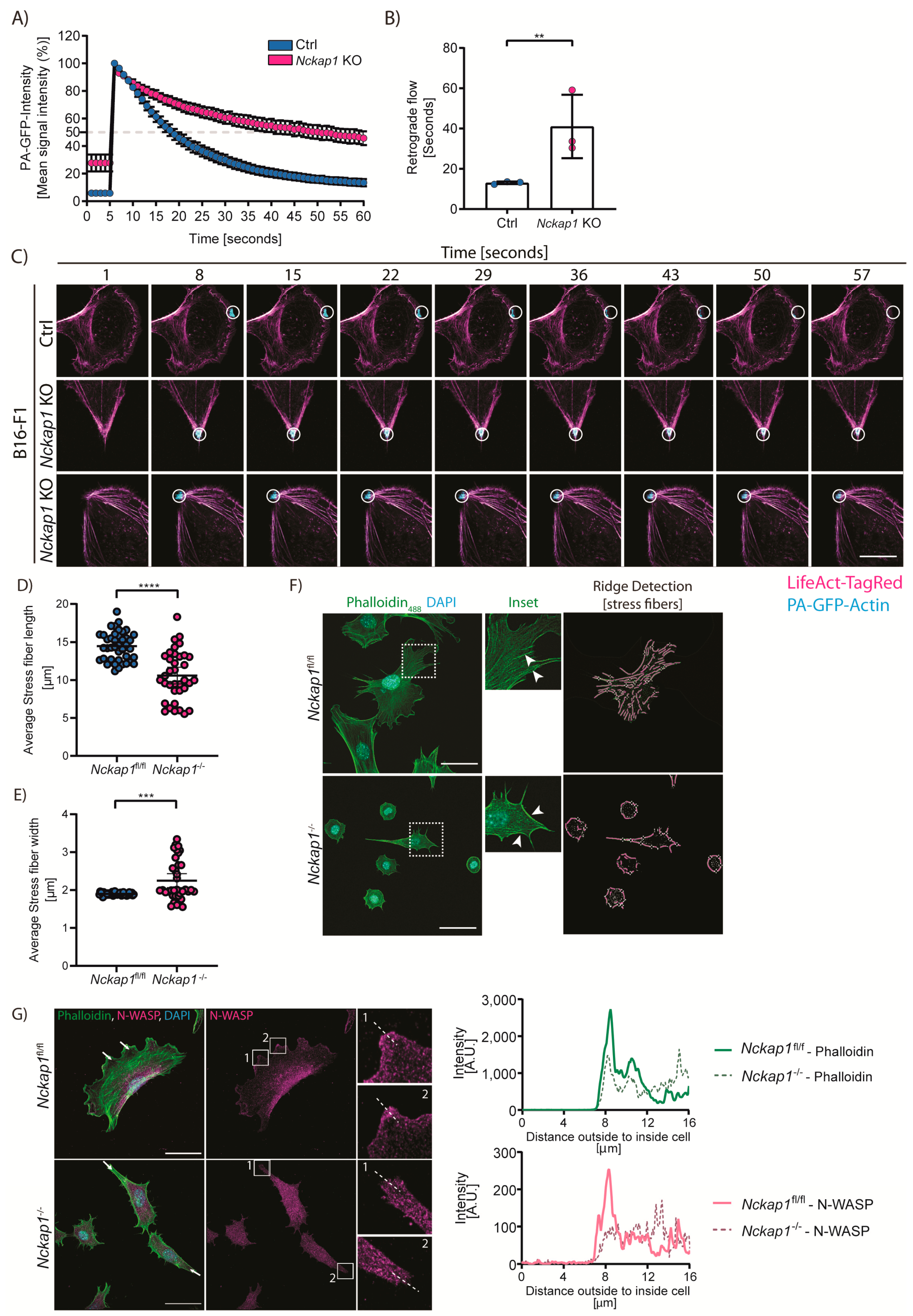
3.3. Loss of NCKAP1 Alters the Actin Dynamics in the Cell
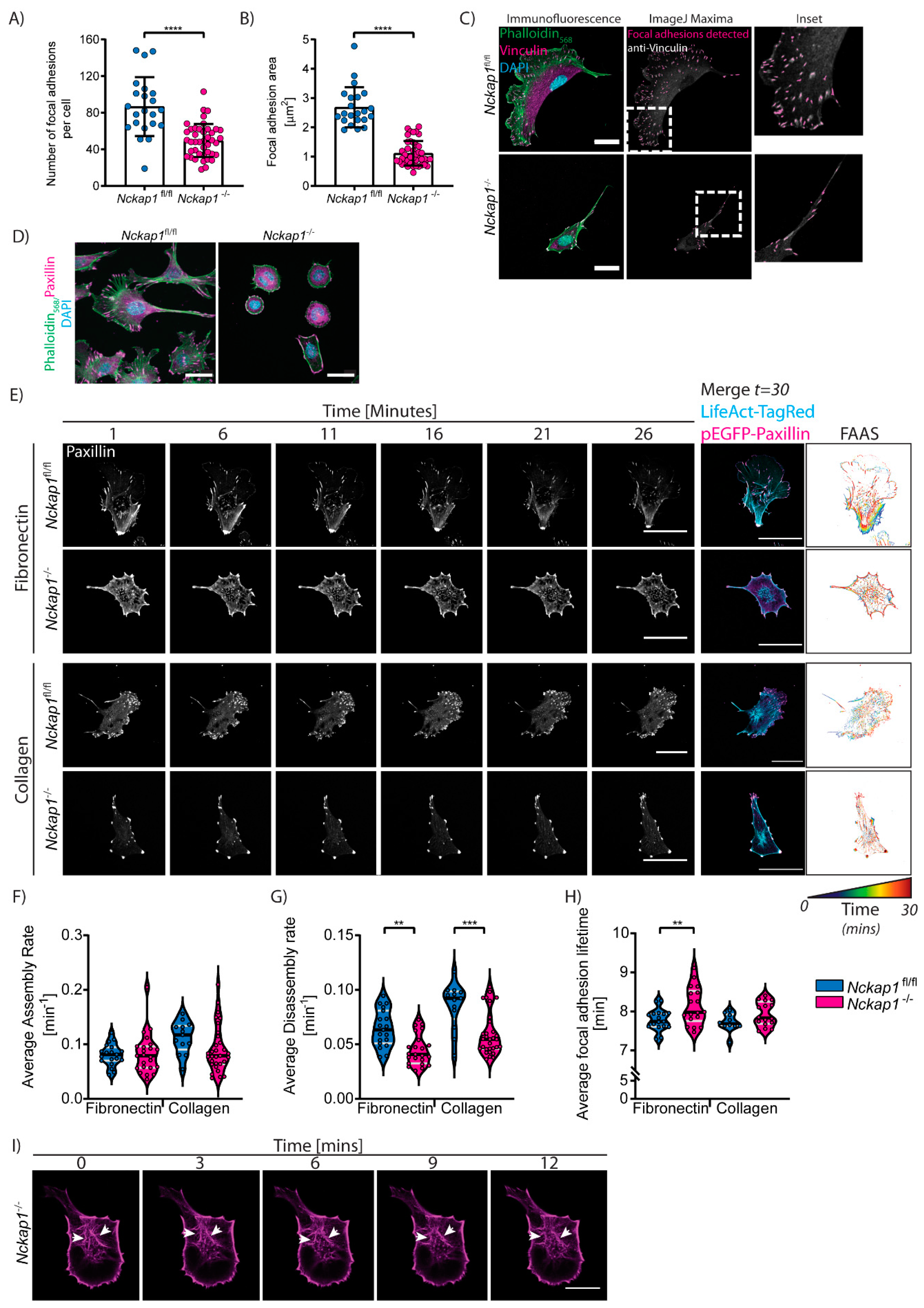
3.4. Loss of the Scar/WAVE Complex Affects Focal Adhesion Dynamics
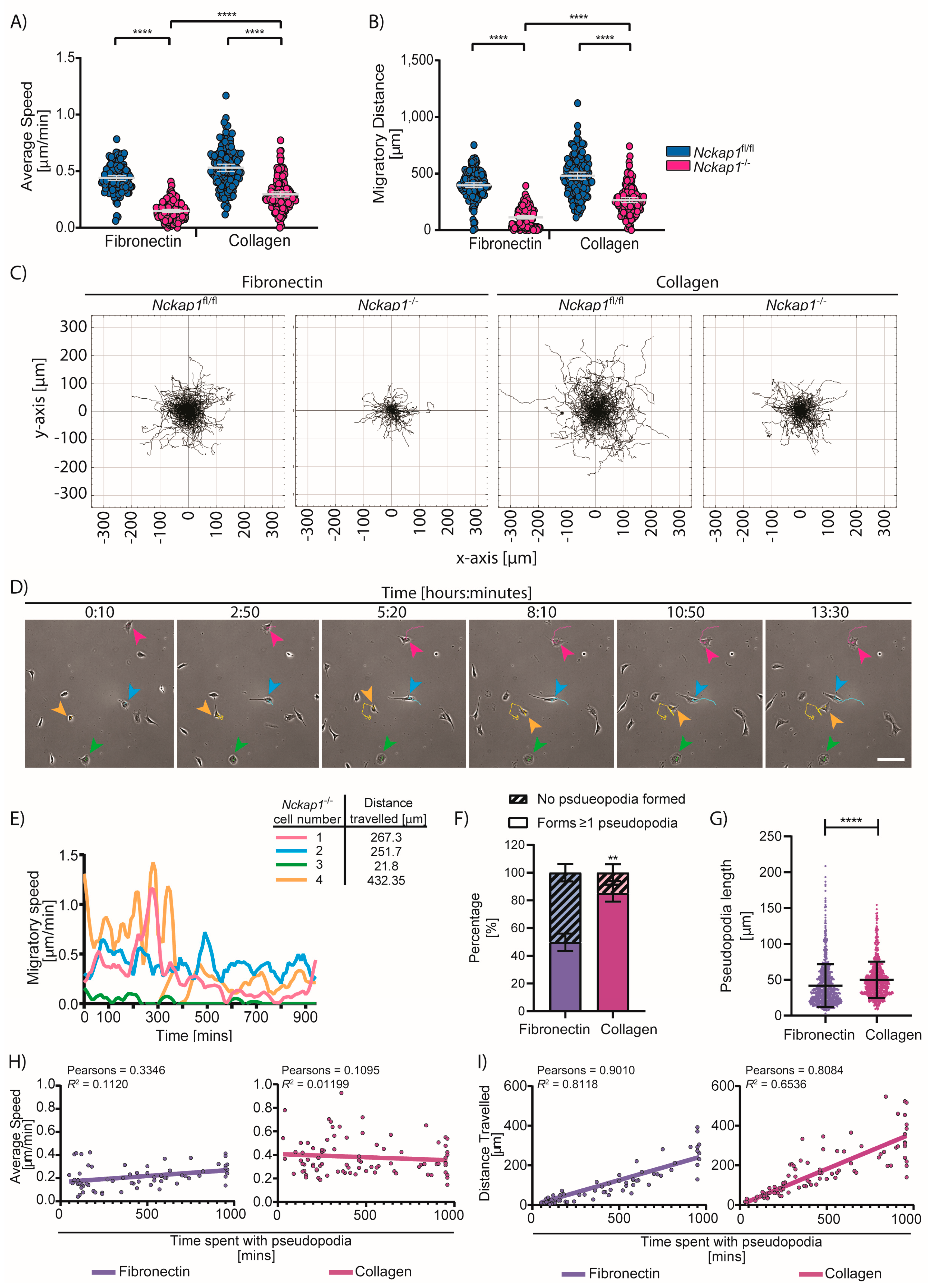
3.5. Nckap1 KO Cells Can Migrate without Lamellipodia on Collagen
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machesky, L.M.; Gould, K.L. The Arp2/3 complex: A multifunctional actin organizer. Curr. Opin. Cell Biol. 1999, 11, 117–121. [Google Scholar] [CrossRef]
- Hoffmann, B.D.; Grashoff, C.; Schwartz, M.A. Dynamic molecular processes mediate cellular mechanotransduction. Nature 2011, 475, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Machesky, L.M.; Insall, R.H. Scar1 and the related Wiskott–Aldrich syndrome protein, WASP, regulate the actin cytoskeleton through the Arp2/3 complex. Curr. Biol. 1998, 8, 1347–1356. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Borek, D.; Padrick, S.B.; Gomez, T.S.; Metlagel, Z.; Ismail, A.M.; Umetani, J.; Billadeau, D.D.; Otwinowski, Z.; Rosen, M.K. Structure and Control of the Actin Regulatory WAVE Complex. Nature 2010, 468, 533–538. [Google Scholar] [CrossRef]
- Chen, B.; Chou, H.-T.; Brautigam, C.A.; Xing, W.; Yang, S.; Henry, L.; Doolittle, L.K.; Walz, T.; Rosen, M.K. Rac1 GTPase Activates the WAVE Regulatory Complex Through Two Distinct Binding Sites. eLife 2017, 6, e29795. [Google Scholar] [CrossRef]
- Schaks, M.; Singh, S.P.; Kage, F.; Thomason, P.A.; Klünemann, T.; Steffen, A.; Blankenfeldt, W.; Stradal, T.E.; Insall, R.H.; Rottner, K. Distinct Interaction Sites of Rac GTPase with WAVE Regulatory Complex Have Non-redundant Functions in Vivo. Curr. Biol. 2018, 28, 3674–3684. [Google Scholar] [CrossRef] [Green Version]
- Dang, I.; Gorelik, R.; Sousa-Blin, C.; Derivery, E.; Guerin, C.; Linkner, J. Inhibitory Signalling to the Arp2/3 Complex Steers Cell Migration. Nature 2013, 503, 281–284. [Google Scholar] [CrossRef]
- Fort, L.; Batista, J.M.; Thomason, P.A.; Spence, H.J.; Whitelaw, J.A.; Tweedy, L.; Greaves, J.; Martin, K.J.; Anderson, K.I.; Brown, P.; et al. Fam49/CYRI interacts with Rac1 and locally supresses protrusions. Nat. Cell Biol. 2018, 20, 1159–1171. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Manser, E.; Loo, T.H.; Lim, L. Coupling of PAK-Interacting Exchange Factor PIX to GIT1 Promotes Focal Complex Disassembly. Mol. Cell. Biol. 2000, 20, 6354–6363. [Google Scholar] [CrossRef]
- Tang, H.; Li, A.; Bi, J.; Veltman, D.M.; Zech, T.; Spence, H.J.; Yu, X.; Timpson, P.; Insall, R.H.; Frame, M.; et al. Loss of Scar/WAVE Complex PromotesN-WASP- and FAK-Dependent Invasion. Curr. Biol. 2013, 23, 107–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.M.; Ezhkova, E.; Silva, J.; Heart, S.; Castillo, M.; Campos, Y.; Castro, V.; Bonilla, F.; Cordon-Cardo, C.; Muthuswamy, S.K.; et al. Cyfip1 Is a Putative Invasion Suppressor in Epithelial Cancers. Cell 2009, 137, 1047–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainero, E.; Howe, J.D.; Caswell, P.T.; Jamieson, N.B.; Anderson, K.I.; Critchley, D.R.; Machesky, L.M.; Norman, J.C. Ligand-Occupied Integrin Internalization Links Nutrient Signaling to Invasive Migration. Cell Rep. 2015, 10, 398–413. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, S.; Martin, D.; Chiquet-Ehrismann, R.; Sutton, J.; Desai, A.; Huang, I.; Kato, K.; Hromas, R. The HEM Proteins: A Novel Family of Tissue-specific Transmembrane Proteins Expressed from Invertebrates Through Mammals with an Essential Function in Oogenesis. J. Mol. Biol. 1995, 251, 41–49. [Google Scholar] [CrossRef]
- Hummel, T.; Leifker, K.; Klambt, C. The Drosophila HEM-2/NAP1 homolog KETTE controls axonal path finding and cytoskeletal organization. Genes Dev. 2000, 14, 863–873. [Google Scholar]
- Yokota, Y.; Ring, C.; Cheung, R.; Pevny, L.; Anton, E.S. Nap1 regulated neuronal cytoskeletal dynamics is essential forthe final differentiation of neurons in cerebral cortex. Neuron 2007, 54, 429–445. [Google Scholar] [CrossRef] [Green Version]
- Weiner, O.D.; Rentel, M.C.; Ott, A.; E Brown, G.; Jedrychowski, M.; Yaffe, M.B.; Gygi, S.P.; Cantley, L.C.; Bourne, H.R.; Kirschner, M.W. Hem-1 Complexes Are Essential for Rac Activation, Actin Polymerization, and Myosin Regulation during Neutrophil Chemotaxis. PLoS Biol. 2006, 4, e38. [Google Scholar] [CrossRef] [Green Version]
- Swaminathan, K. The RAC1 target NCKAP1 plays a crucial role in progression of BRAF/PTEN-driven melanoma in mice. J. Investig. Detmatol. 2020, in press. [Google Scholar]
- Magin, T.M.; McWhir, J.; Melton, D.W. A New Mouse Embryonic Stem cell Line With Good Germ Line Contribution and Gene Targeting Frequency. Nucleic Acids Res. 1992, 20, 3795–3796. [Google Scholar] [CrossRef] [Green Version]
- Nagy, A.; Behringer, R.; Gertsenstein, M.; Nagy, K.V. Manipulating the Mouse Embryo: A Laboratory Manual, 3rd ed.; Brownstein, D.G., Ed.; Cold Spring Harbour Laboratory Press: Suffolk County, NY, USA, 2003; Volume 78. [Google Scholar]
- Hameyer, D.; Loonstra, A.; Eshkind, L.; Schmitt, S.; Antunes, C.; Groen, A.; Bindels, E.; Jonkers, J.; Krimpenfort, P.; Meuwissen, R.; et al. Toxicity of ligand-dependent Cre recombinases and generation of a conditional Cre deleter mouse allowing mosaic recombination in peripheral tissues. Physiol. Genom. 2007, 31, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, J.; Frutschi, M.; Kaloulis, K.; McKee, T.; Trumpp, A.; Beermann, F. Metastasizing melanoma formation caused by expression of activated N-RasQ61K on an INK4a-dificent background. Cancer Res. 2005, 65, 4005–4011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durkin, M.E.; Qian, X.; Popetscu, N.; Lowy, D. Isolation of Mouse Embryo Fibroblasts. Bio. Protoc. 2013, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolati, S.; Kage, F.; Mueller, J.; Müsken, M.; Kirchner, M.; Dittmar, G.; Sixt, M.; Rottner, K.; Falcke, M. On the Relation Between Filament Density, Force Generation, and Protrusion Rate in Mesenchymal Cell Motility. Mol. Biol. Cell 2018, 29, 2674–2686. [Google Scholar] [CrossRef] [PubMed]
- Stahnke, S.; Döring, H.; Kusch, C.; De Gorter, D.J.; Dütting, S.; Guledani, A.; Pleines, I.; Schnoor, M.; Sixt, M.; Geffers, R.; et al. Loss of Hem1 disrupts macrophage function and impacts on migration, phagocytosis and integrin-mediated adhesion. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Berginski, M.E.; Gomez, S.M. The focal adhesions analysis server: A web tool for analyzing focal adhesion dynamics. F1000 Res. 2013, 2, 68. [Google Scholar] [CrossRef]
- Webb, D.J.; Donais, K.; Whitmore, L.A.; Thomas, S.M.; Turner, C.E.; Parsons, J.T.; Horwitz, A.F. FAK-Src signalling through paxillin, ERK and MLCK regulates adhesion disassembly. Nat. Cell Biol. 2004, 6, 154–161. [Google Scholar] [CrossRef]
- Berginski, M.E.; Vitriol, E.A.; Hahn, K.M.; Gomez, S.M. High-Resolution Quantificarion of Focal Adhesion Spatiotemporal Dynamics in Living Cells. PLoS ONE 2011, 6, e22025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papalazarou, V.; Zhang, T.; Paul, N.R.; Juin, A.; Cantini, M.; Maddocks, O.D.K.; Salmerón-Sánchez, M.; Machesky, L.M. The creatine–phosphagen system is mechanoresponsive in pancreatic adenocarcinoma and fuels invasion and metastasis. Nat. Metab. 2020, 2, 62–80. [Google Scholar] [CrossRef]
- Wu, Y.I.; Frey, D.; Lungu, O.I.; Jaehrig, A.; Schlichting, I.; Kuhlman, B.; Hahn, K.M. A Genetically Encoded Photoactivatable Rac Controls the Motility of Living Cells. Nature 2009, 461, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Steffen, A.; Rottner, K.; Ehinger, J.; Innocenti, M.; Scita, G.; Wehland, J.; Stradal, T.E. Sra-1 and Nap1 link Rac to actin assembly driving lamellipodia formation. EMBO J. 2004, 23, 749–759. [Google Scholar] [CrossRef] [Green Version]
- Krause, M.; Gautreau, A. Steering Cell Migration: Lamellipodium Dynamics and the Regulation of Directional Persistence. Nat. Rev. Mol. Cell Biol. 2014, 15, 577–590. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Ren, Y.; Wu, B.; Decourt, B.; Lee, A.C.; Taylor, A.; Suter, D.M. Src and Cortactin Promote Lamellipodia Protrsuion and Filopodia Formation and Stability in Growth Cones. Mol. Biol. Cell 2015, 26, 3229–3244. [Google Scholar] [CrossRef] [PubMed]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef] [Green Version]
- Salmela, M.; Jokinen, J.; Tiitta, S.; Rappu, P.; Cheng, R.H.; Heino, J. Integrin α2β1 in nonactivated conformation can induce focal adhesion kinase signaling. Sci. Rep. 2017, 7, 3414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kho, D.; Macdonald, C.; Johnson, R.; Unsworth, C.P.; O’Carroll, S.J.; Du Mez, E.; Angel, C.E.; Graham, E.S. Application of xCELLigence RTCA Biosensor Technology for Revealing the Profile and Window of Drug Responsiveness in Real Time. Biosensors 2015, 5, 199–222. [Google Scholar] [CrossRef] [Green Version]
- Schaks, M.; Giannone, G.; Rottner, K. Actin dynamics in cell migration. Essays Biochem. 2019, 63, 483–495. [Google Scholar]
- Small, J.V.; Isenberg, G.; Celis, J.E. Polarity of actin at the leading edge of cultured cells. Nature 1978, 272, 638–639. [Google Scholar] [CrossRef]
- Alexnadrova, A.Y.; Arnold, K.; Schaub, S.; Vasiliev, J.M.; Meister, J.-J.; Bershadsky, A.D.; Verkhovsky, A.B. Comparative Dynamics of Retrograde Actin Flow and Focal Adhesions: Formation of Nascent Adhesions Triggers Transition from Fast to Slow Flow. PLoS ONE 2008, 3, e3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashiro, S.; Watanabe, N. A new link between the retrograde actin flow and focal adhesions. J. Biochem. 2014, 156, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Lai, F.P.L.; Szczodrak, M.; Block, J.; Faix, J.; Breitsprecher, D.; Mannherz, H.G.; Stradal, T.E.B.; Dunn, G.A.; Small, J.V.; Rottner, K. Arp2/3 complex interactions and actin network turnover in lamellipodia. EMBO J. 2008, 27, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Amato, C.; Thomason, P.A.; Davidson, A.J.; Swaminathan, K.; Ismail, S.; Machesky, L.M.; Insall, R.H. WASP Restricts Active Rac to Maintain Cells’ Front-Rear Polarization. Curr. Biol. 2019, 29, 4169–4182. [Google Scholar] [CrossRef] [Green Version]
- Zaidel-Bar, R.; Ballestrem, C.; Kam, Z.; Geiger, B. Early molecular events in the assembly of matrixadhesions at the leading edge of migrating cells. J. Cell Sci. 2003, 116, 4605–4613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincente-Manzanares, M.; Choi, C.K.; Horwitz, A.R. Integrins in cell migration—The actin connection. J. Cell Sci. 2009, 122, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mali, K.A.; Barlow, C.A.; Burridge, K. Recruitment of the Arp2/3 complex to vinculin-coupling membrane protrusion to matrix adhesion. J. Cell Biol. 2002, 159, 881–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrels, B.; Serrels, A.; Brunton, V.G.; Holt, M.; McLean, G.W.; Gray, C.H.; Jones, G.E.; Frame, M.C. Focal adhesion kinase controls actin assembly via a FERM-mediated interaction with the Arp2/3 complex. Nat. Cell Biol. 2007, 9, 1046–1056. [Google Scholar] [CrossRef]
- Ryu, J.R.; Echarri, A.; Li, R.; Pendergast, A.M. Regulation of Cell-Cell Adhesion by Abi/Diaphanous Complexes. Mol. Cell. Biol. 2009, 29, 1735–1748. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, S.; Vaezi, A.; Fuchs, E. A role for αβ1 integrins in focal adhesion function and polarized cytoskeletal dynamics. Dev. Cell 2003, 5, 415–427. [Google Scholar] [CrossRef] [Green Version]
- Petrie, R.J.; Gavara, N.; Chadwick, R.S.; Yamada, K.M. Nonpolarized signaling reveals two distinct modes of 3D cell migration. J. Cell Biol. 2012, 197, 439–455. [Google Scholar] [CrossRef] [Green Version]
- Veltman, D.M.; King, J.S.; Machesky, L.M.; Insall, R.H. SCAR knockouts in Dictyostelium: WASP assumes SCAR’s position and upstream regulators in pseudopods. J. Cell Biol. 2012, 198, 501–508. [Google Scholar] [CrossRef] [Green Version]
- Davidson, A.J.; Amato, C.; Thomason, P.A.; Insall, R.H. WASP family proteins and formins compete in pseudopod- and bleb-based migration. J. Cell Biol. 2018, 217, 701–714. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, R.; Ma, L.; Miki, H.; Lopez, M.; Kirchhausen, T.; Takenawa, T.; Kirschner, M.W. The Interaction Between N-WASP and the Arp2/3 Complex Links Cdc42-dependent Signals to Actin Assembly. Cell 1999, 97, 221–231. [Google Scholar] [CrossRef] [Green Version]
- McCann, C.P.; Rericha, E.C.; Wang, C.; Losert, W.; Parent, C.A. Dictyostelium Cells Migrate Similarly on Surfaces of Varying Chemical Composition. PLoS ONE 2014, 9, e87981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardel, M.L.; Schneider, I.C.; Aratyn-Schaus, Y.; Waterman, C.M. Mechanical Integration of Actin and Adhesion Dynamics in Cell Migration. Annul. Rev. Cell. Dev. Biol. 2010, 26, 315–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraley, S.I.; Feng, Y.; Krishnamurthy, R.; Kim, D.-H.; Celedon, A.; Longmore, G.D.; Wirtz, D. A distinctive role for focal adhesion proteins in three-dimensional cell motility. Nat. Cell Biol. 2010, 12, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidel-Bar, R.; Milo, R.; Kam, Z.; Geiger, B. A paxillin tyrosine phosphorylation switch regulatesthe assembly and form of cell-matrix adhesions. J. Cell Sci. 2007, 120, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, N.; Kato, T.; Fujita, A.; Ishizaki, T.; Narumiya, S. Cooperation between mDia1 and ROCK in Rho-induced actin reorganization. Nat. Cell Biol. 1999, 1, 136–143. [Google Scholar] [CrossRef]
- Yamana, N.; Arakawa, Y.; Nishino, T.; Kurokawa, K.; Tanji, M.; Ito, R.E.; Monypenny, J.; Ishizaki, T.; Bito, H.; Nozaki, K.; et al. The Rho-mDia1 Pathway Regulates Cell Polarity and Focal AdhesionTurnover in Migrating Cells through Mobilizing Apc and c-Src. Mol. Cell. Biol. 2006, 26, 6844–6858. [Google Scholar] [CrossRef] [Green Version]
- Gupton, S.L.; Eisenmann, K.; Alberts, A.S.; Waterman-Storer, C.M. mDia2 regulates actin and focal adhesion dynamicsand organization in the lamella for efficient epithelialcell migration. J. Cell Sci. 2007, 120, 3475–3487. [Google Scholar] [CrossRef] [Green Version]
- Blagg, S.L.; Stewart, M.; Sambles, C.M.; Insall, R.H. PIR121 Regulates Pseudopod Dynamics and SCAR Activity in Dictyostelium. Curr. Biol. 2003, 13, 1480–1487. [Google Scholar] [CrossRef] [Green Version]
- Bouchet, B.P.; Akhmanova, A. Microtubules in 3D cell motility. J. Cell Sci. 2017, 130, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Petrie, R.J.; Yamada, K.M. At the leading edge of three: Dimensional cell migration. J. Cell Sci. 2012, 125, 5917–5926. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.M.; Sixt, M. Mechanisms of 3D cell migration. Nat. Rev. Cell Biol. 2019, 20, 738–752. [Google Scholar] [CrossRef]
- Zhu, J.; Mogilner, A. Comparison of cell migration mechanicalstrategies in three-dimensional matrices: A computational study. Interphase Focus 2016, 6, 20160040. [Google Scholar]
- Bouchet, B.P.; Noordstra, I.; Van Amersfoort, M.; Katrukha, E.A.; Ammon, Y.-C.; Ter Hoeve, N.D.; Hodgson, L.; Dogterom, M.; Derksen, P.W.; Akhmanova, A. Mesenchymal Cell Invasion Requires Cooperative Regulation of Persistent Microtubule Growth by SLAIN2 and CLASP1. Dev. Cell 2016, 39, 708–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayatilaka, H.; Giri, A.; Karl, M.; Aifuwa, I.; Trenton, N.J.; Phillip, J.M.; Khatau, S.; Wirtz, D. EB1 and cytoplasmic dynein mediate protrusion dynamics for efficient 3-dimensional cell migration. FASEB J. 2018, 32, 1207–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, A.D.; Wang, F.W.; Matsumoto, K.; Yamada, K.M. One-dimensional topography underlies three-dimensional fibrillar cell migration. J. Cell Biol. 2009, 184, 481–490. [Google Scholar] [CrossRef]
- Etienne-Manneville, S. Actin and microtubules in cell motility: Which one is in control? Traffic 2004, 5, 470–477. [Google Scholar] [CrossRef]
- Waterman-Storer, C.M.; Salmon, E.D. Actomyosin-based retrograde flow of microtubules in the lamella of migrating epithelial cells influences microtubule dynamic instability and turnover and is associated with mircotubule breakage and treadmilling. J. Cell Biol. 1997, 139, 417–434. [Google Scholar] [CrossRef]
- Rodriguez, O.; Schaefer, A.W.; Mandato, C.A.; Forscher, P.; Bement, W.M.; Waterman-Storer, C.M.; Waterman, C.M. Conserved microtubule-actin interactions in cell movements and morphogenesis. Nat. Cell Biol. 2003, 5, 599–609. [Google Scholar] [CrossRef]
- Etienne-Manneville, S.; Hall, A. Integrin-mediated Cdc42 activation controls cell polarity in migrating actrocytes through PKCζ. Cell 2001, 106, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Zinn, A.; Goicoechea, S.M.; Kreider-Letterman, G.; Maity, D.; Awadia, S.; Cedeno-Rosario, L.; Chen, Y.; Garcia-Mata, R. The small GTPase RhoG regulates microtubule-mediated focal adhesion disassembly. Sci. Rep. 2019, 9, 5163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartolini, F.; Moseley, J.B.; Schmoranzer, J.; Cassimeris, L.; Goode, B.L.; Gundersen, G.G. The formin mDia2 stabilizes microtubules independently of its actin nucleation activity. J. Cell Biol. 2008, 181, 523–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.; Yuan, F.N.; Corona, C.; Pasini, S.; Pero, M.E.; Gundersen, G.G.; Shelanski, M.L.; Bartolini, F. Stabilization of dynamic microtubules by mDia1 drives Tau-dependent Aβ1–42 synaptotoxicity. J. Cell Biol. 2017, 216, 3161–3178. [Google Scholar] [CrossRef] [Green Version]
- Courtemanche, N. Mechanisms of formin-mediated actin assembly and dynamics. Biophys. Rev. 2018, 10, 1553–1569. [Google Scholar] [CrossRef]
- Henty-Ridilla, J.L.; Rankova, A.; Eskin, J.A.; Kenny, K.; Goode, B.L. Accelerated actin filament polymerization from microtubule plus ends. Science 2016, 352, 1004–1009. [Google Scholar] [CrossRef] [Green Version]
- Garcin, C.; Straube, A. Microtubules in cell migration. Essays Biochem. 2019, 63, 509–520. [Google Scholar] [PubMed] [Green Version]
- Ezratty, E.J.; Partridge, M.A.; Gundersen, G.G. Microtubule-induced focal adhesion disassembly is mediated by dynamin and focal adhesion kinase. Nat. Cell Biol. 2005, 7, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Rooney, C.; White, G.; Nazgiewicz, A.; A Woodcock, S.; Anderson, K.I.; Ballestrem, C.; Malliri, A. The Rac activator STEF (Tiam2) regulates cell migration by microtubule-mediated focal adhesion disassembly. EMBO Rep. 2010, 11, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Stehbens, S.J.; Paszek, M.; Pemble, H.; Ettinger, A.; Gierke, S.; Wittmann, T. CLASPs link focal-adhesion-associated microtubulecapture to localized exocytosis and adhesionsite turnover. Nat. Cell Biol. 2014, 16, 561–573. [Google Scholar] [CrossRef]
- Gu, Z.; Noss, E.H.; Hsu, V.W.; Brenner, M.B. Integrins traffic rapidly via circular dorsal ruffles and macropinocytosis during stimulated cell migration. J. Cell Biol. 2011, 193, 61–70. [Google Scholar] [CrossRef]
- Eskova, A.; Knapp, B.; Matelska, D.; Reusing, S.; Arjonen, A.; Lisauskas, T.; Pepperkok, R.; Russell, R.B.; Eils, R.; Ivaska, J.; et al. An RNAi Screen Identifies KIF15 as a Novel Regulator of the Endocytic Trafficking of Integrin. J. Cell Sci. 2014, 127, 2433–2447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lansbergen, G.; Grigoriev, I.; Mimori-Kiyosue, Y.; Ohtsuka, T.; Higa, S.; Kitajima, I.; Demmers, J.; Galjart, N.; Houtsmuller, A.B.; Grosveld, F.; et al. CLASPs Attach Microtubule Plus Ends to the Cell Cortex Through a Complex With LL5β. Dev. Cell 2006, 11, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchet, B.P.; E Gough, R.; Ammon, Y.-C.; Van De Willige, D.; Post, H.; Jacquemet, G.; Altelaar, M.; Heck, A.J.; Goult, B.T.; Akhmanova, A. Talin-KANK1 interaction controls the recruitment of cortical microtubule stabilizing complexes to focal adhesions. eLife 2016, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, N.B.M.; Nishimura, Y.; Plotnikov, S.V.; Thiagarajan, V.; Zhang, Z.; Shi, S.; Natarajan, M.; Viasnoff, V.; Kanchanawong, P.; Jones, G.E.; et al. A Mechano-Signalling Network Linking Microtubules, Myosin IIA Filaments and Integrin-Based Adhesions. Nat. Mater. 2019, 18, 638–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whitelaw, J.A.; Swaminathan, K.; Kage, F.; Machesky, L.M. The WAVE Regulatory Complex Is Required to Balance Protrusion and Adhesion in Migration. Cells 2020, 9, 1635. https://doi.org/10.3390/cells9071635
Whitelaw JA, Swaminathan K, Kage F, Machesky LM. The WAVE Regulatory Complex Is Required to Balance Protrusion and Adhesion in Migration. Cells. 2020; 9(7):1635. https://doi.org/10.3390/cells9071635
Chicago/Turabian StyleWhitelaw, Jamie A., Karthic Swaminathan, Frieda Kage, and Laura M. Machesky. 2020. "The WAVE Regulatory Complex Is Required to Balance Protrusion and Adhesion in Migration" Cells 9, no. 7: 1635. https://doi.org/10.3390/cells9071635
APA StyleWhitelaw, J. A., Swaminathan, K., Kage, F., & Machesky, L. M. (2020). The WAVE Regulatory Complex Is Required to Balance Protrusion and Adhesion in Migration. Cells, 9(7), 1635. https://doi.org/10.3390/cells9071635