Wnt-5A/B Signaling in Hematopoiesis throughout Life

{kind=link}
{kind=link}
Abstract
:1. Introduction
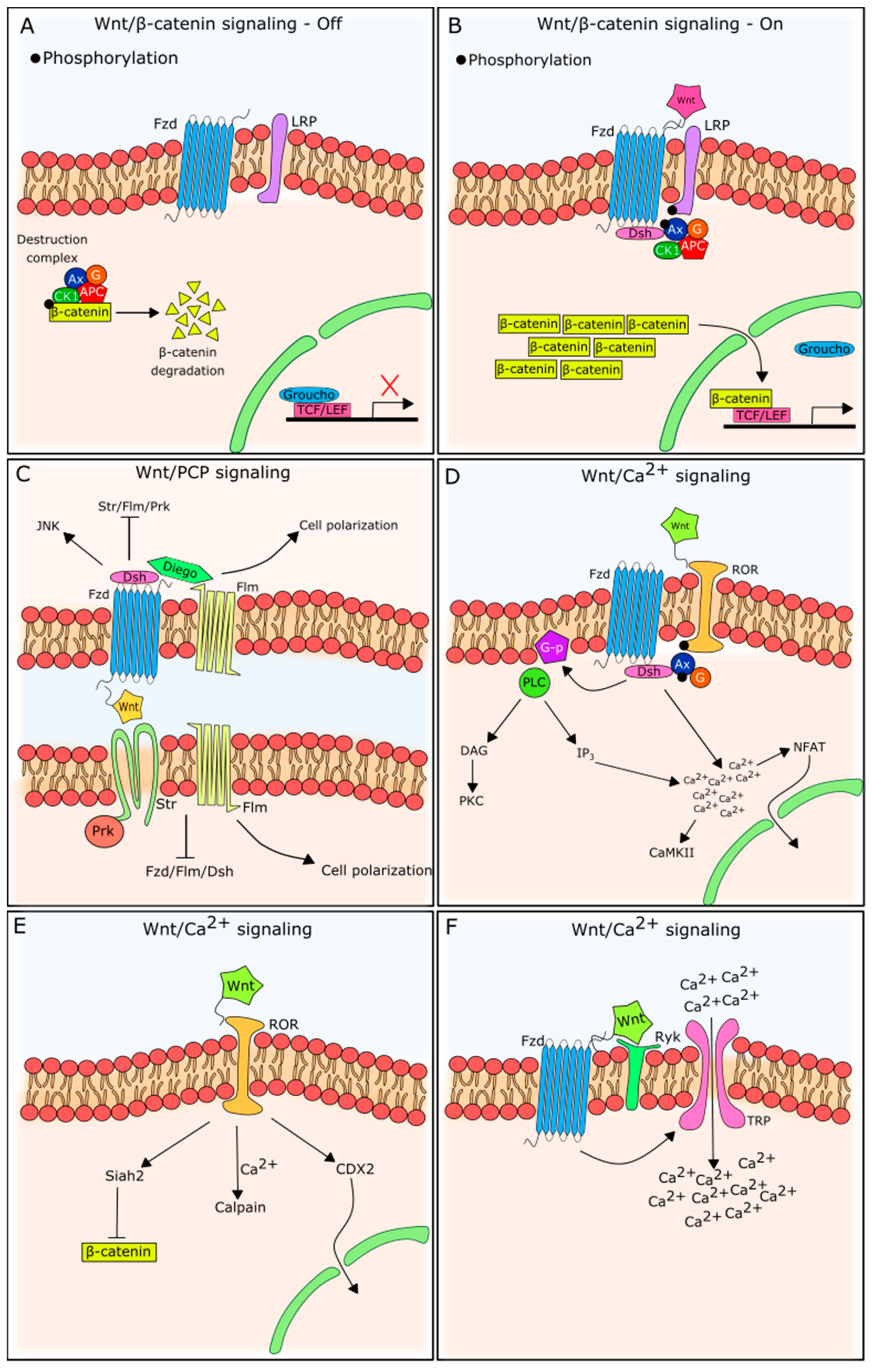
2. Wnt Signaling
2.1. β-Catenin-Dependent Wnt Signaling
2.2. β-Catenin-Independent Wnt Signaling
2.2.1. Wnt/PCP Signaling
2.2.2. Wnt/Ca2+ Signaling
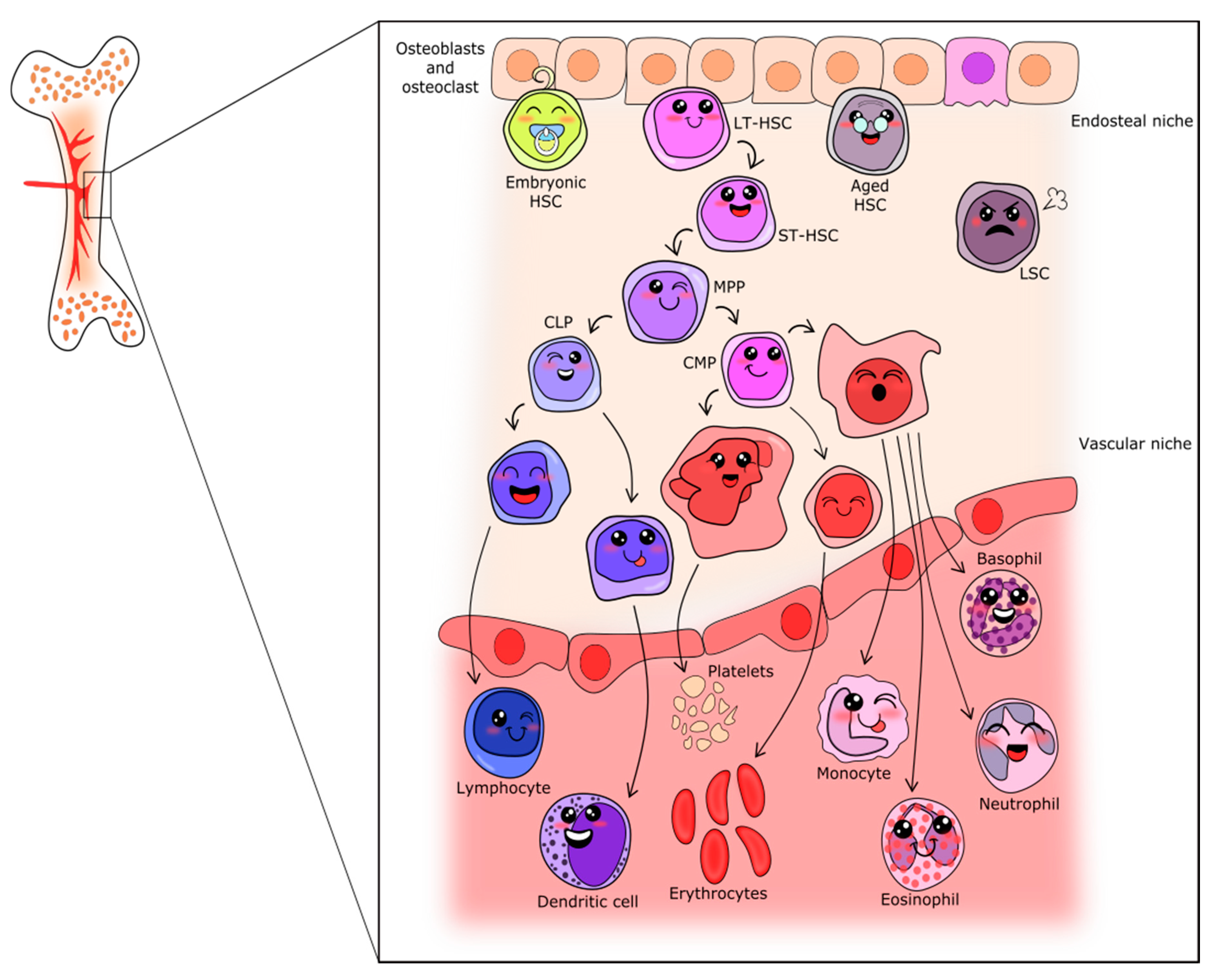
3. Wnt Signaling in Hematopoietic Ontogenesis
3.1. Hematopoietic Development
3.2. Definitive Hematopoiesis
4. Adult Hematopoiesis
5. Hematopoiesis during Aging
6. Hematopoietic Malignances
7. Targeting Wnt Signaling: Therapeutic Opportunities
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loughran, S.; Haas, S.; Wilkinson, A.C.; Klein, A.; Brand, M. Lineage commitment of hematopoietic stem cells and progenitors: Insights from recent single cell and lineage tracing technologies. Exp. Hematol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Carrelha, J.; Meng, Y.; Kettyle, L.M.; Luis, T.C.; Norfo, R.; Alcolea, V.; Boukarabila, H.; Grasso, F.; Gambardella, A.; Grover, A.; et al. Hierarchically related lineage-restricted fates of multipotent haematopoietic stem cells. Nature 2018, 554, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Cumano, A.; Godin, I. Ontogeny of the Hematopoietic System. Annu. Rev. Immunol. 2007, 25, 745–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, A.J.; McCulloch, E.A.; Till, J. Cytological Demonstration of the Clonal Nature of Spleen Colonies Derived from Transplanted Mouse Marrow Cells. Nature 1963, 197, 452–454. [Google Scholar] [CrossRef]
- Passegué, E.; Wagers, A.J.; Giuriato, S.; Anderson, W.C.; Weissman, I.L. Global analysis of proliferation and cell cycle gene expression in the regulation of hematopoietic stem and progenitor cell fates. J. Exp. Med. 2005, 202, 1599–1611. [Google Scholar] [CrossRef] [Green Version]
- Ting, S.B.; Deneault, E.; Hope, K.; Cellot, S.; Chagraoui, J.; Mayotte, N.; Dorn, J.F.; Laverdure, J.-P.; Harvey, M.; Hawkins, E.D.; et al. Asymmetric segregation and self-renewal of hematopoietic stem and progenitor cells with endocytic Ap2a. Blood 2012, 119, 2510–2522. [Google Scholar] [CrossRef]
- Marciniak-Czochra, A.; Stiehl, T.; Wagner, W. Modeling of replicative senescence in hematopoietic development. Aging 2009, 1, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, R.; Morita, Y.; Ooehara, J.; Hamanaka, S.; Onodera, M.; Rudolph, K.L.; Ema, H.; Nakauchi, H. Clonal Analysis Unveils Self-Renewing Lineage-Restricted Progenitors Generated Directly from Hematopoietic Stem Cells. Cell 2013, 154, 1112–1126. [Google Scholar] [CrossRef] [Green Version]
- Kondo, M.; Weissman, I.L.; Akashi, K. Identification of Clonogenic Common Lymphoid Progenitors in Mouse Bone Marrow. Cell 1997, 91, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Akashi, K.; Kondo, M.; Cheshier, S.; Shizuru, J.; Gandy, K.; Domen, J.; Mebius, R.; Traver, D.; Weissman, I. Lymphoid Development from Stem Cells and the Common Lymphocyte Progenitors. Cold Spring Harb. Symp. Quant. Biol. 1999, 64, 1–12. [Google Scholar] [CrossRef]
- Graf, T. Differentiation plasticity of hematopoietic cells. Blood 2002, 99, 3089–3101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, S.W.; Rajendiran, S.; Beaudin, A.E.; Smith-Berdan, S.; Muthuswamy, P.K.; Perez-Cunningham, J.; Martin, E.W.; Cheung, C.; Tsang, H.; Landon, M.; et al. Clonal and Quantitative In Vivo Assessment of Hematopoietic Stem Cell Differentiation Reveals Strong Erythroid Potential of Multipotent Cells. Stem Cell Rep. 2019, 12, 801–815. [Google Scholar] [CrossRef] [Green Version]
- Forsberg, E.C.; Serwold, T.; Kogan, S.; Weissman, I.L.; Passegue, E. New evidence supporting megakaryocyte-erythrocyte potential of flk2/flt3+ multipotent hematopoietic progenitors. Cell 2006, 126, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Miyawaki, K.; Arinobu, Y.; Iwasaki, H.; Kohno, K.; Tsuzuki, H.; Iino, T.; Shima, T.; Kikushige, Y.; Takenaka, K.; Miyamoto, T.; et al. CD41 Marks the Initial Myelo-Erythroid Lineage Specification in Adult Mouse Hematopoiesis: Redefinition of Murine Common Myeloid Progenitor. Stem Cells 2015, 33, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Drissen, R.; Thongjuea, S.; Theilgaard-Mönch, K.; Nerlov, C. Identification of two distinct pathways of human myelopoiesis. Sci. Immunol. 2019, 4, eaau7148. [Google Scholar] [CrossRef] [PubMed]
- Sanjuan-Pla, A.; Macaulay, I.C.; Jensen, C.T.; Woll, P.S.; Luis, T.C.; Mead, A.J.; Moore, S.; Carella, C.; Matsuoka, S.; Jones, T.B.; et al. Platelet-biased stem cells reside at the apex of the haematopoietic stem-cell hierarchy. Nature 2013, 502, 232–236. [Google Scholar] [CrossRef]
- Notta, F.; Zandi, S.; Takayama, N.; Dobson, S.; Gan, O.I.; Wilson, G.; Kaufmann, K.B.; McLeod, J.; Laurenti, E.; Dunant, C.F.; et al. Distinct routes of lineage development reshape the human blood hierarchy across ontogeny. Science 2015, 351, aab2116. [Google Scholar] [CrossRef] [Green Version]
- Paul, F.; Arkin, Y.; Giladi, A.; Jaitin, D.A.; Kenigsberg, E.; Keren-Shaul, H.; Winter, D.; Lara-Astiaso, D.; Gury, M.; Weiner, A.; et al. Transcriptional Heterogeneity and Lineage Commitment in Myeloid Progenitors. Cell 2015, 163, 1663–1677. [Google Scholar] [CrossRef] [Green Version]
- Nestorowa, S.; Hamey, F.K.; Pijuan-Sala, B.; Diamanti, E.; Shepherd, M.; Laurenti, E.; Wilson, N.K.; Kent, D.G.; Göttgens, B. A single-cell resolution map of mouse hematopoietic stem and progenitor cell differentiation. Blood 2016, 128, e20–e31. [Google Scholar] [CrossRef] [Green Version]
- Doulatov, S.; Notta, F.; Eppert, K.; Nguyen, L.T.; Ohashi, P.S.; Dick, J.E. Revised map of the human progenitor hierarchy shows the origin of macrophages and dendritic cells in early lymphoid development. Nat. Immunol. 2010, 11, 585–593. [Google Scholar] [CrossRef]
- Nusse, R. Wnts and Hedgehogs: Lipid-modified proteins and similarities in signaling mechanisms at the cell surface. Development 2003, 130, 5297–5305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willert, K.; Brown, J.D.; Danenberg, E.; Duncan, A.W.; Weissman, I.L.; Reya, T.; Yates, J.R.; Nusse, R. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature 2003, 423, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.-C.; Rattner, A.; Smallwood, P.M.; Nathans, J. Biochemical characterization of Wnt-Frizzled interactions using a soluble, biologically active vertebrate Wnt protein. Proc. Natl. Acad. Sci. USA 1999, 96, 3546–3551. [Google Scholar] [CrossRef] [Green Version]
- Janda, C.Y.; Waghray, D.; Levin, A.M.; Thomas, C.; Garcia, K.C. Structural Basis of Wnt Recognition by Frizzled. Science 2012, 337, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, B.T.; Hien, A.; Zhang, X.; Iranloye, O.; Virshup, D.M.; Waterman, M.L.; He, X. Disulfide Bond Requirements for Active Wnt Ligands. J. Biol. Chem. 2014, 289, 18122–18136. [Google Scholar] [CrossRef] [Green Version]
- Tamai, K.; Semenov, M.; Kato, Y.; Spokony, R.; Liu, C.; Katsuyama, Y.; Hess, F.; Saint-Jeannet, J.-P.; He, X. LDL-receptor-related proteins in Wnt signal transduction. Nature 2000, 407, 530–535. [Google Scholar] [CrossRef]
- Pinson, K.I.; Brennan, J.; Monkley, S.; Avery, B.J.; Skarnes, W.C. An LDL-receptor-related protein mediates Wnt signalling in mice. Nature 2000, 407, 535–538. [Google Scholar] [CrossRef]
- Skronska-Wasek, W.; Gosens, R.; Königshoff, M.; Baarsma, H.A. WNT receptor signalling in lung physiology and pathology. Pharmacol. Ther. 2018, 187, 150–166. [Google Scholar] [CrossRef]
- Kawano, Y.; Kypta, R. Secreted antagonists of the Wnt signalling pathway. J. Cell Sci. 2003, 116, 2627–2634. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Semenov, M.; Tomai, K.; Zeng, X. LDL receptor-related proteins 5 and 6 in Wnt/beta-catenin signaling: Arrows point the way. Development 2004, 131, 1663–1677. [Google Scholar] [CrossRef] [Green Version]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamai, K.; Zeng, X.; Liu, C.; Zhang, X.; Harada, Y.; Chang, Z.; He, X. A Mechanism for Wnt Coreceptor Activation. Mol. Cell 2004, 13, 149–156. [Google Scholar] [CrossRef]
- Clevers, H.; Loh, K.M.; Nusse, R. Faculty Opinions recommendation of Stem cell signaling. An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Science 2014, 346, 1248012. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; A Julius, M.; Giarré, M.; Zheng, Z.; Brown, A.M.; Kitajewski, J. Transformation by Wnt family proteins correlates with regulation of beta-catenin. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 1997, 8, 1349–1358. [Google Scholar]
- Famili, F.; Naber, B.A.E.; Vloemans, S.; De Haas, E.; Tiemessen, E.F.; Staal, F.J.T. Discrete roles of canonical and non-canonical Wnt signaling in hematopoiesis and lymphopoiesis. Cell Death Dis. 2015, 6, e1981. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Orte, E.; Sáenz-Narciso, B.; Moreno, S.; Cabello, J. Multiple functions of the noncanonical Wnt pathway. Trends Genet. 2013, 29, 545–553. [Google Scholar] [CrossRef]
- Semenov, M.V.; Habas, R.; Macdonald, B.T.; He, X. SnapShot: Noncanonical Wnt Signaling Pathways. Cell 2007, 131, 1378.e1–1378.e2. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Mlodzik, M. Wnt-Frizzled/planar cell polarity signaling: Cellular orientation by facing the wind (Wnt). Annu. Rev. Cell Dev. Biol. 2015, 31, 623–646. [Google Scholar] [CrossRef] [Green Version]
- Vinson, C.R.; Adler, P.N. Directional non-cell autonomy and the transmission of polarity information by the frizzled gene of Drosophila. Nature 1987, 329, 549–551. [Google Scholar] [CrossRef]
- Gubb, D.; García-Bellido, A. A genetic analysis of the determination of cuticular polarity during development in Drosophila melanogaster. J. Embryol. Exp. Morphol. 1982, 68, 37–57. [Google Scholar]
- Seifert, J.R.K.; Mlodzik, M. Frizzled/PCP signalling: A conserved mechanism regulating cell polarity and directed motility. Nat. Rev. Genet. 2007, 8, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Chen, Y.-G. Dishevelled: The hub of Wnt signaling. Cell. Signal. 2010, 22, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Nusse, R.; Van Amerongen, R. The Role of Ryk and Ror Receptor Tyrosine Kinases in Wnt Signal Transduction. Cold Spring Harb. Perspect. Biol. 2014, 6, a009175. [Google Scholar] [CrossRef]
- Pataki, C.A.; Couchman, J.R.; Brábek, J. Wnt Signaling Cascades and the Roles of Syndecan Proteoglycans. J. Histochem. Cytochem. 2015, 63, 465–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodrich, L.V.; Strutt, D.I. Principles of planar polarity in animal development. Development 2011, 138, 1877–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.; Chen, S.; Cai, S.; Dong, L.; Liu, L.; Yang, Y.; Guo, F.; Lu, X.; He, H.; Chen, Q.; et al. Wnt5a through Noncanonical Wnt/JNK or Wnt/PKC Signaling Contributes to the Differentiation of Mesenchymal Stem Cells into Type II Alveolar Epithelial Cells In Vitro. PLoS ONE 2014, 9, e90229. [Google Scholar] [CrossRef] [PubMed]
- Saadeddin, A.; Babaei-Jadidi, R.; Spencer-Dene, B.; Nateri, A.S. The Links between Transcription, -catenin/JNK Signaling, and Carcinogenesis. Mol. Cancer Res. 2009, 7, 1189–1196. [Google Scholar] [CrossRef] [Green Version]
- Kühl, M.; Sheldahl, L.C.; Park, M.; Miller, J.R.; Moon, R.T. The Wnt/Ca2+ pathway: A new vertebrate Wnt signaling pathway takes shape. Trends Genet. 2000, 16, 279–283. [Google Scholar] [CrossRef]
- Kühl, M. The WNT/calcium pathway: Biochemical mediators, tools and future requirements. Front Biosci. 2004, 9, 967–974. [Google Scholar] [CrossRef] [Green Version]
- Martineau, X.; Abed, É.; Martel-Pelletier, J.; Pelletier, J.-P.; Lajeunesse, D. Alteration of Wnt5a expression and of the non-canonical Wnt/PCP and Wnt/PKC-Ca2+ pathways in human osteoarthritis osteoblasts. PLoS ONE 2017, 12, e0180711. [Google Scholar] [CrossRef] [Green Version]
- De, A. Wnt/Ca2+ signaling pathway: A brief overview. Acta Biochim. Biophys. Sin. 2011, 43, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.E.J.; Liu, Y.; Cooper, H.M. The Yin and Yang of Wnt/Ryk axon guidance in development and regeneration. Sci. China Life Sci. 2014, 57, 366–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julien, E.; El Omar, R.; Tavian, M. Origin of the hematopoietic system in the human embryo. FEBS Lett. 2016, 590, 3987–4001. [Google Scholar] [CrossRef] [Green Version]
- Waas, B.; Maillard, I. Fetal hematopoietic stem cells are making waves. Stem Cell Investig. 2017, 4, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benz, C.; Copley, M.R.; Kent, D.G.; Wohrer, S.; Cortes, A.; Aghaeepour, N.; Ma, E.; Mader, H.; Rowe, K.; Day, C.; et al. Hematopoietic Stem Cell Subtypes Expand Differentially during Development and Display Distinct Lymphopoietic Programs. Cell Stem Cell 2012, 10, 273–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaudin, A.E.; Boyer, S.W.; Perez-Cunningham, J.; Hernandez, G.E.; Derderian, S.C.; Jujjavarapu, C.; Aaserude, E.; MacKenzie, T.; Forsberg, E.C. A Transient Developmental Hematopoietic Stem Cell Gives Rise to Innate-like B and T Cells. Cell Stem Cell 2016, 19, 768–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Perdiguero, E.; Klapproth, K.; Schulz, C.; Busch, K.; Azzoni, E.; Crozet, L.; Garner, H.; Trouillet, C.; De Bruijn, M.; Geissmann, F. Tissue-resident macrophages originate from yolk sac-derived erythro-myeloid progenitors. Nature 2015, 518, 547–551. [Google Scholar] [CrossRef]
- Hoeffel, G.; Chen, J.; Lavin, Y.; Low, D.; Almeida, F.F.; See, P.; Beaudin, A.E.; Lum, J.; Low, I.; Forsberg, E.C.; et al. C-Myb+ Erythro-Myeloid Progenitor-Derived Fetal Monocytes Give Rise to Adult Tissue-Resident Macrophages. Immunity 2015, 42, 665–678. [Google Scholar] [CrossRef] [Green Version]
- Corrigan, P.M.; Dobbin, E.; Freeburn, R.W.; Wheadon, H. Patterns of Wnt/Fzd/LRP Gene Expression During Embryonic Hematopoiesis. Stem Cells Dev. 2009, 18, 759–772. [Google Scholar] [CrossRef]
- Lako, M.; Lindsay, S.; Lincoln, J.; Cairns, P.M.; Armstrong, L.; Hole, N. Characterisation of Wnt gene expression during the differentiation of murine embryonic stem cells in vitro: Role of Wnt3 in enhancing haematopoietic differentiation. Mech. Dev. 2001, 103, 49–59. [Google Scholar] [CrossRef]
- Tai, C.C.; Sala, F.G.; Ford, H.R.; Wang, K.S.; Li, C.; Minoo, P.; Grikscheit, T.C.; Bellusci, S. Wnt5a Knock-out Mouse as a New Model of Anorectal Malformation. J. Surg. Res. 2009, 156, 278–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements, W.K.; Kim, A.D.; Ong, K.G.; Morre, J.C.; Lawson, N.D.; Traver, D. A somitic Wnt16/Notch pathway specifies haematopoietic stem cells. Nature 2011, 474, 220–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busch, K.; Klapproth, K.; Barile, M.; Flossdorf, M.; Holland-Letz, T.; Schlenner, S.M.; Reth, M.; Höfer, T.; Rodewald, H.-R. Fundamental properties of unperturbed haematopoiesis from stem cells in vivo. Nature 2015, 518, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Ciriza, J.; Thompson, H.; Petrosian, R.; Manilay, J.O.; García-Ojeda, M.E. The migration of hematopoietic progenitors from the fetal liver to the fetal bone marrow: Lessons learned and possible clinical applications. Exp. Hematol. 2013, 41, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wan, L.; Wang, C.; Zhou, K. Integration of Shh and Wnt Signaling Pathways Regulating Hematopoiesis. DNA Cell Biol. 2015, 34, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, D.; Kokkaliaris, K.; Schroeder, T. Wnt to Notch Relay Signaling Induces Definitive Hematopoiesis. Cell Stem Cell 2011, 9, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.B.; Chabner, K.T.; Alley, I.R.; Olson, D.P.; Szczepiorkowski, Z.M.; Poznansky, M.C.; Kos, C.H.; Pollak, M.R.; Brown, E.M.; Scadden, D.T. Stem cell engraftment at the endosteal niche is specified by the calcium-sensing receptor. Nature 2006, 439, 599–603. [Google Scholar] [CrossRef]
- Adams, G.B.; Scadden, D.T. The hematopoietic stem cell in its place. Nat. Immunol. 2006, 7, 333–337. [Google Scholar] [CrossRef]
- Birbrair, A.; Frenette, P.S. Niche heterogeneity in the bone marrow. Ann. New York Acad. Sci. 2016, 1370, 82–96. [Google Scholar] [CrossRef]
- Cheshier, S.H.; Morrison, S.J.; Liao, X.; Weissman, I.L. In vivo proliferation and cell cycle kinetics of long-term self-renewing hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 1999, 96, 3120–3125. [Google Scholar] [CrossRef] [Green Version]
- Seita, J.; Weissman, I.L. Hematopoietic stem cell: Self-renewal versus differentiation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 640–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, T.W.; Solar, G.P.; Ziegler, F.C.; Liem, L.; Matthews, W. A role for the Wnt gene family in hematopoiesis: Expansion of multilineage progenitor cells. Blood 1997, 89, 3624–3635. [Google Scholar] [CrossRef]
- Berg, D.J.V.D.; Sharma, A.K.; Bruno, E.; Hoffman, R. Role of members of the Wnt gene family in human hematopoiesis. Blood 1998, 92, 3189–3202. [Google Scholar] [CrossRef]
- Reya, T.; O’Riordan, M.X.D.; Okamura, R.; Devaney, E.; Willert, K.; Nusse, R.; Grosschedl, R. Wnt Signaling Regulates B Lymphocyte Proliferation through a LEF-1 Dependent Mechanism. Immunology 2000, 13, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, M.J.; Topol, L.; Anderson, S.M.; Yang, Y.; Bodine, D.M. Wnt5a inhibits canonical Wnt signaling in hematopoietic stem cells and enhances repopulation. Proc. Natl. Acad. Sci. USA 2007, 104, 15436–15441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolova, T.; Wu, M.; Brumbarov, K.; Alt, R.; Opitz, H.; Boheler, K.R.; Cross, M.; Wobus, A.M. WNT-conditioned media differentially affect the proliferation and differentiation of cord blood-derived CD133+ cells in vitro. Differentiation 2007, 75, 100–111. [Google Scholar] [CrossRef]
- Malhotra, S.; Baba, Y.; Garrett, K.P.; Staal, F.J.T.; Gerstein, R.; Kincade, P.W. Contrasting Responses of Lymphoid Progenitors to Canonical and Non-canonical Wnt Signals. J. Immunol. 2008, 181, 3955–3964. [Google Scholar] [CrossRef]
- Jafarzadeh, N.; Safari, Z.; Pornour, M.; Amirizadeh, N.; Moghadam, M.F.; Sadeghizadeh, M. Alteration of cellular and immune-related properties of bone marrow mesenchymal stem cells and macrophages by K562 chronic myeloid leukemia cell derived exosomes. J. Cell. Physiol. 2019, 234, 3697–3710. [Google Scholar] [CrossRef]
- Xu, J.; Li, X.; Cole, A.; Sherman, Z.; Du, W. Reduced Cell Division Control Protein 42 Activity Compromises Hematopoiesis-Supportive Function of Fanconi Anemia Mesenchymal Stromal Cells. Stem Cells 2018, 36, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Povinelli, B.J.; Nemeth, M.J. Wnt5a Regulates Hematopoietic Stem Cell Proliferation and Repopulation Through the Ryk Receptor. Stem Cells 2014, 32, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Florian, M.C.; Nattamai, K.J.; Dörr, K.; Marka, G.; Überle, B.; Vas, V.; Eckl, C.; Andrä, I.; Schiemann, M.; Oostendorp, R.; et al. A canonical to non-canonical Wnt signalling switch in haematopoietic stem-cell ageing. Nature 2013, 503, 392–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.M.; Mishina, Y.M.; Cong, F. Tankyrase inhibition stabilizes axin and antagonizes Wnt signalling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Schaap-Oziemlak, A.M.; Schouteden, S.; Khurana, S.; Verfaillie, C. Wnt5a Does Not Support Hematopoiesis in Stroma-Free, Serum-Free Cultures. PLoS ONE 2013, 8, e53669. [Google Scholar] [CrossRef] [PubMed]
- Buckley, S.M.; Ulloa-Montoya, F.; Abts, D.; Oostendorp, R.; Dzierzak, E.; Ekker, S.C.; Verfaillie, C.M. Maintenance of HSC by Wnt5a secreting AGM-derived stromal cell line. Exp. Hematol. 2011, 39, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimura, R.; He, X.C.; Venkatraman, A.; Arai, F.; Box, A.; Semerad, C.; Haug, J.S.; Peng, L.; Zhong, X.-B.; Suda, T.; et al. Noncanonical Wnt signaling maintains hematopoietic stem cells in the niche. Cell 2012, 150, 351–365. [Google Scholar] [CrossRef] [Green Version]
- Despeaux, M.; Chicanne, G.; Rouer, E.; De Toni-Costes, F.; Bertrand, J.; Mas, V.M.-D.; Vergnolle, N.; Eaves, C.; Payrastre, B.; Girault, J.-A.; et al. Focal Adhesion Kinase Splice Variants Maintain Primitive Acute Myeloid Leukemia Cells Through Altered Wnt Signaling. Stem Cells 2012, 30, 1597–1610. [Google Scholar] [CrossRef]
- Vannini, N.; Roch, A.; Naveiras, O.; Griffa, A.; Köbel, S.; Lutolf, M. Identification of in vitro HSC fate regulators by differential lipid raft clustering. Cell Cycle 2012, 11, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, B.; Chadwick, K.; Martin, M.; Shojaei, F.; Shah, K.V.; Gallacher, L.; Moon, R.T.; Bhatia, M. Wnt-5A augments repopulating capacity and primitive hematopoietic development of human blood stem cells in vivo. Proc. Natl. Acad. Sci. USA 2003, 100, 3422–3427. [Google Scholar] [CrossRef] [Green Version]
- Trowbridge, J.J.; Guezguez, B.; Moon, R.T.; Bhatia, M. Wnt3a Activates Dormant c-Kit− Bone Marrow-Derived Cells with Short-Term Multilineage Hematopoietic Reconstitution Capacity. Stem Cells 2010, 28, 1379–1389. [Google Scholar] [CrossRef] [Green Version]
- Busslinger, M. Transcriptional Control of Early B Cell Development. Annu. Rev. Immunol. 2004, 22, 55–79. [Google Scholar] [CrossRef]
- Xiao, J.; Zhou, H.; Wu, N.; Wu, L. The non-canonical Wnt pathway negatively regulates dendritic cell differentiation by inhibiting the expansion of Flt3+ lymphocyte-primed multipotent precursors. Cell. Mol. Immunol. 2016, 13, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Istvanffy, R.; Schreck, C.; Ziegenhein, C.; Sippenauer, T.; Romero, S.; Hettler, F.; Florian, C.; Waskow, C.; Essers, M.; Peschel, C.; et al. Niche Wnt5a regulates the actin cytoskeleton during regeneration of hematopoietic stem cells. Exp. Hematol. 2017, 53, S96. [Google Scholar] [CrossRef] [Green Version]
- De Rezende, M.M.; Ng-Blichfeldt, J.-P.; Justo, G.Z.; Paredes-Gamero, E.J.; Gosens, R. Divergent effects of Wnt5b on IL-3- and GM-CSF-induced myeloid differentiation. Cell. Signal. 2020, 67, 109507. [Google Scholar] [CrossRef] [PubMed]
- Nteliopoulos, G.; Marley, S.B.; Gordon, M.Y. Influence of PI-3K/Akt pathway on Wnt signalling in regulating myeloid progenitor cell proliferation. Evidence for a role of autocrine/paracrine Wnt regulation. Br. J. Haematol. 2009, 146, 637–651. [Google Scholar] [CrossRef]
- Brandon, C.; Eisenberg, L.M.; Eisenberg, C.A. WNT signaling modulates the diversification of hematopoietic cells. Blood 2000, 96, 4132–4141. [Google Scholar] [CrossRef]
- Louwette, S.; Labarque, V.; Wittevrongel, C.; Thys, C.; Metz, J.R.; Gijsbers, R.; Debyser, Z.; Arnout, J.; Van Geet, C.; Freson, K. Regulator of G-protein signaling 18 controls megakaryopoiesis and the cilia-mediated vertebrate mechanosensory system. FASEB J. 2012, 26, 2125–2136. [Google Scholar] [CrossRef]
- Skokowa, J.; Cario, G.; Uenalan, M.; Schambach, A.; Germeshausen, M.; Battmer, K.; Zeidler, C.; Lehmann, U.; Eder, M.; Baum, C.; et al. LEF-1 is crucial for neutrophil granulocytopoiesis and its expression is severely reduced in congenital neutropenia. Nat. Med. 2006, 12, 1191–1197. [Google Scholar] [CrossRef]
- Wang, Y.; Sano, S.; Oshima, K.; Sano, M.; Watanabe, Y.; Katanasaka, Y.; Yura, Y.; Jung, C.; Anzai, A.; Swirski, F.K.; et al. Wnt5a-Mediated Neutrophil Recruitment Has an Obligatory Role in Pressure Overload-Induced Cardiac Dysfunction. Circulation 2019, 140, 487–499. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Q.; Yang, Z.; Shao, Y.; Xue, P.; Qu, W.; Jia, X.; Cheng, L.; He, M.; He, R.; et al. Cadmium Activates Noncanonical Wnt Signaling to Impair Hematopoietic Stem Cell Function in Mice. Toxicol. Sci. 2018, 165, 254–266. [Google Scholar] [CrossRef]
- Ashihara, E.; Takada, T.; Maekawa, T. Targeting the canonical Wnt/β-catenin pathway in hematological malignancies. Cancer Sci. 2015, 106, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.; Ju, W.; Huang, Y.; Li, J.; Ozer, H.; Qiao, X.; Qian, Z. Activation of Wnt/β-catenin Signaling Blocks Monocyte-Macrophage Differentiation Through Antagonizing PU.1-Targeted Gene Transcription. Leukemia 2016, 30, 2106–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, E.J.; Hwang, S.-G.; Nguyen, P.; Lee, S.; Kim, J.-S.; Kim, J.W.; Henkart, P.A.; Bottaro, D.; Soon, L.; Bonvini, P.; et al. Regulation of leukemic cell adhesion, proliferation, and survival by β-catenin. Blood 2002, 100, 982–990. [Google Scholar] [CrossRef]
- Ysebaert, L.; Chicanne, G.; Demur, C.; De Toni, F.; Prade-Houdellier, N.; Ruidavets, J.-B.; Mas, V.M.-D.; Rigal-Huguet, F.; Laurent, G.; Payrastre, B.; et al. Expression of β-catenin by acute myeloid leukemia cells predicts enhanced clonogenic capacities and poor prognosis. Leukemia 2006, 20, 1211–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serinsöz, E.; Neusch, M.; Büsche, G.; Wasielewski, R.; Kreipe, H.; Bock, O. Aberrant expression of beta-catenin discriminates acute myeloid leukaemia from acute lymphoblastic leukaemia. Br. J. Haematol. 2004, 126, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Fathi, E.; Valipour, B.; Sanaat, Z.; Charoudeh, H.N.; Farahzadi, R. Interleukin-6, -8, and TGF-β Secreted from Mesenchymal Stem Cells Show Functional Role in Reduction of Telomerase Activity of Leukemia Cell Via Wnt5a/β-Catenin and P53 Pathways. Adv. Pharm. Bull. 2020, 10, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Müller-Tidow, C.; Steffen, B.; Cauvet, T.; Tickenbrock, L.; Ji, P.; Diederichs, S.; Sargin, B.; Koöhler, G.; Stelljes, M.; Puccetti, E.; et al. Translocation Products in Acute Myeloid Leukemia Activate the Wnt Signaling Pathway in Hematopoietic Cells. Mol. Cell. Biol. 2004, 24, 2890–2904. [Google Scholar] [CrossRef] [Green Version]
- Heidel, F.H.; Bullinger, L.; Feng, Z.; Wang, Z.; Neff, T.; Stein, L.; Kalaitzidis, D.; Lane, S.W.; Armstrong, S.A. Genetic and Pharmacologic Inhibition of β-Catenin Targets Imatinib-Resistant Leukemia Stem Cells in CML. Cell Stem Cell 2012, 10, 412–424. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, L.; Mahajan, S.; Capietto, A.-H.; Yang, Z.; Zamani, A.; Ricci, B.; Bumpass, D.; Meyer, M.A.; Su, X.; Wang-Gillam, A.; et al. Dickkopf-related protein 1 (Dkk1) regulates the accumulation and function of myeloid derived suppressor cells in cancer. J. Cell Biol. 2016, 213, 2131. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, S.; Chaklader, M.; Law, S. Aberrant Wnt Signaling Pathway in the Hematopoietic Stem/Progenitor Compartment in Experimental Leukemic Animal. J. Cell Commun. Signal. 2019, 13, 39–52. [Google Scholar] [CrossRef]
- Yu, J.; Chen, L.; Cui, B.; Widhopf, G.F., II; Shen, Z.; Wu, R.; Zhang, L.; Zhang, S.; Briggs, S.P.; Kipps, T.J. Wnt5a induces ROR1/ROR2 heterooligomerization to enhance leukemia chemotaxis and proliferation. J. Clin. Investig. 2016, 126, 585–598. [Google Scholar] [CrossRef] [Green Version]
- Rozovski, U.; Harris, D.M.; Li, P.; Liu, Z.; Jain, P.; Ferrajoli, A.; Burger, J.A.; Bose, P.; Thompson, P.A.; Jain, N.; et al. STAT3-Induced Wnt5a Provides Chronic Lymphocytic Leukemia Cells with Survival Advantage. J. Immunol. 2019, 203, 3078–3085. [Google Scholar] [CrossRef]
- Hartmann, T.N.; Grabovsky, V.; Wang, W.; Desch, P.; Rubenzer, G.; Wollner, S.; Binsky, I.; Vallon-Eberhard, A.; Sapoznikov, A.; Burger, M.; et al. Circulating B-Cell Chronic Lymphocytic Leukemia Cells Display Impaired Migration to Lymph Nodes and Bone Marrow. Cancer Res. 2009, 69, 3121–3130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Chen, L.; Chen, Y.; Hasan, M.K.; Ghia, E.M.; Zhang, L.; Wu, R.; Rassenti, L.Z.; Widhopf, G.F.; Shen, Z.; et al. Wnt5a induces ROR1 to associate with 14-3-3ζ for enhanced chemotaxis and proliferation of chronic lymphocytic leukemia cells. Leukemia 2017, 31, 2608–2614. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.K.; Yu, J.; Chen, L.; Cui, B.; Ii, G.F.W.; Rassenti, L.; Shen, Z.; Briggs, S.P.; Kipps, T.J. Wnt5a induces ROR1 to complex with HS1 to enhance migration of chronic lymphocytic leukemia cells. Leukemia 2017, 31, 2615–2622. [Google Scholar] [CrossRef] [Green Version]
- Daneshmanesh, A.H.; Hojjat-Farsangi, M.; Moshfegh, A.; Khan, S.; Mikaelsson, E.; Österborg, A.; Mellstedt, H. The PI3K/AKT/mTOR pathway is involved in direct apoptosis of CLL cells induced by ROR1 monoclonal antibodies. Br. J. Haematol. 2015, 169, 455–458. [Google Scholar] [CrossRef] [Green Version]
- Cui, B.; Ghia, E.M.; Chen, L.; Rassenti, L.Z.; DeBoever, C.; Widhopf, G.F.; Yu, J.; Neuberg, D.S.; Wierda, W.G.; Rai, K.R.; et al. High-level ROR1 associates with accelerated disease progression in chronic lymphocytic leukemia. Blood 2016, 128, 2931–2940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janovská, P.; Bryja, V. Wnt signalling pathways in chronic lymphocytic leukaemia and B-cell lymphomas. Br. J. Pharmacol. 2017, 174, 4701–4715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Chen, L.; Yu, J.; Ghia, E.M.; Choi, M.Y.; Zhang, L.; Zhang, S.; Sánchez-López, E.; Widhopf, G.F.; Messer, K.; et al. Cirmtuzumab blocks Wnt5a/ROR1 stimulation of NF-κB to repress autocrine STAT3 activation in chronic lymphocytic leukemia. Blood 2019, 134, 1084–1094. [Google Scholar] [CrossRef]
- Memarian, A.; Vosough, P.; Asgarian-Omran, H.; Tabrizi, M.; Shabani, M.; Shokri, F. Differential WNT gene expression in various subtypes of acute lymphoblastic leukemia. Iran. J. Immunol. 2012, 9, 61–71. [Google Scholar]
- Memarian, A.; Hojjat-Farsangi, M.; Asgarian-Omran, H.; Younesi, V.; Jeddi-Tehrani, M.; Sharifian, R.A.; Khoshnoodi, J.; Razavi, S.M.; Rabbani, H.; Shokri, F. Variation in WNT genes expression in different subtypes of chronic lymphocytic leukemia. Leuk. Lymphoma 2009, 50, 2061–2070. [Google Scholar] [CrossRef]
- Bicocca, V.T.; Chang, B.H.; Masouleh, B.K.; Müschen, M.; Loriaux, M.M.; Druker, B.J.; Tyner, J.W. Crosstalk between ROR1 and the Pre-B Cell Receptor Promotes Survival of t(1;19) Acute Lymphoblastic Leukemia. Cancer Cell 2012, 22, 656–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Harris, D.; Liu, Z.; Liu, J.; Keating, M.; Estrov, Z. Stat3 Activates the Receptor Tyrosine Kinase Like Orphan Receptor-1 Gene in Chronic Lymphocytic Leukemia Cells. PLoS ONE 2010, 5, e11859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karvonen, H.; Perttilä, R.; Niininen, W.; Hautanen, V.; Barker, H.; Murumägi, A.; Heckman, C.A.; Ungureanu, D. Wnt5a and ROR1 activate non-canonical Wnt signaling via RhoA in TCF3-PBX1 acute lymphoblastic leukemia and highlight new treatment strategies via Bcl-2 co-targeting. Oncogene 2019, 38, 3288–3300. [Google Scholar] [CrossRef] [PubMed]
- Karvonen, H.; Chiron, D.; Niininen, W.; Ek, S.; Jerkeman, M.; Moradi, E.; Nykter, M.; Heckman, C.A.; Kallioniemi, O.; Murumägi, A.; et al. Crosstalk between ROR1 and BCR pathways defines novel treatment strategies in mantle cell lymphoma. Blood Adv. 2017, 1, 2257–2268. [Google Scholar] [CrossRef] [Green Version]
- Rassenti, L.Z.; Ballati, V.; Ghia, E.M.; Palamarchuk, A.; Tomasello, L.; Fadda, P.; Pekarsky, Y.; Widhopf, G.F., II; Kipps, T.J.; Crace, C.M. Dysregulation to identify therapeutic target combinations for chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2017, 114, 10731–10736. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chen, Y.; Chen, L.; Zhang, L.; Rassenti, L.Z.; Widhopf, G.F.; Kipps, T.J. Cirmtuzumab inhibits ibrutinib-resistant, Wnt5a-induced Rac1 activation and proliferation in mantle cell lymphoma. Oncotarget 2018, 9, 24731–24736. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Fan, X.; Fu, B.; Zheng, M.; Zhang, A.; Zhong, K.; Yan, J.; Sun, R.; Tian, Z.; Wei, H. EpCAM Inhibition Sensitizes Chemoresistant Leukemia to Immune Surveillance. Cancer Res. 2017, 77, 482–493. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Zavala, M.; Riveros-Magaña, A.R.; García-Castro, B.; Barrera-Chairez, E.; Rubio-Jurado, B.; Garcés-Ruíz, O.M.; Ramos-Solano, M.; Aguilar-Lemarroy, A.; Jave-Suárez, L.F. WNT receptors profile expression in mature blood cells and immature leukemic cells: RYK emerges as a hallmark receptor of acute leukemia. Eur. J. Haematol. 2016, 97, 155–165. [Google Scholar] [CrossRef]
- Hofbauer, S.W.; Kren, P.W.; Ganghammer, S.; Asslaber, D.; Pichler, V.; Oberascher, K.; Henschler, R.; Wallner, M.; Kerschbaum, H.; Grei, R. Tiam1/Rac1 signals contribute to the proliferation and chemoresistance, but not motility, of chronic lymphocytic leukemia cells. Blood 2014, 123, 2181–2188. [Google Scholar] [CrossRef] [Green Version]
- Kaucka, M.; Plevova, K.; Pavlova, S.; Janovská, P.; Mishra, A.; Verner, J.; Prochazkova, J.; Krejci, P.; Kotaŝková, J.; Ovesná, P.; et al. The Planar Cell Polarity Pathway Drives Pathogenesis of Chronic Lymphocytic Leukemia by the Regulation of B-Lymphocyte Migration. Cancer Res. 2013, 73, 1491–1501. [Google Scholar] [CrossRef] [Green Version]
- Voronkov, A.; Krauss, S. Wnt/beta-catenin signaling and small molecule inhibitors. Curr. Pharm. Des. 2013, 19, 634–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kida, A.; Kahn, M. Hypoxia selects for a quiescent, CML stem/leukemia initiating-like population dependent on CBP/catenin transcription. Curr. Mol. Pharmacol. 2013, 6, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Gang, E.J.; Hsieh, Y.-T.; Pham, J.; Zhao, Y.; Nguyen, C.; Huantes, S.; Park, E.; Naing, K.; Klemm, L.; Swaminathan, S.; et al. Small-molecule inhibition of CBP/catenin interactions eliminates drug-resistant clones in acute lymphoblastic leukemia. Oncogene 2014, 33, 2169–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emami, K.H.; Nguyen, C.; Ma, H.; Kim, D.H.; Jeong, K.W.; Eguchi, M.; Moon, R.T.; Teo, J.-L.; Kim, H.Y.; Moon, S.H.; et al. A small molecule inhibitor of beta-catenin/CREB-binding protein transcription [corrected]. Proc. Natl. Acad. Sci. USA 2004, 101, 12682–12687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.Y.; Widhopf, G.F.; Wu, C.C.; Cui, B.; Lao, F.; Sadarangani, A.; Cavagnaro, J.; Prussak, C.; Carson, D.A.; Jamieson, C.; et al. Pre-clinical Specificity and Safety of UC-961, a First-In-Class Monoclonal Antibody Targeting ROR. Clin. Lymphoma Myeloma Leuk. 2015, 15, S167–S169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudecek, M.; Schmitt, T.M.; Baskar, S.; Lupo-Stanghellini, M.T.; Nishida, T.; Yamamoto, T.N.; Bleakley, M.; Turtle, C.J.; Chang, W.-C.; Greisman, H.A.; et al. The B-cell tumor–associated antigen ROR1 can be targeted with T cells modified to express a ROR1-specific chimeric antigen receptor. Blood 2010, 116, 4532–4541. [Google Scholar] [CrossRef] [Green Version]
- Hudecek, M.; Lupo-Stanghellini, M.-T.; Kosasih, P.L.; Sommermeyer, D.; Jensen, M.C.; Rader, C.; Riddell, S.R. Receptor affinity and extracellular domain modifications affect tumor recognition by ROR1-specific chimeric antigen receptor T cells. Clin. Cancer Res. 2013, 19, 3153–3164. [Google Scholar] [CrossRef] [Green Version]
- Berger, C.; Sommermeyer, D.; Hudecek, M.; Berger, M.; Balakrishnan, A.; Paszkiewicz, P.J.; Kosasih, P.L.; Rader, C.; Riddell, S.R. Safety of Targeting ROR1 in Primates with Chimeric Antigen Receptor–Modified T Cells. Cancer Immunol. Res. 2014, 3, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Halford, M.M.; Macheda, M.; Parish, C.L.; Takano, E.A.; Fox, S.; Layton, D.; Nice, E.; Stacker, S.A. A Fully Human Inhibitory Monoclonal Antibody to the Wnt Receptor RYK. PLoS ONE 2013, 8, e75447. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastelaro de Rezende, M.; Zenker Justo, G.; Julian Paredes-Gamero, E.; Gosens, R. Wnt-5A/B Signaling in Hematopoiesis throughout Life. Cells 2020, 9, 1801. https://doi.org/10.3390/cells9081801
Mastelaro de Rezende M, Zenker Justo G, Julian Paredes-Gamero E, Gosens R. Wnt-5A/B Signaling in Hematopoiesis throughout Life. Cells. 2020; 9(8):1801. https://doi.org/10.3390/cells9081801
Chicago/Turabian StyleMastelaro de Rezende, Marina, Giselle Zenker Justo, Edgar Julian Paredes-Gamero, and Reinoud Gosens. 2020. "Wnt-5A/B Signaling in Hematopoiesis throughout Life" Cells 9, no. 8: 1801. https://doi.org/10.3390/cells9081801
APA StyleMastelaro de Rezende, M., Zenker Justo, G., Julian Paredes-Gamero, E., & Gosens, R. (2020). Wnt-5A/B Signaling in Hematopoiesis throughout Life. Cells, 9(8), 1801. https://doi.org/10.3390/cells9081801