Circulating MicroRNAs from Serum Exosomes May Serve as a Putative Biomarker in the Diagnosis and Treatment of Patients with Focal Cortical Dysplasia

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Isolation of Exosomes and RNA Extraction
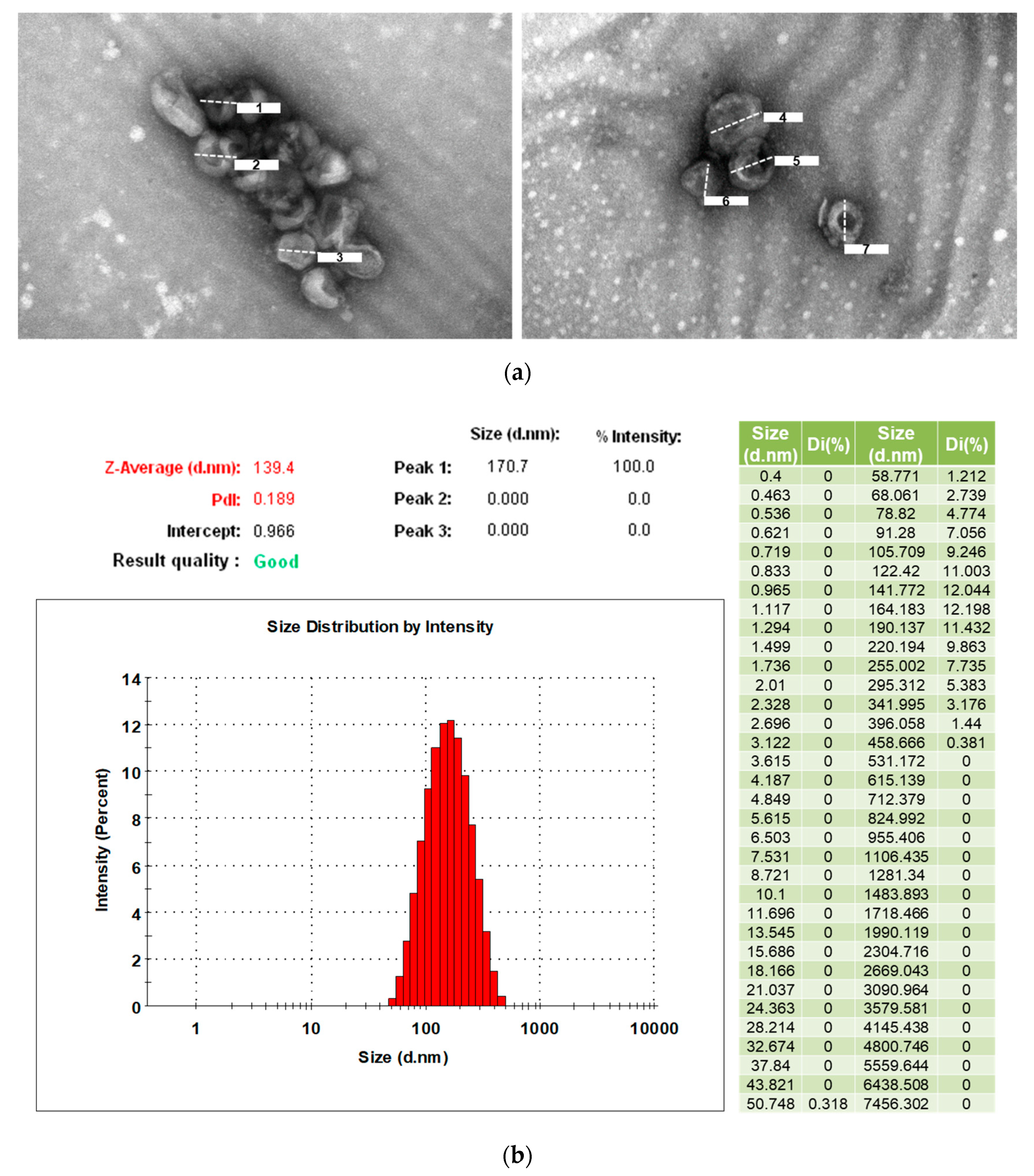
2.3. Transmission Electron Microscopy for Identification of Exosomes
2.4. Measurement of Particle Size and Distribution of Exosomes
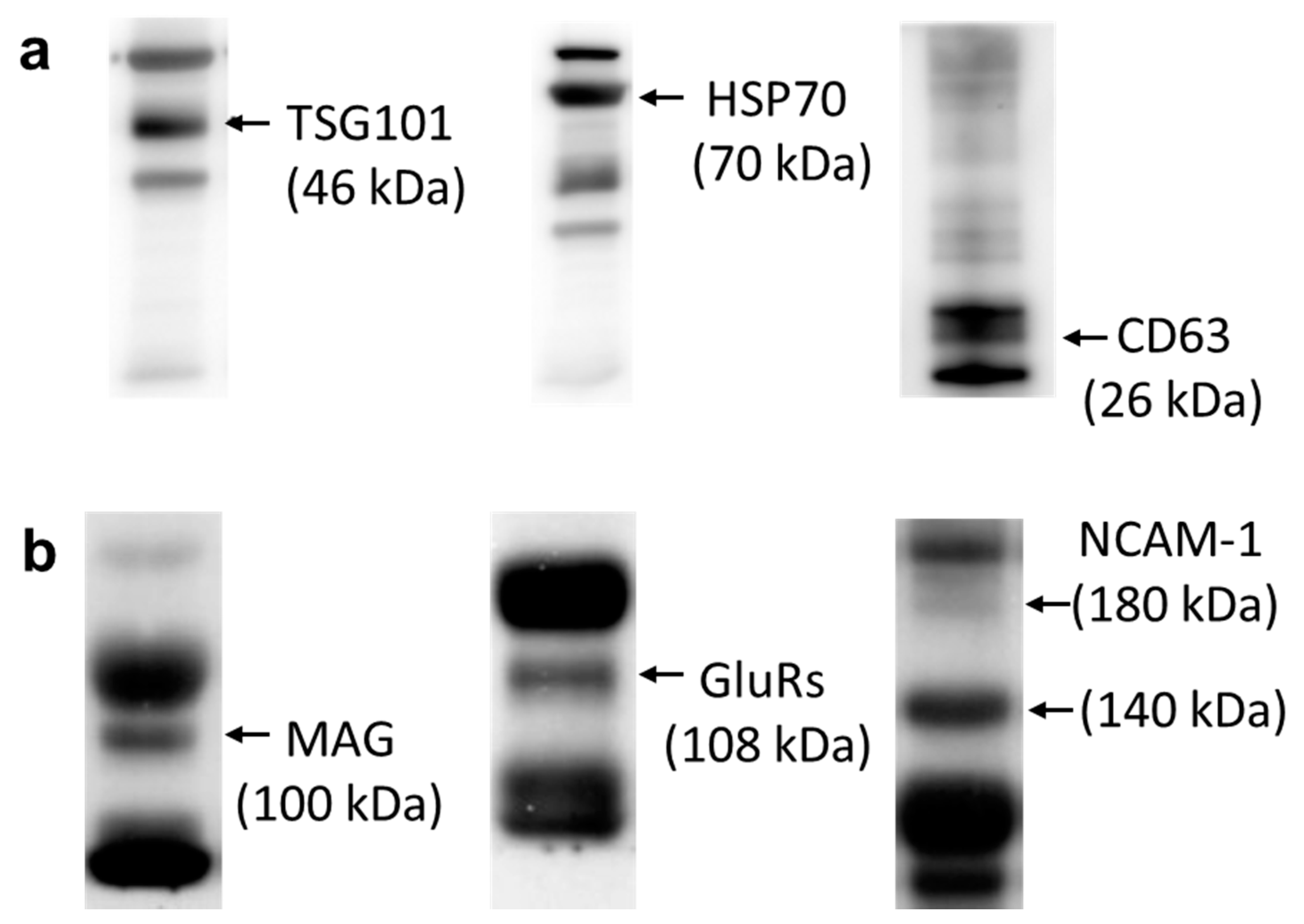
2.5. Western Blot Analysis
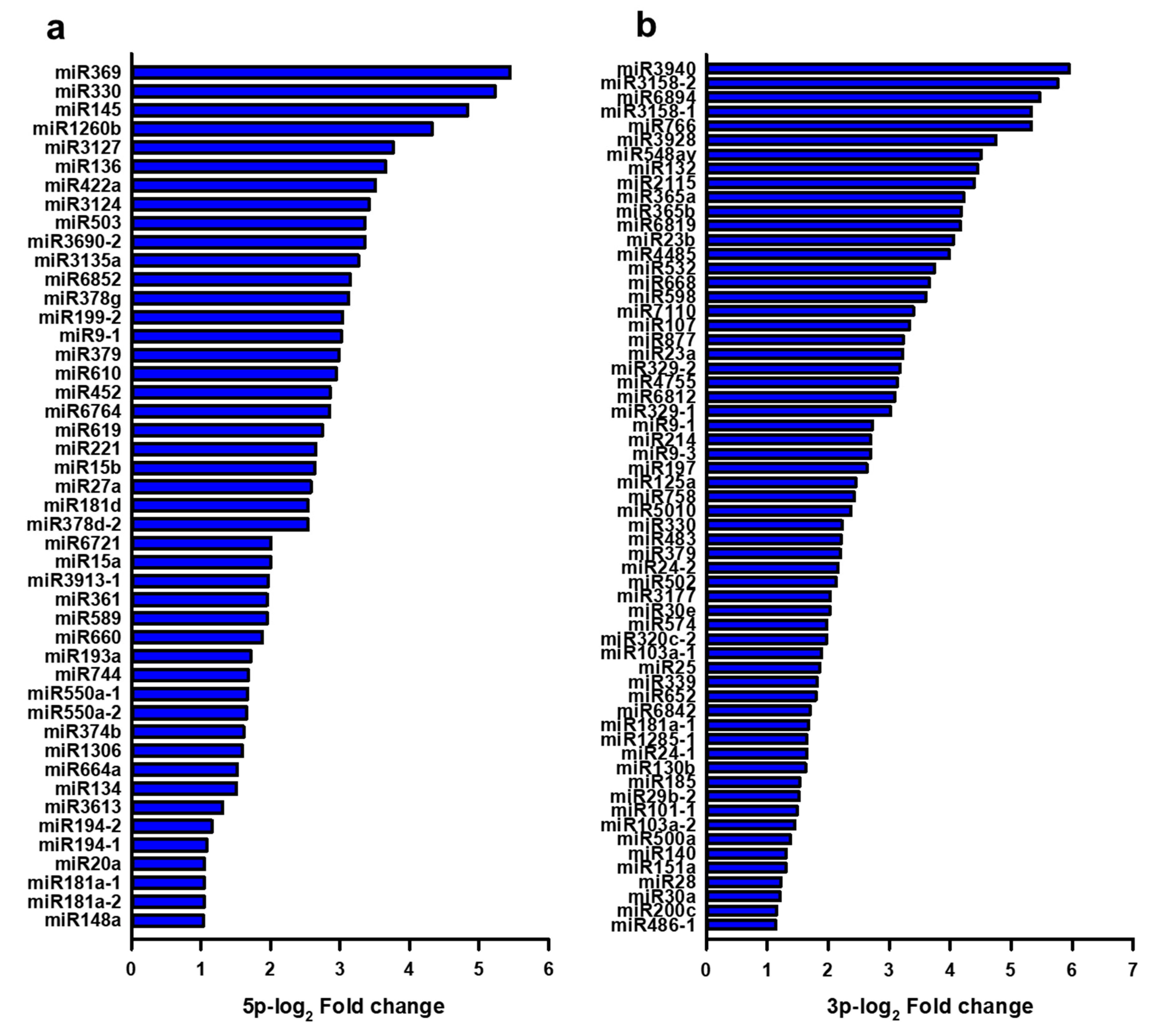
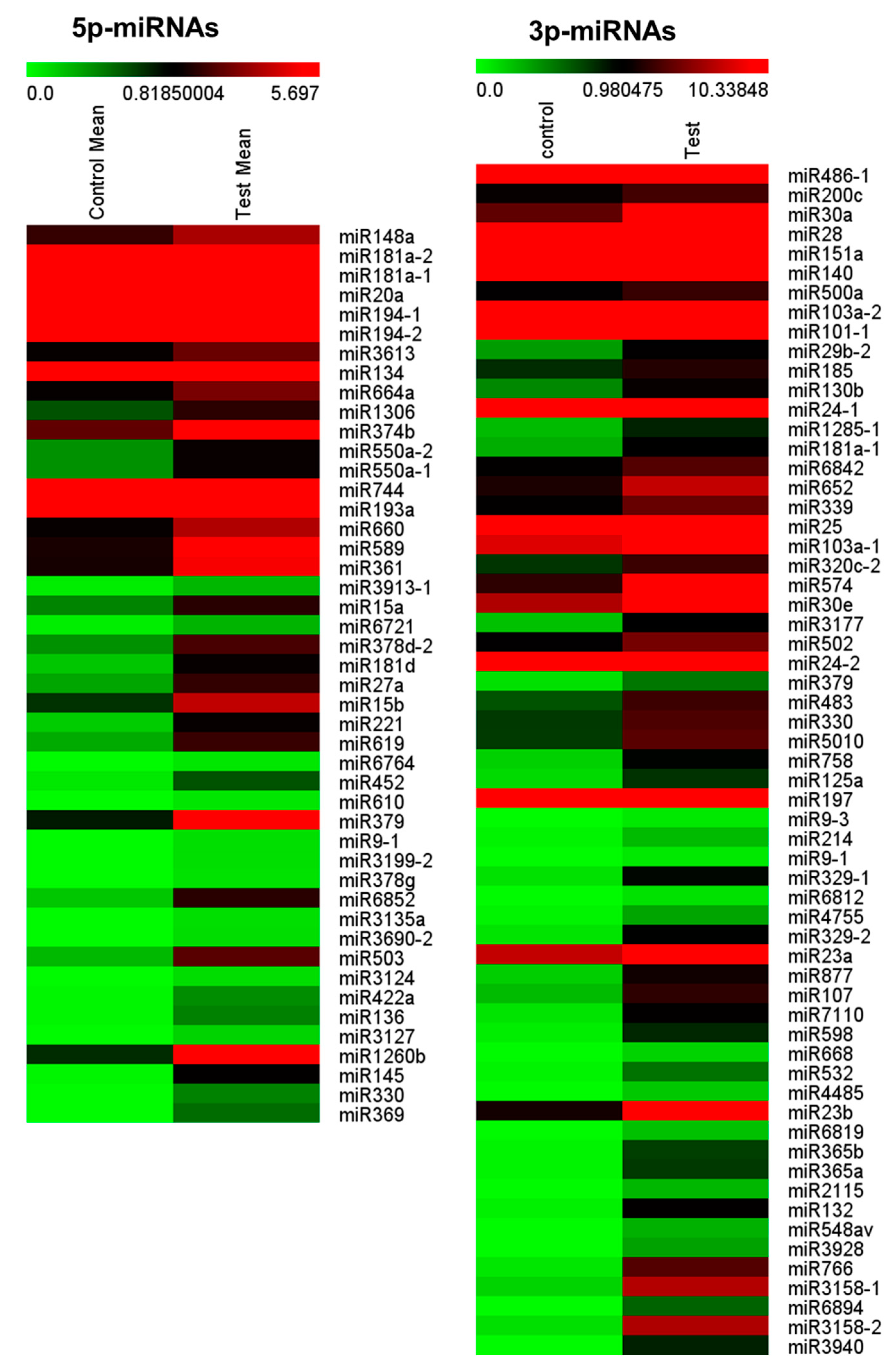
2.6. Next-Generation Sequencing Analysis
2.7. Analyses Using MicroRNA Target Predicting Databases
2.8. Analysis of Disease-Related Pathways from Predicted MicroRNA and Pathway Category
3. Results
3.1. Characteristics and Demographic Data
3.2. Characterization of Exosomes from Human Serum
3.3. Expressional Changes of MicroRNA in Serum Exosomes from Patients with Focal Cortical Dysplasia
3.4. Identification of Target MicroRNAs Related to Focal Cortical Dysplasia and Epilepsy
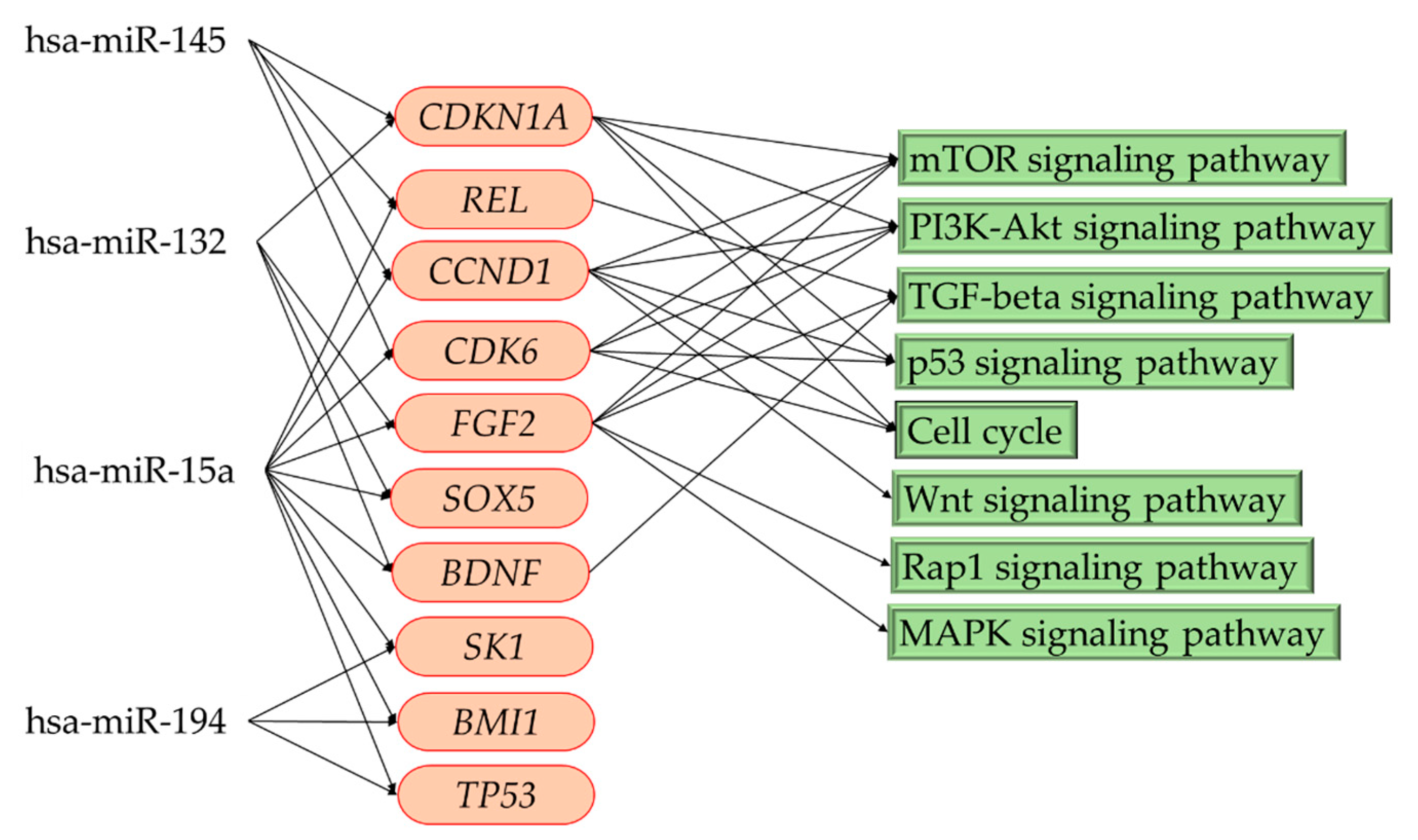
3.5. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis and Pathway Category
3.6. Related Pathways in Patients with Focal Cortical Dysplasia
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crino, P.B. Focal Cortical Dysplasia. Semin. Neurol. 2015, 35, 201–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desikan, R.S.; Barkovich, A.J. Malformations of cortical development. Ann. Neurol. 2016, 80, 797–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najm, I.M.; Sarnat, H.B.; Blumcke, I. Review: The international consensus classification of Focal Cortical Dysplasia—A critical update 2018. Neuropathol. Appl. Neurobiol. 2018, 44, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Kabat, J.; Krol, P. Focal cortical dysplasia—Review. Pol. J. Radiol. 2012, 77, 35–43. [Google Scholar] [CrossRef]
- Sisodiya, S.M. Malformations of cortical development: Burdens and insights from important causes of human epilepsy. Lancet Neurol. 2004, 3, 29–38. [Google Scholar] [CrossRef]
- Adler, S.; Lorio, S.; Jacques, T.S.; Benova, B.; Gunny, R.; Cross, J.H.; Baldeweg, T.; Carmichael, D.W. Towards in vivo focal cortical dysplasia phenotyping using quantitative MRI. Neuroimage Clin. 2017, 15, 95–105. [Google Scholar] [CrossRef]
- Chang, W.S.; Nakajima, M.; Ochi, A.; Widjaja, E.; Rutka, J.T.; Yau, I.; Baba, S.; Otsubo, H. Detection of epileptogenic focus using advanced dynamic statistical parametric mapping with magnetoencephalography in a patient with MRI-negative focal cortical dysplasia type IIB. J. Neurosurg. Pediatr. 2019. [Google Scholar] [CrossRef]
- Lorio, S.; Adler, S.; Gunny, R.; D’Arco, F.; Kaden, E.; Wagstyl, K.; Jacques, T.S.; Clark, C.A.; Cross, J.H.; Baldeweg, T.; et al. MRI profiling of focal cortical dysplasia using multi-compartment diffusion models. Epilepsia 2020, 61, 433–444. [Google Scholar] [CrossRef]
- Crino, P.B. Molecular pathogenesis of focal cortical dysplasia and hemimegalencephaly. J. Child. Neurol. 2005, 20, 330–336. [Google Scholar] [CrossRef]
- Iffland, P.H., 2nd; Crino, P.B. Focal Cortical Dysplasia: Gene Mutations, Cell Signaling, and Therapeutic Implications. Annu. Rev. Pathol. 2017, 12, 547–571. [Google Scholar] [CrossRef]
- Mirzaa, G.M.; Campbell, C.D.; Solovieff, N.; Goold, C.; Jansen, L.A.; Menon, S.; Timms, A.E.; Conti, V.; Biag, J.D.; Adams, C.; et al. Association of MTOR Mutations With Developmental Brain Disorders, Including Megalencephaly, Focal Cortical Dysplasia, and Pigmentary Mosaicism. JAMA Neurol. 2016, 73, 836–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Tomankova, T.; Petrek, M.; Kriegova, E. Involvement of microRNAs in physiological and pathological processes in the lung. Respir. Res. 2010, 11, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasu, S.; Kumano, K.; Darden, C.M.; Rahman, I.; Lawrence, M.C.; Naziruddin, B. MicroRNA Signatures as Future Biomarkers for Diagnosis of Diabetes States. Cells 2019, 8, 1533. [Google Scholar] [CrossRef] [Green Version]
- Ferrero, G.; Cordero, F.; Tarallo, S.; Arigoni, M.; Riccardo, F.; Gallo, G.; Ronco, G.; Allasia, M.; Kulkarni, N.; Matullo, G.; et al. Small non-coding RNA profiling in human biofluids and surrogate tissues from healthy individuals: Description of the diverse and most represented species. Oncotarget 2018, 9, 3097–3111. [Google Scholar] [CrossRef] [Green Version]
- Yeri, A.; Courtright, A.; Reiman, R.; Carlson, E.; Beecroft, T.; Janss, A.; Siniard, A.; Richholt, R.; Balak, C.; Rozowsky, J.; et al. Total Extracellular Small RNA Profiles from Plasma, Saliva, and Urine of Healthy Subjects. Sci. Rep. 2017, 7, 44061. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, Y.H.; Wang, H. Exploring MicroRNA Biomarkers for Parkinson’s Disease from mRNA Expression Profiles. Cells 2018, 7, 245. [Google Scholar] [CrossRef] [Green Version]
- Saghazadeh, A.; Rezaei, N. MicroRNA machinery in Parkinson’s disease: A platform for neurodegenerative diseases. Expert Rev. Neurother. 2015. [Google Scholar] [CrossRef]
- Denk, J.; Boelmans, K.; Siegismund, C.; Lassner, D.; Arlt, S.; Jahn, H. MicroRNA Profiling of CSF Reveals Potential Biomarkers to Detect Alzheimer`s Disease. PLoS ONE 2015, 10, e0126423. [Google Scholar] [CrossRef] [Green Version]
- Ricci, C.; Marzocchi, C.; Battistini, S. MicroRNAs as Biomarkers in Amyotrophic Lateral Sclerosis. Cells 2018, 7, 219. [Google Scholar] [CrossRef] [Green Version]
- Eyileten, C.; Wicik, Z.; De Rosa, S.; Mirowska-Guzel, D.; Soplinska, A.; Indolfi, C.; Jastrzebska-Kurkowska, I.; Czlonkowska, A.; Postula, M. MicroRNAs as Diagnostic and Prognostic Biomarkers in Ischemic Stroke-A Comprehensive Review and Bioinformatic Analysis. Cells 2018, 7, 249. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.; Dixit, A.B.; Banerjee, J.; Tripathi, M.; Sarat Chandra, P. Role of inflammation and its miRNA based regulation in epilepsy: Implications for therapy. Clin. Chim. Acta 2016, 452, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bielefeld, P.; Mooney, C.; Henshall, D.C.; Fitzsimons, C.P. miRNA-Mediated Regulation of Adult Hippocampal Neurogenesis; Implications for Epilepsy. Brain Plast. 2017, 3, 43–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demos, M.; Guella, I.; DeGuzman, C.; McKenzie, M.B.; Buerki, S.E.; Evans, D.M.; Toyota, E.B.; Boelman, C.; Huh, L.L.; Datta, A.; et al. Diagnostic Yield and Treatment Impact of Targeted Exome Sequencing in Early-Onset Epilepsy. Front. Neurol. 2019, 10, 434. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y. The Challenge of microRNA as a Biomarker of Epilepsy. Curr. Neuropharmacol. 2018, 16, 37–42. [Google Scholar] [CrossRef]
- Zhan, L.; Yao, Y.; Fu, H.; Li, Z.; Wang, F.; Zhang, X.; He, W.; Zheng, W.; Zhang, Y.; Zheng, H. Protective role of miR-23b-3p in kainic acid-induced seizure. Neuroreport 2016, 27, 764–768. [Google Scholar] [CrossRef]
- Zheng, H.; Tang, R.; Yao, Y.; Ji, Z.; Cao, Y.; Liu, Z.; Peng, F.; Wang, W.; Can, D.; Xing, H.; et al. MiR-219 Protects Against Seizure in the Kainic Acid Model of Epilepsy. Mol. Neurobiol. 2016, 53, 1–7. [Google Scholar] [CrossRef]
- Haenisch, S.; von Ruden, E.L.; Wahmkow, H.; Rettenbeck, M.L.; Michler, C.; Russmann, V.; Bruckmueller, H.; Waetzig, V.; Cascorbi, I.; Potschka, H. miRNA-187-3p-Mediated Regulation of the KCNK10/TREK-2 Potassium Channel in a Rat Epilepsy Model. ACS Chem. Neurosci. 2016, 7, 1585–1594. [Google Scholar] [CrossRef]
- Wang, W.; Guo, Y.; He, L.; Chen, C.; Luo, J.; Ma, Y.; Li, J.; Yang, Y.; Yang, Q.; Du, C.; et al. Overexpression of miRNA-137 in the brain suppresses seizure activity and neuronal excitability: A new potential therapeutic strategy for epilepsy. Neuropharmacology 2018, 138, 170–181. [Google Scholar] [CrossRef]
- Surges, R.; Kretschmann, A.; Abnaof, K.; van Rikxoort, M.; Ridder, K.; Frohlich, H.; Danis, B.; Kaminski, R.M.; Foerch, P.; Elger, C.E.; et al. Changes in serum miRNAs following generalized convulsive seizures in human mesial temporal lobe epilepsy. Biochem. Biophys. Res. Commun. 2016, 481, 13–18. [Google Scholar] [CrossRef]
- Jimenez-Mateos, E.M.; Bray, I.; Sanz-Rodriguez, A.; Engel, T.; McKiernan, R.C.; Mouri, G.; Tanaka, K.; Sano, T.; Saugstad, J.A.; Simon, R.P.; et al. miRNA Expression profile after status epilepticus and hippocampal neuroprotection by targeting miR-132. Am. J. Pathol. 2011, 179, 2519–2532. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Rahmati, S.; Shojaei, F.; Shojaeian, A.; Rezakhani, L.; Dehkordi, M.B. An overview of current knowledge in biological functions and potential theragnostic applications of exosomes. Chem. Phys. Lipids 2020, 226, 104836. [Google Scholar] [CrossRef] [PubMed]
- Jan, A.T.; Rahman, S.; Khan, S.; Tasduq, S.A.; Choi, I. Biology, Pathophysiological Role, and Clinical Implications of Exosomes: A Critical Appraisal. Cells 2019, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruivo, C.F.; Adem, B.; Silva, M.; Melo, S.A. The Biology of Cancer Exosomes: Insights and New Perspectives. Cancer Res. 2017, 77, 6480–6488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhome, R.; Del Vecchio, F.; Lee, G.H.; Bullock, M.D.; Primrose, J.N.; Sayan, A.E.; Mirnezami, A.H. Exosomal microRNAs (exomiRs): Small molecules with a big role in cancer. Cancer Lett. 2018, 420, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zhang, H.; Xie, W.; Meng, F.; Zhang, K.; Jiang, Y.; Zhang, X.; Zhang, J. Altered microRNA profiles in plasma exosomes from mesial temporal lobe epilepsy with hippocampal sclerosis. Oncotarget 2017, 8, 4136–4146. [Google Scholar] [CrossRef]
- Shi, M.; Sheng, L.; Stewart, T.; Zabetian, C.P.; Zhang, J. New windows into the brain: Central nervous system-derived extracellular vesicles in blood. Prog. Neurobiol. 2019, 175, 96–106. [Google Scholar] [CrossRef]
- Fisher, R.S. The New Classification of Seizures by the International League against Epilepsy 2017. Curr. Neurol. NeuroSci. Rep. 2017, 17, 48. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Fruhbeis, C.; Frohlich, D.; Kramer-Albers, E.M. Emerging roles of exosomes in neuron-glia communication. Front. Physiol. 2012, 3, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faure, J.; Lachenal, G.; Court, M.; Hirrlinger, J.; Chatellard-Causse, C.; Blot, B.; Grange, J.; Schoehn, G.; Goldberg, Y.; Boyer, V.; et al. Exosomes are released by cultured cortical neurones. Mol. Cell Neurosci. 2006, 31, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Fiandaca, M.S.; Kapogiannis, D.; Mapstone, M.; Boxer, A.; Eitan, E.; Schwartz, J.B.; Abner, E.L.; Petersen, R.C.; Federoff, H.J.; Miller, B.L.; et al. Identification of preclinical Alzheimer’s disease by a profile of pathogenic proteins in neurally derived blood exosomes: A case-control study. Alzheimers Dement. 2015, 11, 600–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.J.; Chang, W.A.; Liao, S.H.; Chang, K.F.; Sheu, C.C.; Kuo, P.L. The Effects of Epigallocatechin Gallate (EGCG) on Pulmonary Fibroblasts of Idiopathic Pulmonary Fibrosis (IPF)-A Next-Generation Sequencing and Bioinformatic Approach. Int. J. Mol. Sci. 2019, 20, 1958. [Google Scholar] [CrossRef] [Green Version]
- Kal, A.J.; van Zonneveld, A.J.; Benes, V.; van den Berg, M.; Koerkamp, M.G.; Albermann, K.; Strack, N.; Ruijter, J.M.; Richter, A.; Dujon, B.; et al. Dynamics of gene expression revealed by comparison of serial analysis of gene expression transcript profiles from yeast grown on two different carbon sources. Mol. Biol. Cell 1999, 10, 1859–1872. [Google Scholar] [CrossRef]
- Williams, J.R.; Yang, R.; Clifford, J.L.; Watson, D.; Campbell, R.; Getnet, D.; Kumar, R.; Hammamieh, R.; Jett, M. Functional Heatmap: An automated and interactive pattern recognition tool to integrate time with multi-omics assays. BMC Bioinform. 2019, 20, 81. [Google Scholar] [CrossRef]
- Huang, Z.; Shi, J.; Gao, Y.; Cui, C.; Zhang, S.; Li, J.; Zhou, Y.; Cui, Q. HMDD v3.0: A database for experimentally supported human microRNA-disease associations. Nucleic Acids Res. 2019, 47, D1013–D1017. [Google Scholar] [CrossRef] [Green Version]
- Das, S.S.; Saha, P.; Chakravorty, N. miRwayDB: A database for experimentally validated microRNA-pathway associations in pathophysiological conditions. Database (Oxford) 2018, 2018. [Google Scholar] [CrossRef]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Du, J.; Yuan, Z.; Ma, Z.; Song, J.; Xie, X.; Chen, Y. KEGG-PATH: Kyoto encyclopedia of genes and genomes-based pathway analysis using a path analysis model. Mol. Biosyst. 2014, 10, 2441–2447. [Google Scholar] [CrossRef]
- Huang, R.; Grishagin, I.; Wang, Y.; Zhao, T.; Greene, J.; Obenauer, J.C.; Ngan, D.; Nguyen, D.T.; Guha, R.; Jadhav, A.; et al. The NCATS BioPlanet—An Integrated Platform for Exploring the Universe of Cellular Signaling Pathways for Toxicology, Systems Biology, and Chemical Genomics. Front. Pharmacol. 2019, 10, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Li, Z.; Zhao, N.; Xu, Y.; Liu, Y.; Zhou, Y.; Shang, D.; Qiu, F.; Zhang, R.; Chang, Z.; et al. Functional analysis of microRNA and transcription factor synergistic regulatory network based on identifying regulatory motifs in non-small cell lung cancer. BMC Syst. Biol. 2013, 7, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yu, J.T.; Tan, L.; Tian, Y.; Ma, J.; Tan, C.C.; Wang, H.F.; Liu, Y.; Tan, M.S.; Jiang, T.; et al. Genome-wide circulating microRNA expression profiling indicates biomarkers for epilepsy. Sci. Rep. 2015, 5, 9522. [Google Scholar] [CrossRef]
- Sun, J.; Cheng, W.; Liu, L.; Tao, S.; Xia, Z.; Qi, L.; Huang, M. Identification of serum miRNAs differentially expressed in human epilepsy at seizure onset and post-seizure. Mol. Med. Rep. 2016, 14, 5318–5324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korotkov, A.; Broekaart, D.W.M.; Banchaewa, L.; Pustjens, B.; van Scheppingen, J.; Anink, J.J.; Baayen, J.C.; Idema, S.; Gorter, J.A.; van Vliet, E.A.; et al. microRNA-132 is overexpressed in glia in temporal lobe epilepsy and reduces the expression of pro-epileptogenic factors in human cultured astrocytes. Glia 2020, 68, 60–75. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Wang, H.; Wang, Q.; Chen, Y.; Chen, S. Expression of p-CREB and activity-dependent miR-132 in temporal lobe epilepsy. Int. J. Clin. Exp. Med. 2014, 7, 1297–1306. [Google Scholar]
- Antonio, L.G.L.; Freitas-Lima, P.; Pereira-da-Silva, G.; Assirati, J.A., Jr.; Matias, C.M.; Cirino, M.L.A.; Tirapelli, L.F.; Velasco, T.R.; Sakamoto, A.C.; Carlotti, C.G., Jr.; et al. Expression of MicroRNAs miR-145, miR-181c, miR-199a and miR-1183 in the Blood and Hippocampus of Patients with Mesial Temporal Lobe Epilepsy. J. Mol. Neurosci. 2019, 69, 580–587. [Google Scholar] [CrossRef]
- Tiwari, D.; Peariso, K.; Gross, C. MicroRNA-induced silencing in epilepsy: Opportunities and challenges for clinical application. Develop. Dyn. 2018, 247, 94–110. [Google Scholar] [CrossRef] [Green Version]
- Karnati, H.K.; Panigrahi, M.K.; Gutti, R.K.; Greig, N.H.; Tamargo, I.A. miRNAs: Key Players in Neurodegenerative Disorders and Epilepsy. J. Alzheimers Dis. 2015, 48, 563–580. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.; Wu, Y.; Wang, J.; Luo, Z.; Long, L.; Deng, N.; Ning, S.; Zeng, Y.; Long, H.; Xiao, B. Network and Pathway-Based Analysis of Single-Nucleotide Polymorphism of miRNA in Temporal Lobe Epilepsy. Mol. Neurobiol. 2019, 56, 7022–7031. [Google Scholar] [CrossRef]
- Manna, I.; Labate, A.; Borzi, G.; Mumoli, L.; Cavalli, S.M.; Sturniolo, M.; Quattrone, A.; Gambardella, A. An SNP site in pri-miR-124, a brain expressed miRNA gene, no contribution to mesial temporal lobe epilepsy in an Italian sample. Neurol. Sci. 2016, 37, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Henshall, D.C. MicroRNAs in the pathophysiology and treatment of status epilepticus. Front. Mol. Neurosci. 2013, 6, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, L.A.; Mirzaa, G.M.; Ishak, G.E.; O’Roak, B.J.; Hiatt, J.B.; Roden, W.H.; Gunter, S.A.; Christian, S.L.; Collins, S.; Adams, C.; et al. PI3K/AKT pathway mutations cause a spectrum of brain malformations from megalencephaly to focal cortical dysplasia. Brain 2015, 138, 1613–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Jiang, J.; Zheng, G.; Chen, J.; Lu, H.; Guo, H.; Wu, C. miR-139-5p modulates cortical neuronal migration by targeting Lis1 in a rat model of focal cortical dysplasia. Int. J. Mol. Med. 2014, 33, 1407–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Sun, Y.; Tan, Z.; Che, N.; Ji, A.; Luo, X.; Sun, X.; Li, X.; Yang, K.; Wang, G.; et al. Serum MicroRNA-4521 is a Potential Biomarker for Focal Cortical Dysplasia with Refractory Epilepsy. Neurochem. Res. 2016, 41, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Che, N.; Zu, G.; Zhou, T.; Wang, X.; Sun, Y.; Tan, Z.; Liu, Y.; Wang, D.; Luo, X.; Zhao, Z.; et al. Aberrant Expression of miR-323a-5p in Patients with Refractory Epilepsy Caused by Focal Cortical Dysplasia. Genet. Test. Mol. Biomark. 2017, 21, 3–9. [Google Scholar] [CrossRef]
- D’Gama, A.M.; Woodworth, M.B.; Hossain, A.A.; Bizzotto, S.; Hatem, N.E.; LaCoursiere, C.M.; Najm, I.; Ying, Z.; Yang, E.; Barkovich, A.J.; et al. Somatic Mutations Activating the mTOR Pathway in Dorsal Telencephalic Progenitors Cause a Continuum of Cortical Dysplasias. Cell Rep. 2017, 21, 3754–3766. [Google Scholar] [CrossRef] [Green Version]
- Marsan, E.; Baulac, S. Review: Mechanistic target of rapamycin (mTOR) pathway, focal cortical dysplasia and epilepsy. Neuropathol. Appl. Neurobiol. 2018, 44, 6–17. [Google Scholar] [CrossRef]
- Berdichevsky, Y.; Dryer, A.M.; Saponjian, Y.; Mahoney, M.M.; Pimentel, C.A.; Lucini, C.A.; Usenovic, M.; Staley, K.J. PI3K-Akt signaling activates mTOR-mediated epileptogenesis in organotypic hippocampal culture model of post-traumatic epilepsy. J. Neurosci. 2013, 33, 9056–9067. [Google Scholar] [CrossRef]
- Kastenhuber, E.R.; Lowe, S.W. Putting p53 in Context. Cell 2017, 170, 1062–1078. [Google Scholar] [CrossRef] [Green Version]
- Simabuco, F.M.; Morale, M.G.; Pavan, I.C.B.; Morelli, A.P.; Silva, F.R.; Tamura, R.E. p53 and metabolism: From mechanism to therapeutics. Oncotarget 2018, 9, 23780–23823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaika, A.I.; Wei, J.; Noto, J.M.; Peek, R.M. Microbial Regulation of p53 Tumor Suppressor. PLoS Pathog. 2015, 11, e1005099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bizzotto, S.; Francis, F. Morphological and functional aspects of progenitors perturbed in cortical malformations. Front. Cell Neurosci. 2015, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Zhang, C.; Shu, H.; Wion, D.; Yang, H. Cortical dysplasia: A possible substrate for brain tumors. Future Oncol. 2012, 8, 251–258. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, S.S.; Wong, M. Therapeutic role of mammalian target of rapamycin (mTOR) inhibition in preventing epileptogenesis. Neurosci. Lett. 2011, 497, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Huang, B.; Wang, H.Y.; Chang, A.; Zheng, X.F.S. Emerging Role of MicroRNAs in mTOR Signaling. Cell Mol. Life Sci. 2017, 74, 2613–2625. [Google Scholar] [CrossRef]
- Dehay, C.; Kennedy, H. Cell-cycle control and cortical development. Nat. Rev. Neurosci. 2007, 8, 438–450. [Google Scholar] [CrossRef]
- Kashima, R.; Hata, A. The role of TGF-beta superfamily signaling in neurological disorders. Acta Biochim. Biophys Sin. (Shanghai) 2018, 50, 106–120. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.; Dixit, A.B.; Paul, D.; Tripathi, M.; Sarkar, C.; Chandra, P.S.; Banerjee, J. Comparative analysis of cytokine/chemokine regulatory networks in patients with hippocampal sclerosis (HS) and focal cortical dysplasia (FCD). Sci. Rep. 2017, 7, 15904. [Google Scholar] [CrossRef] [Green Version]
- Weissberg, I.; Wood, L.; Kamintsky, L.; Vazquez, O.; Milikovsky, D.Z.; Alexander, A.; Oppenheim, H.; Ardizzone, C.; Becker, A.; Frigerio, F.; et al. Albumin induces excitatory synaptogenesis through astrocytic TGF-beta/ALK5 signaling in a model of acquired epilepsy following blood-brain barrier dysfunction. Neurobiol. Dis. 2015, 78, 115–125. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Sex/Age | Seizure Semiology | Family History | MRI Finding | AEDs |
|---|---|---|---|---|---|
| 1 | F/39 | Focal to bilateral tonic‒clonic seizure | - | Multiple bilateral subcortical heterotopia; right frontal FCD | CBZ, LTG |
| 2 | M/40 | Focal onset clonic seizure | - | Bilateral frontoparietal and occipital heterotopia | LEV, ZNS, PER, LCM |
| 3 | M/29 | Focal to bilateral automatic seizure | - | Right parietal FCD | OXC, PER |
| 4 | F/34 | Focal impaired awareness clonic seizure | - | Right temporo-parietal FCD | LTG, LCM |
| 5 | M/36 | Focal impaired awareness clonic seizure | - | Right frontoparietal FCD | LTG, PER, LCM |
| 6 | M/63 | Focal impaired awareness dyscognitive seizure | - | Left frontoparietal lobe and subcortical band heterotopia | TPM, LEV |
| 7 | M/31 | Focal to bilateral tonic‒clonic seizure | - | Left temporal lobe button of sulcus heterotopia | CBZ, LTG, PHT |
| 8 | F/18 | Focal impaired awareness clonic seizure | + | Right frontoparietal lobe heterotopia | LEV, ZNS |
| 9 | F/47 | Focal to bilateral tonic‒clonic seizure | - | Right posterior tempo-parietal and subcortical band heterotopia | VPA, LEV, LCM |
| miRNA ID | Stem‒Loop | Expression | Evidence | Disease Publication | PMID |
|---|---|---|---|---|---|
| miR-194-2 | 5p | Up | Diagnostic biomarker | Epilepsy | 25825351 |
| miR-15a | 5p | Up | Diagnostic biomarker | Epilepsy Seizure onset and post-seizure | 25825351 27840934 |
| miR-132 | 3p | Up | Therapeutic target | Status epilepticus Temporal lobe epilepsy | 21945804 31408236 24995086 |
| miR-145 | 5p | Up | Circulating biomarker | Mesial temporal lobe epilepsy | 27833019 31368064 |
| MID: PubMed identification. | |||||
| miRNA | Regulation | Target Genes |
|---|---|---|
| hsa-miR-194 | Down | TP53, SKI, CARM1, BMI1, DNMT3A, CAV1, ICOSLG, IL10, YWHAE, SLC7A5, CPT1A, PEX26, AP1S1 |
| hsa-miR-15a | Down | USP8, CCND2, CHD4, DIAPH1, BSG, BRCA1, ZBTB18, GNAL, RS1, SLC7A5, IRF4, CCND1, CDK6, RASSF5, CDC42SE2, APP, RBPJ, OCRL, BHLHE40, GCLM, MYB, BMI1, PEX13, ACOX1, EIF2B2, AKT3, EN2, SBNO1, TRAK1, CYP26B1, MBD4, WNK3, CHEK1, BCL2, HNRNPA1, NUFIP2, IKBKG, MTHFR, IFNG, BACE1, PAK2, AP3M1, BAP1, B3GNT2, CLCN3, PDE4D, RPS6KA3, ITGA2, NR2C2, RIMS3, HMGA1, CADM1, TMEM245, YAP1, CASK, KPNA3, WT1, CDKN2B, KIF1A, TLL1, L2HGDH, PAG1, DICER1, HNRNPDL, GRB2KMT2D, PDIK1L, PI4K2B, CTNNA3, SCAMP5, EFNB2, GSK3B, ARHGDIA, KIF5B, PNPO, MYO5A, REL, CA8, UBE2H, ALDH3B1 |
| hsa-miR-132 | Down | RAB18, ADGRF4, GDF5, FGF2, CHRNA5, NBN, NLGN2, S100A9 CHL1, MMP9, STMN1, IRAK1, CDKN1A, MAPK1, RGMB, NCS1, LIFR RTN4, SOX5, SOX6, POLK, BDNF, KPNA1, SIRT1, GPR153, SMN1, B3GAT1 |
| hsa-miR-145 | Down | SOD2, IFNB1, CD44, MDM2, TGFBR2, NR1D2, HDAC2, JAG1, GGCX, CTGF, NTRK2, MAP3K3, SERPINE1, CDKN1A, IRS2, DUSP6, CCND1 TPM1, CDK6, FXN, TGFB2, ACTB, BTG1, ITGB8, ESR1, REL, SMAD3 TPM3, CD40, SOX2, IGF1R, VEGFA, MUC1 |
| Pathway Category | Gene Count | miRNAs | p-Value |
|---|---|---|---|
| Nervous System | |||
| Glioma | 25 | 4 | 0.00000 |
| TGF-beta signaling pathway | 27 | 4 | 0.00005 |
| mTOR signaling pathway | 25 | 4 | 0.00015 |
| ErbB signaling pathway | 28 | 4 | 0.00015 |
| FoxO signaling pathway | 41 | 4 | 0.00101 |
| PI3K-Akt signaling pathway | 79 | 4 | 0.00613 |
| Hedgehog signaling pathway | 18 | 4 | 0.01366 |
| Axon guidance | 30 | 4 | 0.03526 |
| Cancer | |||
| Proteoglycans in cancer | 73 | 4 | 0.00000 |
| Pathways in cancer | 102 | 4 | 0.00000 |
| Transcriptional misregulation in cancer | 39 | 4 | 0.00101 |
| Cellular community | |||
| Signaling pathways regulating pluripotency of stem cells | 53 | 4 | 0.00000 |
| Focal adhesion | 66 | 4 | 0.00000 |
| Adherens junction | 24 | 4 | 0.00011 |
| Cell growth and death | |||
| Hippo signaling pathway | 52 | 4 | 0.00000 |
| Wnt signaling pathway | 40 | 4 | 0.00011 |
| p53 signaling pathway | 25 | 4 | 0.00048 |
| Cell cycle | 37 | 4 | 0.00311 |
| MAPK signaling pathway | 66 | 4 | 0.00483 |
| Rap1 signaling pathway | 51 | 4 | 0.01187 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-D.; Pan, H.-Y.; Huang, J.-B.; Liu, X.-P.; Li, J.-H.; Ho, C.-J.; Tsai, M.-H.; Yang, J.-L.; Chen, S.-F.; Chen, N.-C.; et al. Circulating MicroRNAs from Serum Exosomes May Serve as a Putative Biomarker in the Diagnosis and Treatment of Patients with Focal Cortical Dysplasia. Cells 2020, 9, 1867. https://doi.org/10.3390/cells9081867
Chen S-D, Pan H-Y, Huang J-B, Liu X-P, Li J-H, Ho C-J, Tsai M-H, Yang J-L, Chen S-F, Chen N-C, et al. Circulating MicroRNAs from Serum Exosomes May Serve as a Putative Biomarker in the Diagnosis and Treatment of Patients with Focal Cortical Dysplasia. Cells. 2020; 9(8):1867. https://doi.org/10.3390/cells9081867
Chicago/Turabian StyleChen, Shang-Der, Hsiu-Yung Pan, Jyun-Bin Huang, Xuan-Ping Liu, Jie-Hau Li, Chen-Jui Ho, Meng-Han Tsai, Jenq-Lin Yang, Shu-Fang Chen, Nai-Ching Chen, and et al. 2020. "Circulating MicroRNAs from Serum Exosomes May Serve as a Putative Biomarker in the Diagnosis and Treatment of Patients with Focal Cortical Dysplasia" Cells 9, no. 8: 1867. https://doi.org/10.3390/cells9081867
APA StyleChen, S. -D., Pan, H. -Y., Huang, J. -B., Liu, X. -P., Li, J. -H., Ho, C. -J., Tsai, M. -H., Yang, J. -L., Chen, S. -F., Chen, N. -C., & Chuang, Y. -C. (2020). Circulating MicroRNAs from Serum Exosomes May Serve as a Putative Biomarker in the Diagnosis and Treatment of Patients with Focal Cortical Dysplasia. Cells, 9(8), 1867. https://doi.org/10.3390/cells9081867