RNA-Binding Protein Rbm24 as a Multifaceted Post-Transcriptional Regulator of Embryonic Lineage Differentiation and Cellular Homeostasis



Abstract
:1. Introduction
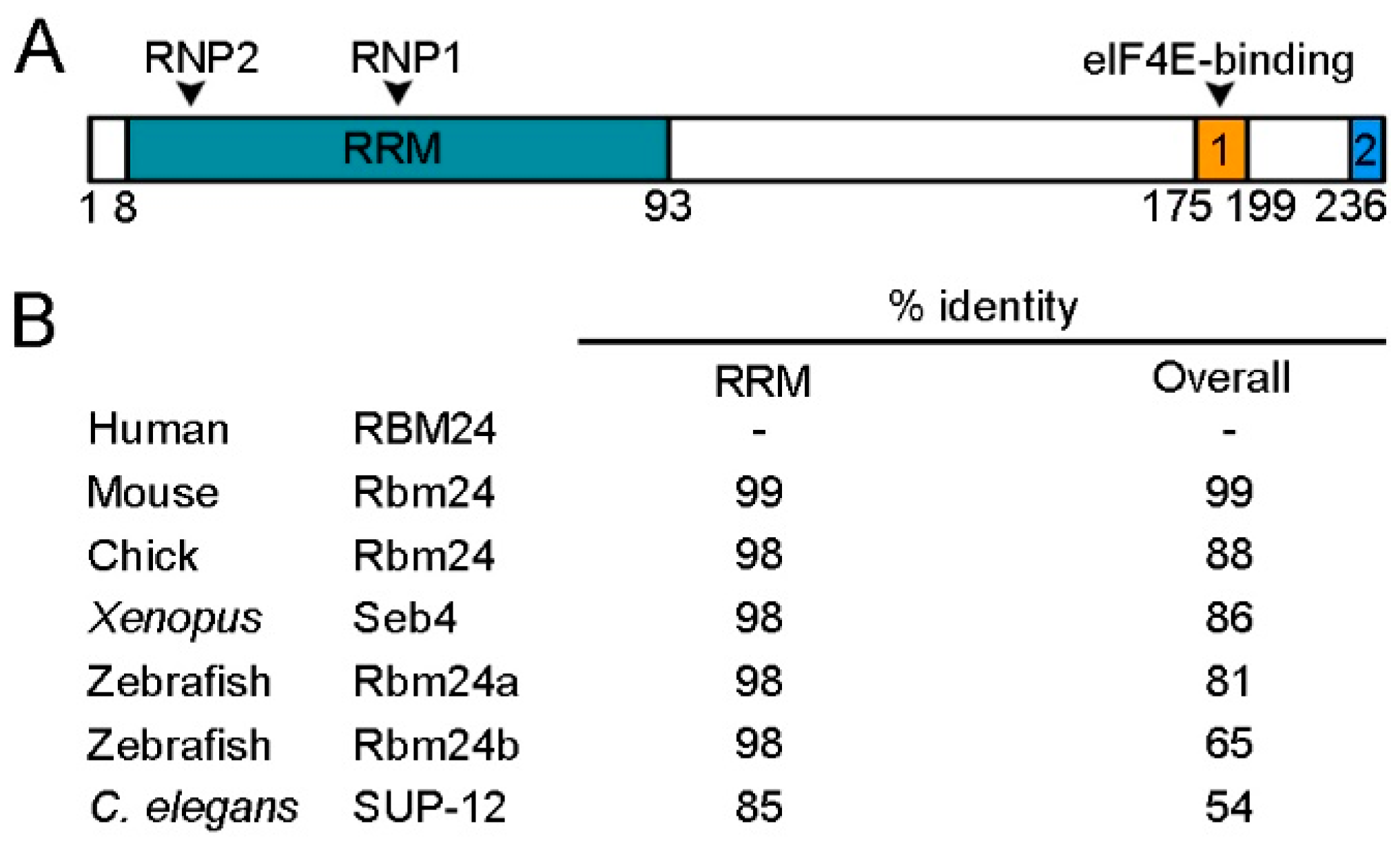
2. Rbm24 Functional Domains
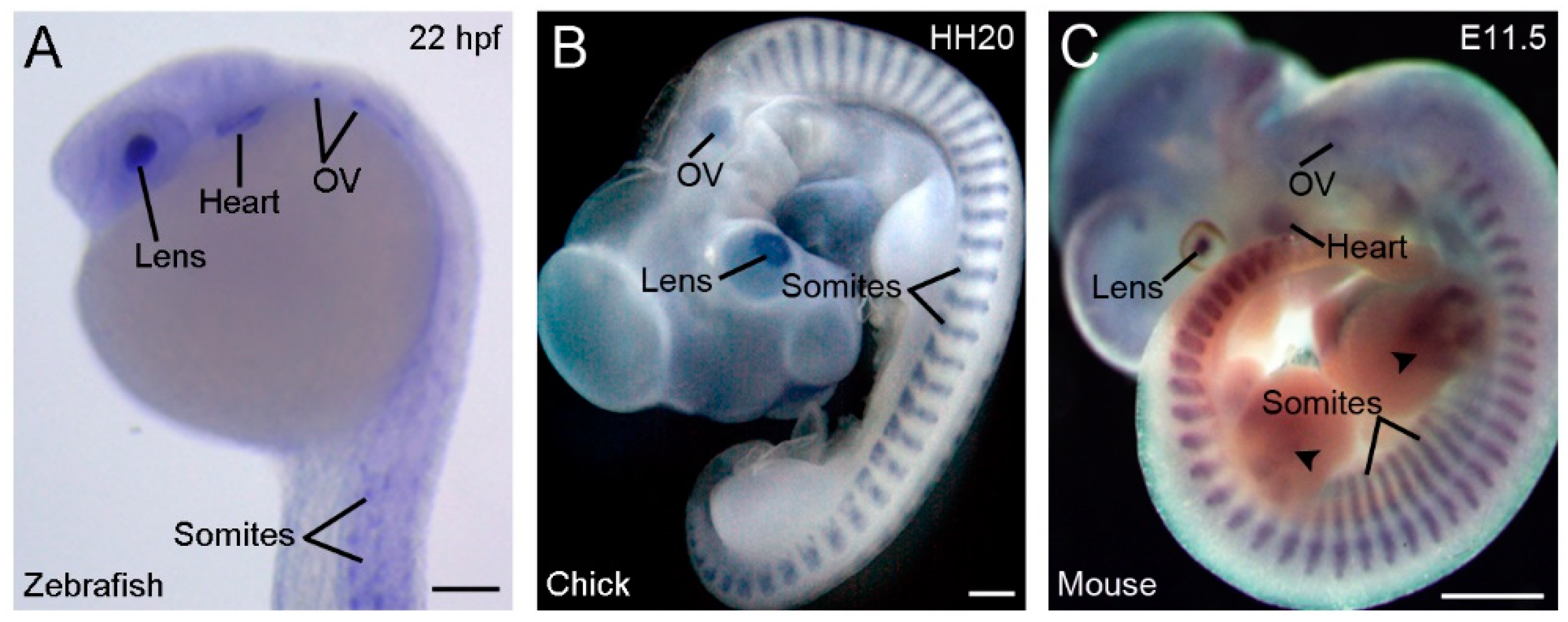
3. Rbm24 Displays Restricted Expression Patterns in Developing Vertebrate Embryos
4. Rbm24 Regulates Muscle Cell Development through Distinct Mechanisms
4.1. Rbm24 in Skeletal Myogenesis
4.2. Rbm24 is Required for Heart Development
4.3. Dynamic Subcellular Localization and Function of Rbm24
5. Rbm24 in Head Sensory Organ Development
5.1. Rbm24 Regulates Lens Fiber Cell Differentiation
5.2. Rbm24 and Rbm38 in Retinal Differentiation
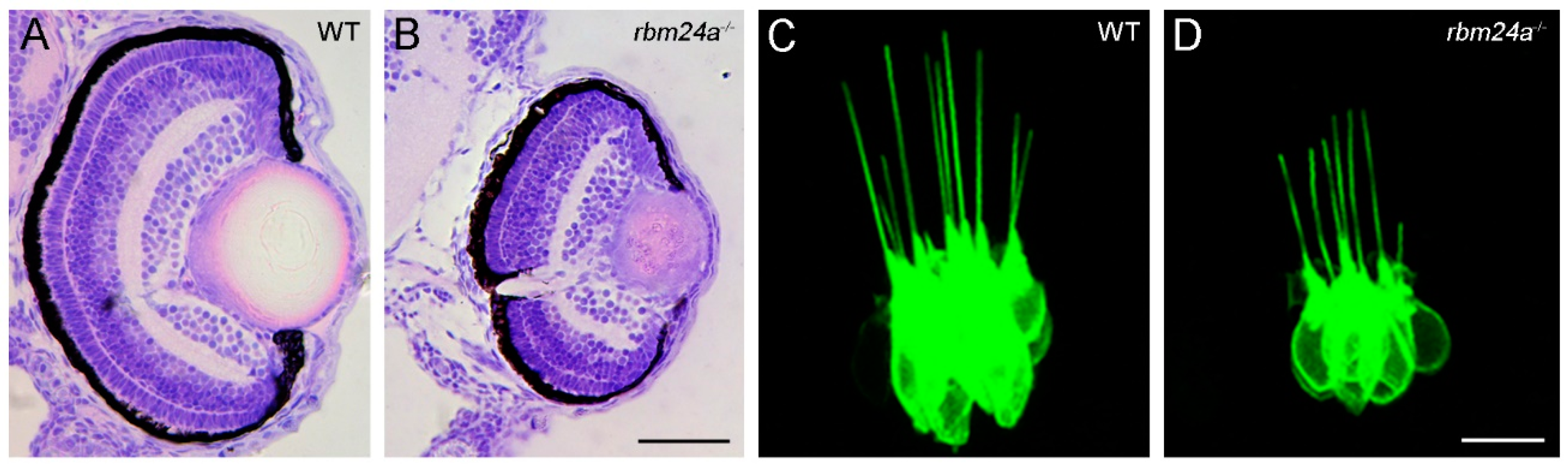
5.3. Rbm24 Is Required for Inner Ear Hair Cell Development
5.4. Rbm24 and Differentiation of Olfactory Sensory Neurons
6. Rbm24 in Embryonic Germ Layer Formation
7. Potential Implication of Rbm24 in Disease
7.1. A possible Tumor Suppressor Activity of Rbm24 and Rbm38
7.2. Interaction Between Rbm24 and microRNAs to Maintain Cellular Homeostasis
7.3. Rbm24 in Congenital Disorders and Infection Diseases
8. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Glisovic, T.; Bachorik, J.L.; Yong, J.; Dreyfuss, G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 2008, 582, 1977–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, A.R.; Mukherjee, N.; Keene, J.D. Systematic analysis of posttranscriptional gene expression. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 162–180. [Google Scholar] [CrossRef] [PubMed]
- Lukong, K.E.; Chang, K.-W.; Khandjian, E.W.; Richard, S. RNA-binding proteins in human genetic disease. Trends Genet. 2008, 24, 416–425. [Google Scholar] [CrossRef]
- Castello, A.; Fischer, B.; Hentze, M.W.; Preiss, T.; Palomares, A.C. RNA-binding proteins in Mendelian disease. Trends Genet. 2013, 29, 318–327. [Google Scholar] [CrossRef]
- Brinegar, A.E.; Cooper, T.A. Roles for RNA-binding proteins in development and disease. Brain Res. 2016, 1647, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Conlon, E.G.; Manley, J.L. RNA-binding proteins in neurodegeneration: Mechanisms in aggregate. Genes Dev. 2017, 31, 1509–1528. [Google Scholar] [CrossRef]
- Pereira, B.; Billaud, M.; Almeida, R. RNA-Binding Proteins in Cancer: Old Players and New Actors. Trends Cancer 2017, 3, 506–528. [Google Scholar] [CrossRef]
- Hong, S. RNA Binding Protein as an Emerging Therapeutic Target for Cancer Prevention and Treatment. J. Cancer Prev. 2017, 22, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Shotwell, C.R.; Cleary, J.D.; Berglund, J.A. The potential of engineered eukaryotic RNA-binding proteins as molecular tools and therapeutics. Wiley Interdiscip. Rev. RNA 2019, 11, e1573. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.W.; Preet, R.; Dhir, T.; Dixon, D.A.; Brody, J.R. Understanding and targeting the disease-related RNA binding protein human antigen R (HuR). Wiley Interdiscip. Rev. RNA 2020, 11, 1581. [Google Scholar] [CrossRef]
- Fetka, I.; Radeghieri, A.; Bouwmeester, T. Expression of the RNA recognition motif-containing protein SEB-4 during Xenopus embryonic development. Mech. Dev. 2000, 94, 283–286. [Google Scholar] [CrossRef]
- Poon, K.L.; Tan, K.T.; Wei, Y.Y.; Ng, C.P.; Colman, A.; Korzh, V.; Xu, X.Q. RNA-binding protein RBM24 is required for sarcomere assembly and heart contractility. Cardiovasc. Res. 2012, 94, 418–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Hu, Y.; Xu, H.; He, H.; Han, C.; Liu, H.; Wang, J.; Li, L. Characterization of the duck (Anas platyrhynchos) Rbm24 and Rbm38 genes and their expression profiles in myoblast and skeletal muscle tissues. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2016, 198, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Grifone, R.; Xie, X.; Bourgeois, A.; Saquet, A.; Duprez, D.; Shi, D.-L. The RNA-binding protein Rbm24 is transiently expressed in myoblasts and is required for myogenic differentiation during vertebrate development. Mech. Dev. 2014, 134, 1–15. [Google Scholar] [CrossRef]
- Yang, J.; Hung, L.-H.; Licht, T.; Kostin, S.; Looso, M.; Khrameeva, E.E.; Bindereif, A.; Schneider, A.; Braun, T. RBM24 Is a Major Regulator of Muscle-Specific Alternative Splicing. Dev. Cell 2014, 31, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Weeland, C.; Hoogenhof, M.V.D.; Beqqali, A.; E Creemers, E. Insights into alternative splicing of sarcomeric genes in the heart. J. Mol. Cell. Cardiol. 2015, 81, 107–113. [Google Scholar] [CrossRef]
- Liu, J.; Kong, X.; Zhang, M.; Yang, X.; Xu, X.Q. RNA binding protein 24 deletion disrupts global alternative splicing and causes dilated cardiomyopathy. Protein Cell 2018, 10, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Shao, M.; Lu, T.; Zhang, C.; Zhang, Y.-Z.; Kong, S.-H.; Shi, D.-L. Rbm24 controls poly(A) tail length and translation efficiency of crystallin mRNAs in the lens via cytoplasmic polyadenylation. Proc. Natl. Acad. Sci. USA 2020, 117, 7245–7254. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.; Kazan, H.; Chan, E.T.; Peña-Castillo, L.; Chaudhry, S.; Talukder, S.; Blencowe, B.J.; Morris, Q.; Hughes, T.R. Rapid and systematic analysis of the RNA recognition specificities of RNA-binding proteins. Nat. Biotechnol. 2009, 27, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Lucchesi, C.; Zhang, J.; Chen, X. Modulation of the p53 family network by RNA-binding proteins. Transl. Cancer Res. 2016, 5, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afroz, T.; Cienikova, Z.; Cléry, A.; Allain, F.H.-T. One, Two, Three, Four! How Multiple RRMs Read the Genome Sequence. Methods Enzymol. 2015, 558, 235–278. [Google Scholar] [CrossRef]
- Qian, K.; Li, M.; Wang, J.; Zhang, M.; Wang, M. Structural basis for mRNA recognition by human RBM38. Biochem. J. 2020, 477, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Boy, S.; Souopgui, J.; Amato, M.A.; Wegnez, M.; Pieler, T.; Perron, M. XSEB4R, a novel RNA-binding protein involved in retinal cell differentiation downstream of bHLH proneural genes. Development 2004, 131, 851–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Cho, S.-J.; Shu, L.; Yan, W.; Guerrero, T.; Kent, M.S.; Skorupski, K.; Chen, H.; Chen, X. Translational repression of p53 by RNPC1, a p53 target overexpressed in lymphomas. Genes Dev. 2011, 25, 1528–1543. [Google Scholar] [CrossRef] [Green Version]
- Lucchesi, C.A.; Zhang, J.; Ma, B.; Chen, M.; Chen, X. Disruption of the Rbm38-eIF4E Complex with a Synthetic Peptide Pep8 Increases p53 Expression. Cancer Res. 2018, 79, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhang, J.; Chen, X.; Cho, S.-J.; Chen, X.; Chen, X. Glycogen synthase kinase 3 promotes p53 mRNA translation via phosphorylation of RNPC1. Genes Dev. 2013, 27, 2246–2258. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Kong, X.; Lee, Y.M.; Zhang, M.K.; Guo, L.Y.; Lin, Y.; Lim, T.K.; Lin, Q.; Xu, X.Q.; Mun, L.Y.; et al. Stk38 Modulates Rbm24 Protein Stability to Regulate Sarcomere Assembly in Cardiomyocytes. Sci. Rep. 2017, 7, 44870. [Google Scholar] [CrossRef]
- Li, H.-Y.; Bourdelas, A.; Carron, C.; Shi, D.-L. The RNA-binding protein Seb4/RBM24 is a direct target of MyoD and is required for myogenesis during Xenopus early development. Mech. Dev. 2010, 127, 281–291. [Google Scholar] [CrossRef]
- Miller, R.A.; Christoforou, N.; Pevsner, J.; McCallion, A.S.; Gearhart, J.D. Efficient Array-Based Identification of Novel Cardiac Genes through Differentiation of Mouse ESCs. PLoS ONE 2008, 3, e2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.Q.; Soo, S.Y.; Sun, W.; Zweigerdt, R. Global Expression Profile of Highly Enriched Cardiomyocytes Derived from Human Embryonic Stem Cells. Stem Cells 2009, 27, 2163–2174. [Google Scholar] [CrossRef]
- Maragh, S.; A Miller, R.; Bessling, S.L.; McGaughey, D.M.; Wessels, M.W.; De Graaf, B.; Stone, E.A.; Bertoli-Avella, A.M.; Gearhart, J.D.; Fisher, S.; et al. Identification of RNA binding motif proteins essential for cardiovascular development. BMC Dev. Biol. 2011, 11, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maragh, S.; Miller, R.A.; Bessling, S.L.; Wang, G.; Hook, P.W.; McCallion, A.S. Rbm24a and Rbm24b Are Required for Normal Somitogenesis. PLoS ONE 2014, 9, e105460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, T.; Jen, H.-I.; Kang, H.; Klisch, T.J.; Zoghbi, H.Y.; Groves, A.K. Characterization of the Transcriptome of Nascent Hair Cells and Identification of Direct Targets of the Atoh1 Transcription Factor. J. Neurosci. 2015, 35, 5870–5883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grifone, R.; Saquet, A.; Xu, Z.; Shi, D.-L. Expression Patterns of Rbm24 in Lens, Nasal Epithelium, and Inner Ear During Mouse Embryonic Development. Dev. Dyn. 2018, 247, 1160–1169. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.; Hidaka, K.; Shirai, M.; Morisaki, T. RNA-binding motif protein 24 regulates myogenin expression and promotes myogenic differentiation. Genes Cells 2010, 15, 1158–1167. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.; Xu, E.; Mohibi, S.; De Anda, D.M.; Jiang, Y.; Zhang, J.; Chen, X. Rbm24, a target of p53, is necessary for proper expression of p53 and heart development. Cell Death Differ. 2018, 25, 1118–1130. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Brastrom, L.K.; Patel, S.D.; Scott, C.A.; Slusarski, D.C.; Lachke, S.A. The master transcription factor SOX2, mutated in anophthalmia/microphthalmia, is post-transcriptionally regulated by the conserved RNA-binding protein RBM24 in vertebrate eye development. Hum. Mol. Genet. 2019, 29, 591–604. [Google Scholar] [CrossRef]
- Cheng, X.N.; Zhang, J.J.; Shi, D.L. Loss of Rbm24a causes defective hair cell development in the zebrafish inner ear and neuromasts. J. Genet. Genomics 2020. [Google Scholar] [CrossRef]
- Wei, P.; Yang, J.; Zhang, D.; Cui, M.; Li, L. lncRNA HAND2-AS1 Regulates Prostate Cancer Cell Growth Through Targeting the miR-106a-5p/RBM24 Axis. OncoTargets Ther. 2020, 13, 4523–4531. [Google Scholar] [CrossRef] [PubMed]
- Hua, W.-F.; Zhong, Q.; Xia, T.-L.; Chen, Q.; Zhang, M.-Y.; Zhou, A.-J.; Tu, Z.-W.; Qu, C.; Li, M.-Z.; Xia, Y.-F.; et al. RBM24 suppresses cancer progression by upregulating miR-25 to target MALAT1 in nasopharyngeal carcinoma. Cell Death Dis. 2016, 7, e2352. [Google Scholar] [CrossRef] [Green Version]
- Du, C.; Shen, Z.; Zang, R.; Xie, H.; Li, H.; Chen, P.; Hang, B.; Xu, X.; Tang, W.; Xia, Y. Negative feedback circuitry between MIR143HG and RBM24 in Hirschsprung disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2016, 1862, 2127–2136. [Google Scholar] [CrossRef] [PubMed]
- Ohe, K.; Yoshida, M.; Nakano-Kobayashi, A.; Hosokawa, M.; Sako, Y.; Sakuma, M.; Okuno, Y.; Usui, T.; Ninomiya, K.; Nojima, T.; et al. RBM24 promotes U1 snRNP recognition of the mutated 5′ splice site in the IKBKAP gene of familial dysautonomia. RNA 2017, 23, 1393–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.; Zhao, K.; Yao, Y.; Guo, J.; Gao, X.; Yang, Q.; Guo, M.; Zhu, W.; Wang, Y.; Wu, C.; et al. RNA binding protein 24 regulates the translation and replication of hepatitis C virus. Protein Cell 2018, 9, 930–944. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Yang, B.; Cao, H.; Zhao, K.; Yuan, Y.; Chen, Y.; Zhang, Z.; Wang, Y.; Pei, R.; Chen, J.; et al. RBM24 stabilizes hepatitis B virus pregenomic RNA but inhibits core protein translation by targeting the terminal redundancy sequence. Emerg. Microbes Infect. 2018, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Yang, B.; Chen, Y.; Wang, H.; Hu, X.; Zhou, Y.; Gao, X.; Lu, M.; Niu, J.; Wen, Z.; et al. RNA-Binding Motif Protein 24 (RBM24) Is Involved in Pregenomic RNA Packaging by Mediating Interaction between Hepatitis B Virus Polymerase and the Epsilon Element. J. Virol. 2019, 93, 02161-18. [Google Scholar] [CrossRef] [Green Version]
- Green, Y.S.; Vetter, M.L. EBF proteins participate in transcriptional regulation of Xenopus muscle development. Dev. Biol. 2011, 358, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Anyanful, A.; Ono, K.; Johnsen, R.C.; Ly, H.; Jensen, V.L.; Baillie, D.L.; Ono, S. The RNA-binding protein SUP-12 controls muscle-specific splicing of the ADF/cofilin pre-mRNA in C. elegans. J. Cell Biol. 2004, 167, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Kuroyanagi, H.; Ohno, G.; Mitani, S.; Hagiwara, M. The Fox-1 Family and SUP-12 Coordinately Regulate Tissue-Specific Alternative Splicing In Vivo. Mol. Cell. Biol. 2007, 27, 8612–8621. [Google Scholar] [CrossRef] [Green Version]
- Ohno, G.; Ono, K.; Togo, M.; Watanabe, Y.; Ono, S.; Hagiwara, M.; Kuroyanagi, H. Muscle-Specific Splicing Factors ASD-2 and SUP-12 Cooperatively Switch Alternative Pre-mRNA Processing Patterns of the ADF/Cofilin Gene in Caenorhabditis elegans. PLoS Genet. 2012, 8, e1002991. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, M. Myogenic progenitor cells and skeletal myogenesis in vertebrates. Curr. Opin. Genet. Dev. 2006, 16, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Nikonova, E.; Kao, S.-Y.; Ravichandran, K.; Wittner, A.; Spletter, M.L. Conserved functions of RNA-binding proteins in muscle. Int. J. Biochem. Cell Biol. 2019, 110, 29–49. [Google Scholar] [CrossRef] [PubMed]
- Llorian, M.; Smith, C.W. Decoding muscle alternative splicing. Curr. Opin. Genet. Dev. 2011, 21, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Hidaka, K.; Jin, D.; Morisaki, T. RNA-binding proteins Rbm38 and Rbm24 regulate myogenic differentiation via p21-dependent and -independent regulatory pathways. Genes Cells 2009, 2009 14, 1241–1252. [Google Scholar] [CrossRef]
- de Groot, N.E.; van den Hoogenhof, M.M.G.; Najafi, A.; van der Made, I.; van der Velden, J.; Beqqali, A.; Pinto, Y.M.; Creemers, E.E. Heterozygous loss of Rbm24 in the adult mouse heart increases sarcomere slack length but does not affect function. Sci. Rep. 2020, 10, 7687. [Google Scholar] [CrossRef]
- Guo, W.; Schäfer, S.; Greaser, M.L.; Radke, M.H.; Liss, M.; Govindarajan, T.; Maatz, H.; Schulz, H.; Li, S.; Parrish, A.M.; et al. RBM20, a gene for hereditary cardiomyopathy, regulates titin splicing. Nat. Med. 2012, 18, 766–773. [Google Scholar] [CrossRef]
- Fochi, S.; Lorenzi, P.; Galasso, M.; Stefani, C.; Trabetti, E.; Zipeto, D.; Romanelli, M.G. The Emerging Role of the RBM20 and PTBP1 Ribonucleoproteins in Heart Development and Cardiovascular Diseases. Genes 2020, 11, 402. [Google Scholar] [CrossRef] [Green Version]
- Brauch, K.M.; Karst, M.L.; Herron, K.J.; De Andrade, M.; Pellikka, P.A.; Rodeheffer, R.J.; Michels, V.V.; Olson, T.M. Mutations in Ribonucleic Acid Binding Protein Gene Cause Familial Dilated Cardiomyopathy. J. Am. Coll. Cardiol. 2009, 54, 930–941. [Google Scholar] [CrossRef] [Green Version]
- Ware, J.S.; Cook, S.A. Role of titin in cardiomyopathy: From DNA variants to patient stratification. Nat. Rev. Cardiol. 2017, 15, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Hey, T.M.; Rasmussen, T.B.; Madsen, T.; Aagaard, M.M.; Harbo, M.; Mølgaard, H.; Moller, J.E.; Eiskjær, H.; Mogensen, J. Pathogenic RBM20-Variants Are Associated With a Severe Disease Expression in Male Patients With Dilated Cardiomyopathy. Circ. Hear. Fail. 2019, 12, e005700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaertner, A.; Brodehl, A.; Milting, H. Screening for mutations in human cardiomyopathy- is RBM24 a new but rare disease gene? Protein Cell 2018, 10, 393–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, J.; Iijima, M.; Yoshimoto, N.; Niimi, T.; Kuroda, S.; Maturana, A.D. RBM20 and RBM24 cooperatively promote the expression of short enh splice variants. FEBS Lett. 2016, 590, 2262–2274. [Google Scholar] [CrossRef]
- Kuwasako, K.; Takahashi, M.; Unzai, S.; Tsuda, K.; Yoshikawa, S.; He, F.; Kobayashi, N.; Güntert, P.; Shirouzu, M.; Ito, T.; et al. RBFOX and SUP-12 sandwich a G base to cooperatively regulate tissue-specific splicing. Nat. Struct. Mol. Biol. 2014, 21, 778–786. [Google Scholar] [CrossRef]
- Morita, H.; Komuro, I. Heart Failure as an Aging-Related Phenotype. Int. Hear. J. 2018, 59, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Tan, K.T.; Liu, J.; Kong, X.; Huang, Z.; Xu, X.Q. Global profiling of Rbm24 bound RNAs uncovers a multi-tasking RNA binding protein. Int. J. Biochem. Cell Biol. 2018, 94, 10–21. [Google Scholar] [CrossRef]
- Ehler, E. RBM24: A ’regional business manager’ in charge of the maintenance of sarcomeric protein expression 24 h a day? Cardiovasc. Res. 2012, 94, 393–394. [Google Scholar] [CrossRef] [Green Version]
- Relaix, F.; Rocancourt, D.; Mansouri, A.; Buckingham, M. A Pax3/Pax7-dependent population of skeletal muscle progenitor cells. Nature 2005, 435, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Lin, Y.; Liu, J.; Zhang, Z.G.; Fu, W.; Guo, L.Y.; Pan, L.; Kong, X.; Zhang, M.K.; Lu, Y.H.; et al. Rbm24 Regulates Alternative Splicing Switch in Embryonic Stem Cell Cardiac Lineage Differentiation. Stem Cells 2016, 34, 1776–1789. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Miller, J.W.; Mankodi, A.; Kanadia, R.N.; Yuan, Y.; Moxley, R.T.; Swanson, M.; Thornton, C.A. Failure of MBNL1-dependent post-natal splicing transitions in myotonic dystrophy. Hum. Mol. Genet. 2006, 15, 2087–2097. [Google Scholar] [CrossRef] [PubMed]
- Brastrom, L.K.; Scott, C.A.; Dawson, D.V.; Slusarski, D.C. A High-Throughput Assay for Congenital and Age-Related Eye Diseases in Zebrafish. Biomedicines 2019, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dash, S.; Siddam, A.D.; Barnum, C.E.; Janga, S.C.; Lachke, S.A. RNA-binding proteins in eye development and disease: Implication of conserved RNA granule components. Wiley Interdiscip. Rev. RNA 2016, 7, 527–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachke, S.A.; Alkuraya, F.S.; Kneeland, S.C.; Ohn, T.; Aboukhalil, A.; Howell, G.R.; Saadi, I.; Cavallesco, R.; Yue, Y.; Tsai, A.C.-H.; et al. Mutations in the RNA Granule Component TDRD7 Cause Cataract and Glaucoma. Science 2011, 331, 1571–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvekl, A.; Ashery-Padan, R. The cellular and molecular mechanisms of vertebrate lens development. Development 2014, 141, 4432–4447. [Google Scholar] [CrossRef] [Green Version]
- Greiling, T.M.S.; Aose, M.; Clark, J.I. Cell fate and differentiation of the developing ocular lens. Investig. Opthalmology Vis. Sci. 2009, 51, 1540–1546. [Google Scholar] [CrossRef] [Green Version]
- Eckmann, C.R.; Rammelt, C.; Wahle, E. Control of poly(A) tail length. Wiley Interdiscip. Rev. RNA 2010, 2, 348–361. [Google Scholar] [CrossRef]
- Goss, D.J.; Kleiman, F.E. Poly(A) binding proteins: Are they all created equal? Wiley Interdiscip. Rev. RNA 2012, 4, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, A.; Pasquinelli, A.E. Tales of Detailed Poly(A) Tails. Trends Cell Biol. 2019, 29, 191–200. [Google Scholar] [CrossRef]
- Charlesworth, A.; Meijer, H.A.; De Moor, C.H. Specificity factors in cytoplasmic polyadenylation. Wiley Interdiscip. Rev. RNA 2013, 4, 437–461. [Google Scholar] [CrossRef]
- Reyes, J.M.; Ross, P.J. Cytoplasmic polyadenylation in mammalian oocyte maturation. Wiley Interdiscip. Rev. RNA 2015, 7, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Barnum, C.E.; Al Saai, S.; Patel, S.D.; Cheng, C.; Anand, D.; Xu, X.; Dash, S.; Siddam, A.D.; Glazewski, L.; Paglione, E.; et al. The Tudor-domain protein TDRD7, mutated in congenital cataract, controls the heat shock protein HSPB1 (HSP27) and lens fiber cell morphology. Hum. Mol. Genet. 2020, 29, 2076–2097. [Google Scholar] [CrossRef]
- Mishima, Y.; Fukao, A.; Kishimoto, T.; Sakamoto, H.; Fujiwara, T.; Inoue, K. Translational inhibition by deadenylation-independent mechanisms is central to microRNA-mediated silencing in zebrafish. Proc. Natl. Acad. Sci. USA 2012, 109, 1104–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco-Sánchez, B.; Clément, A.; Phillips, J.; Westerfield, M. Zebrafish models of human eye and inner ear diseases. Methods Cell Biol. 2017, 138, 415–467. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, T. The genetics of hair-cell function in zebrafish. J. Neurogenet. 2017, 31, 102–112. [Google Scholar] [CrossRef]
- Lush, M.E.; Diaz, D.C.; Koenecke, N.; Baek, S.; Boldt, H.; Peter, M.K.S.; Gaitan-Escudero, T.; Romero-Carvajal, A.; Busch-Nentwich, E.M.; Perera, A.G.; et al. scRNA-Seq reveals distinct stem cell populations that drive hair cell regeneration after loss of Fgf and Notch signaling. eLife 2019, 8, 44431. [Google Scholar] [CrossRef]
- Chatterjee, P.; Padmanarayana, M.; Abdullah, N.; Holman, C.L.; Ladu, J.; Tanguay, R.L.; Johnson, C.P. Otoferlin Deficiency in Zebrafish Results in Defects in Balance and Hearing: Rescue of the Balance and Hearing Phenotype with Full-Length and Truncated Forms of Mouse Otoferlin. Mol. Cell. Biol. 2015, 35, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- DeSmidt, A.A.; Zou, B.; Grati, M.; Yan, D.; Mittal, R.; Yao, Q.; Richmond, M.T.; Denyer, S.; Liu, X.Z.; Lu, Z. Zebrafish Model for Nonsyndromic X-Linked Sensorineural Deafness, DFNX1. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2019, 303, 544–555. [Google Scholar] [CrossRef]
- Sokpor, G.; Abbas, E.; Rosenbusch, J.; Staiger, J.F.; Tuoc, T. Transcriptional and Epigenetic Control of Mammalian Olfactory Epithelium Development. Mol. Neurobiol. 2018, 55, 8306–8327. [Google Scholar] [CrossRef]
- Shum, E.Y.; Espinoza, J.L.; Ramaiah, M.; Wilkinson, M.F. Identification of novel post-transcriptional features in olfactory receptor family mRNAs. Nucleic Acids Res. 2015, 43, 9314–9326. [Google Scholar] [CrossRef] [Green Version]
- Souopgui, J.; Rust, B.; Vanhomwegen, J.; Heasman, J.; Henningfeld, K.A.; Bellefroid, E.; Pieler, T. The RNA-binding protein XSeb4R: A positive regulator of VegT mRNA stability and translation that is required for germ layer formation in Xenopus. Genes Dev. 2008, 22, 2347–2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carron, C.; Shi, D.-L. Specification of anteroposterior axis by combinatorial signaling during Xenopus development. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 150–168. [Google Scholar] [CrossRef] [PubMed]
- Bentaya, S.; Ghogomu, S.M.; Vanhomwegen, J.; Van Campenhout, C.; Thelie, A.; Dhainaut, M.; Bellefroid, E.J.; Souopgui, J. The RNA-binding protein XSeb4R regulates maternal Sox3 at the posttranscriptional level during maternal-zygotic transition in Xenopus. Dev. Biol. 2012, 363, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Despic, V.; Neugebauer, K.M. RNA tales – how embryos read and discard messages from mom. J. Cell Sci. 2018, 131, jcs201996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, M.C.; Dhellemmes, M.; Lambert, E.; Ray, P.F.; Arnoult, C. Diversity of RNA-Binding Proteins Modulating Post-Transcriptional Regulation of Protein Expression in the Maturing Mammalian Oocyte. Cells 2020, 9, 662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendez, R.; Richter, J.D. Translational control by CPEB: A means to the end. Nat. Rev. Mol. Cell Biol. 2001, 2, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Villalba, A.; Coll, O.; Gebauer, F. Cytoplasmic polyadenylation and translational control. Curr. Opin. Genet. Dev. 2011, 21, 452–457. [Google Scholar] [CrossRef]
- Winata, C.L.; Łapiński, M.; Pryszcz, L.P.; Vaz, C.; Bin Ismail, M.H.; Nama, S.; Hajan, H.S.; Lee, S.G.P.; Korzh, V.; Sampath, P.; et al. Cytoplasmic polyadenylation-mediated translational control of maternal mRNAs directs maternal-to-zygotic transition. Development 2017, 145, dev159566. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zhang, M.; Qian, Y.; Xu, E.; Zhang, J.; Chen, X. Rbm24, an RNA-binding Protein and a Target of p53, Regulates p21 Expression via mRNA Stability. J. Biol. Chem. 2013, 289, 3164–3175. [Google Scholar] [CrossRef] [Green Version]
- Shu, L.; Yan, W.; Chen, X. RNPC1, an RNA-binding protein and a target of the p53 family, is required for maintaining the stability of the basal and stress-induced p21 transcript. Genes Dev. 2006, 20, 2961–2972. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Cho, S.J.; Chen, X. RNPC1, an RNA-binding protein and a target of the p53 family, regulates p63 expression through mRNA stability. Proc. Natl. Acad. Sci. USA 2010, 107, 9614–9619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, E.; Zhang, J.; Zhang, M.; Jiang, Y.; Cho, S.-J.; Chen, X. RNA-binding protein RBM24 regulates p63 expression via mRNA stability. Mol. Cancer Res. 2013, 12, 359–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Feng, X.; Sun, W.; Zhang, J.; Chen, X. Serine 195 phosphorylation in the RNA-binding protein Rbm38 increases p63 expression by modulating Rbm38’s interaction with the Ago2–miR203 complex. J. Biol. Chem. 2018, 294, 2449–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amrane, S.; Rebora, K.; Zniber, I.; Dupuy, D.; Mackereth, C.D. Backbone-independent nucleic acid binding by splicing factor SUP-12 reveals key aspects of molecular recognition. Nat. Commun. 2014, 5, 4595. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, S.K.; Mackereth, C.D. 1H, 15N and 13C backbone and side chain resonance assignments of the RRM domain from human RBM24. Biomol. NMR Assign. 2016, 10, 237–240. [Google Scholar] [CrossRef]
- Cao, Q.; Anyansi, C.; Hu, X.; Xu, L.; Xiong, L.; Tang, W.; Mok, M.T.S.; Cheng, C.; Fan, X.; Gerstein, M.; et al. Reconstruction of enhancer–target networks in 935 samples of human primary cells, tissues and cell lines. Nat. Genet. 2017, 49, 1428–1436. [Google Scholar] [CrossRef]
- Ciafrè, S.A.; Galardi, S. microRNAs and RNA-binding proteins. RNA Biol. 2013, 10, 935–942. [Google Scholar] [CrossRef] [Green Version]
- Vacchi-Suzzi, C.; Hahne, F.; Scheubel, P.; Marcellin, M.; Dubost, V.; Westphal, M.; Boeglen, C.; Büchmann-Møller, S.; Cheung, M.S.; Cordier, A.; et al. Heart Structure-Specific Transcriptomic Atlas Reveals Conserved microRNA-mRNA Interactions. PLoS ONE 2013, 8, e52442. [Google Scholar] [CrossRef] [Green Version]
- Cardinali, B.; Cappella, M.; Provenzano, C.; Garcia-Manteiga, J.M.; Lazarevic, D.; Cittaro, D.; Martelli, F.; Falcone, G. MicroRNA-222 regulates muscle alternative splicing through Rbm24 during differentiation of skeletal muscle cells. Cell Death Dis. 2016, 7, e2086. [Google Scholar] [CrossRef]
- Slaugenhaupt, S.A.; Gusella, J.F. Familial dysautonomia. Curr. Opin. Genet. Dev. 2002, 12, 307–311. [Google Scholar] [CrossRef]
- van den Hoogenhof, M.M.G.; van Der Made, I.; de Groot, N.E.; Damanafshan, A.; van Amersfoorth, S.C.M.; Zentilin, L.; Giacca, M.; Pinto, Y.M.; Creemers, E.E. AAV9-mediated Rbm24 overexpression induces fibrosis in the mouse heart. Sci. Rep. 2018, 8, 11696. [Google Scholar] [CrossRef] [PubMed]
- Müller-McNicoll, M.; Rossbach, O.; Hui, J.; Medenbach, J. Auto-regulatory feedback by RNA-binding proteins. J. Mol. Cell Biol. 2019, 11, 930–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue or Disease | Post-Transcriptional Regulation | Role in Development or Disease |
|---|---|---|
| Skeletal muscle | Muscle-specific pre-mRNA splicing, mRNA stability | Myogenic differentiation, somitogenesis, and sarcomere organization [16,17,30,34,37] |
| Cardiac muscle | Muscle-specific pre-mRNA splicing, mRNA stability and translation | Heart development, sarcomere assembly, and cardiac contractility [14,17,19,33,38] |
| Lens | Cytoplasmic polyadenylation, mRNA stability | Lens fiber cell differentiation, and lens transparency [20,39] |
| Inner ear/Neuromasts | mRNA stability | Hair cell morphogenesis and differentiation [40] |
| Olfactory epithelium | Unknown (cytoplasmic localization in neuronal cells) | Unknown |
| Blastula/Gastrula | Unknown | Germ layer formation [30] |
| Prostate cancer | Interaction with miR-106a-5p | Inhibition of tumorigenesis [41] |
| Nasopharyngeal carcinoma | Interaction with miR-25 | Inhibition of cell proliferation, migration and invasion [42] |
| Hirschsprung disease | Interaction with MIR143HG | Proliferation or migration of enteric neural crest cells, and gangliogenesis [43] |
| Familial dysautonomia | Increased recognition of the mutated 5’ splicing site in IKBKAP gene by U1 snRNP | Possible protective role against the aberrant splicing of the mutated gene [44] |
| Hepatitis A and B | Pregenomic RNA packaging and replication | Possible host factor for the viruses [45,46,47] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grifone, R.; Shao, M.; Saquet, A.; Shi, D.-L. RNA-Binding Protein Rbm24 as a Multifaceted Post-Transcriptional Regulator of Embryonic Lineage Differentiation and Cellular Homeostasis. Cells 2020, 9, 1891. https://doi.org/10.3390/cells9081891
Grifone R, Shao M, Saquet A, Shi D-L. RNA-Binding Protein Rbm24 as a Multifaceted Post-Transcriptional Regulator of Embryonic Lineage Differentiation and Cellular Homeostasis. Cells. 2020; 9(8):1891. https://doi.org/10.3390/cells9081891
Chicago/Turabian StyleGrifone, Raphaëlle, Ming Shao, Audrey Saquet, and De-Li Shi. 2020. "RNA-Binding Protein Rbm24 as a Multifaceted Post-Transcriptional Regulator of Embryonic Lineage Differentiation and Cellular Homeostasis" Cells 9, no. 8: 1891. https://doi.org/10.3390/cells9081891
APA StyleGrifone, R., Shao, M., Saquet, A., & Shi, D. -L. (2020). RNA-Binding Protein Rbm24 as a Multifaceted Post-Transcriptional Regulator of Embryonic Lineage Differentiation and Cellular Homeostasis. Cells, 9(8), 1891. https://doi.org/10.3390/cells9081891