A Novel Approach for Detecting Unique Variations among Infectious Bacterial Species in Endocarditic Cardiac Valve Vegetation




Abstract
:1. Introduction
2. Material and Methods
2.1. Patients and Histological Analysis
2.2. Vegetation Preparation
2.3. Scanning Electron Microscopy and Energy-Dispersive X-ray Spectroscopy
2.4. Statistical Analysis
3. Results
3.1. Histological and Immunohistochemical Observations
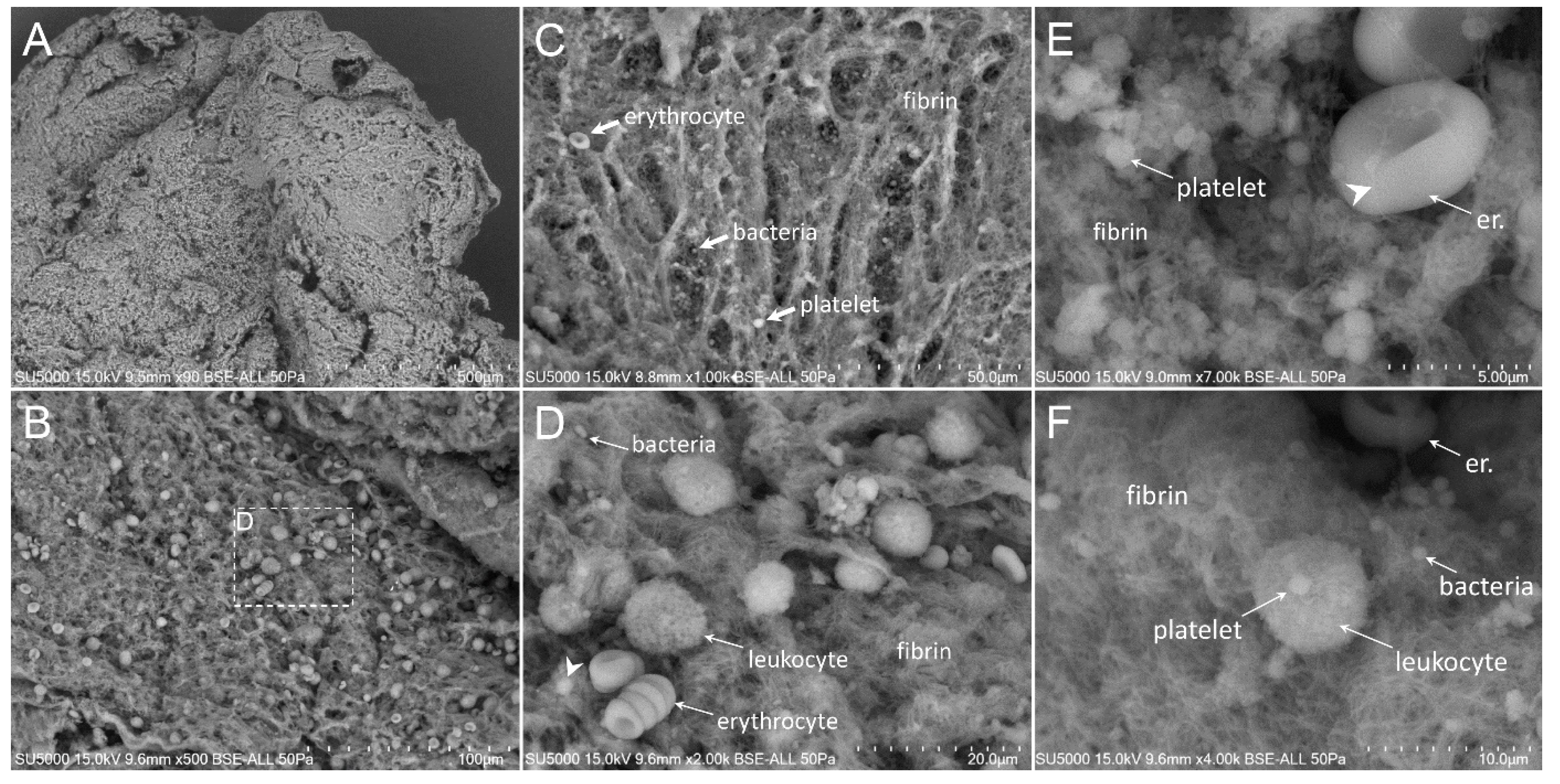
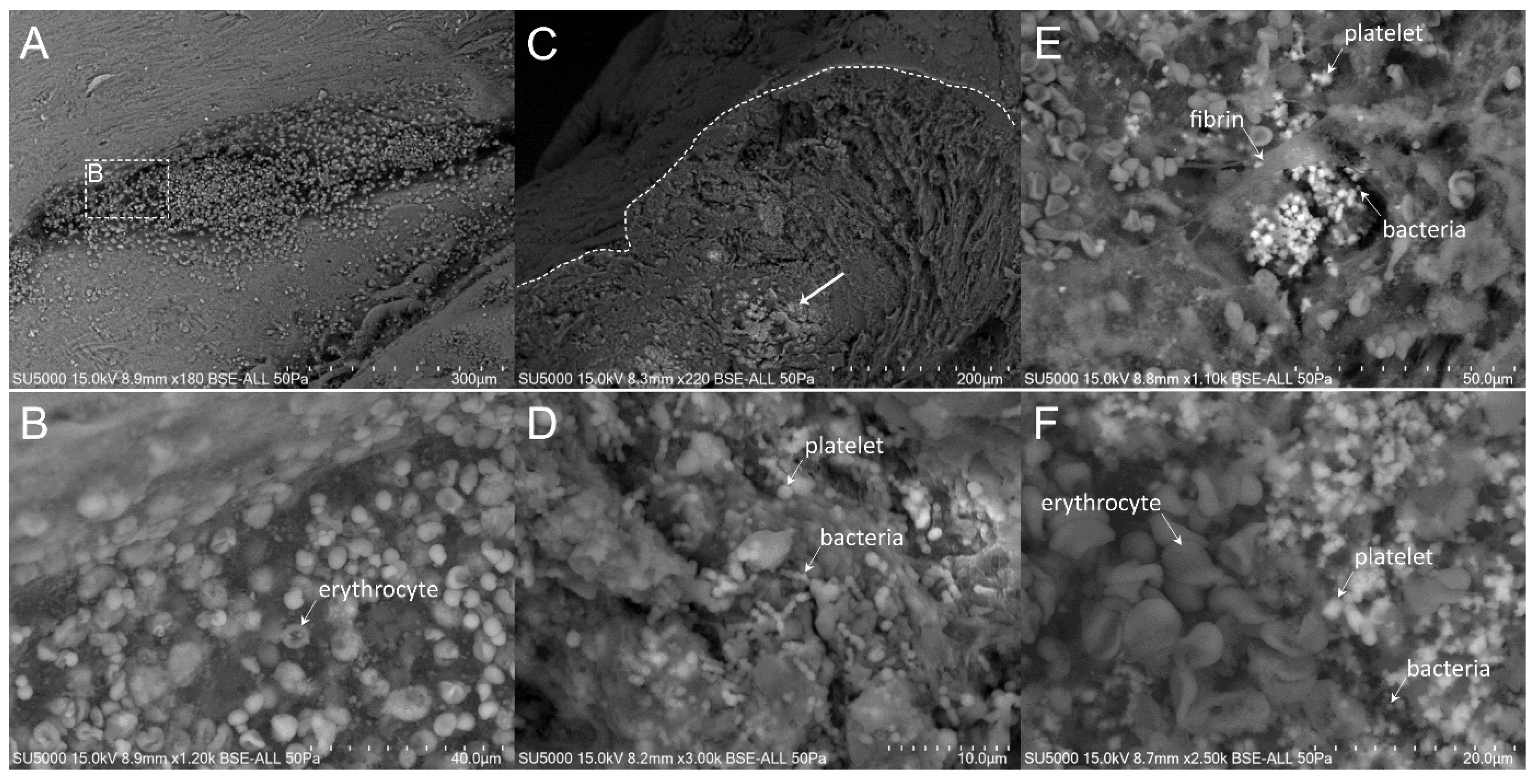
3.2. Scanning Electron Microscopy
3.3. Presence and Organization of Bacteria in the Vegetative Tissue
3.4. The Different Extent of Inflammation
3.5. Platelet Recruitment and Extent of the Fibrin Network
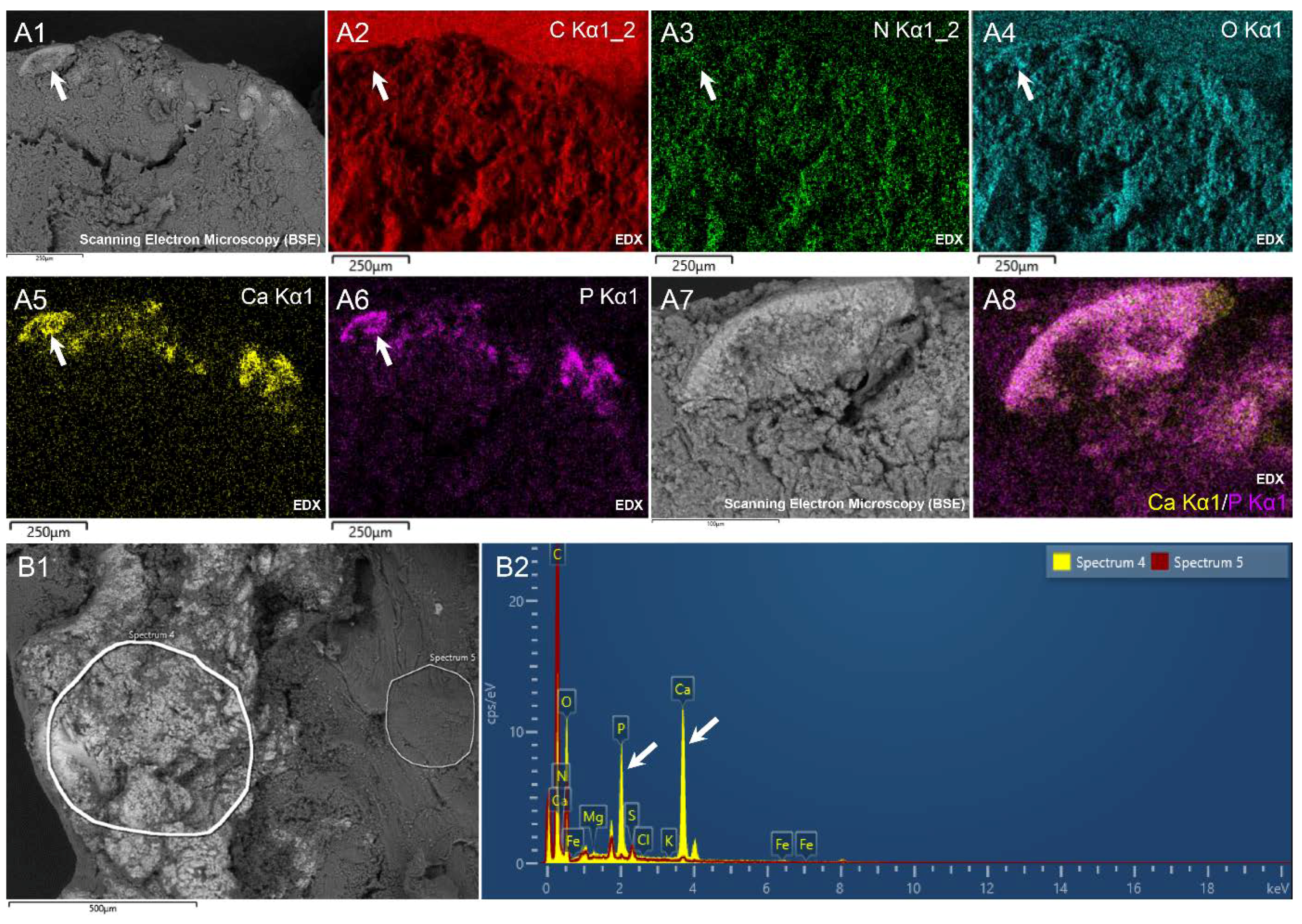
3.6. Energy Dispersive X-Ray Spectroscopy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Habib, G.; Lancellotti, P.; Erba, P.-A.; Sadeghpour, A.; Meshaal, M.; Sambola, A.; Furnaz, S.; Citro, R.; Ternacle, J.; Donal, E.; et al. The ESC-EORP EURO-ENDO (European Infective Endocarditis) registry. Eur. Hear. J. Qual. Care Clin. Outcomes 2019, 5, 202–207. [Google Scholar] [CrossRef]
- Klein, M.; Wang, A. Infective Endocarditis. J. Intensiv. Care Med. 2014, 31, 151–163. [Google Scholar] [CrossRef]
- Iung, B.; Duval, X. Infective endocarditis: Innovations in the management of an old disease. Nat. Rev. Cardiol. 2019, 16, 623–635. [Google Scholar] [CrossRef]
- Rasmussen, R.V. Anticoagulation in Patients with Stroke with Infective Endocarditis is Safe. Stroke 2011, 42, 1795–1796. [Google Scholar] [CrossRef] [Green Version]
- Tornos, P.; Almirante, B.; Mirabet, S.; Permanyer, G.; Pahissa, A.; Soler-Soler, J. Infective Endocarditis Due to Staphylococcus aureus. Arch. Intern. Med. 1999, 159, 473–475. [Google Scholar] [CrossRef] [Green Version]
- Anavekar, N.; Tleyjeh, I.M.; Mirzoyev, Z.; Steckelberg, J.M.; Haddad, C.; Khandaker, M.H.; Wilson, W.R.; Chandrasekaran, K.; Baddour, L.M. Impact of Prior Antiplatelet Therapy on Risk of Embolism in Infective Endocarditis. Clin. Infect. Dis. 2007, 44, 1180–1186. [Google Scholar] [CrossRef]
- Chan, K.-L.; Dumesnil, J.G.; Cujec, B.; Sanfilippo, A.J.; Jue, J.A.; Turek, M.I.; Robinson, T.; Moher, D. Investigators of the Multicenter Aspirin Study in Infective Endocarditis A Randomized Trial of Aspirin on the Risk of Embolic Events in Patients with Infective Endocarditis. J. Am. Coll. Cardiol. 2003, 42, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Arman, M.; Krauel, K.; Tilley, D.O.; Weber, C.; Cox, D.; Greinacher, A.; Kerrigan, S.W.; Watson, S.P. Amplification of bacteria-induced platelet activation is triggered by FcγRIIA, integrin αIIbβ3, and platelet factor 4. Blood 2014, 123, 3166–3174. [Google Scholar] [CrossRef]
- Hannachi, N.; Baudoin, J.-P.; Prasanth, A.; Habib, G.; Camoin-Jau, L. The distinct effects of aspirin on platelet aggregation induced by infectious bacteria. Platelets 2019, 1–11. [Google Scholar] [CrossRef]
- Lepidi, H.; Casalta, J.-P.; Gouriet, F.; Collart, F.; Habib, G.; Raoult, D. Infective endocarditis incidentally discovered by pathological examination. J. Clin. Pathol. 2007, 61, 233–234. [Google Scholar] [CrossRef]
- Hubers, S.A.; DeSimone, D.C.; Gersh, B.J.; Anavekar, N.S. Infective Endocarditis: A Contemporary Review. Mayo Clin. Proc. 2020, 95, 982–997. [Google Scholar] [CrossRef] [PubMed]
- Scimeca, M.; Bischetti, S.; Lamsira, H.K.; Bonfiglio, R.; Bonanno, E. Energy Dispersive X-ray (EDX) microanalysis: A powerful tool in biomedical research and diagnosis. Eur. J. Histochem. 2018, 62, 2841. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.I.; Oh, S.-W.; Kim, Y.-J. Power of Scanning Electron Microscopy and Energy Dispersive X-Ray Analysis in Rapid Microbial Detection and Identification at the Single Cell Level. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Scotuzzi, M.; Kuipers, J.; Wensveen, D.I.; De Boer, P.; Hagen, K.W.; Hoogenboom, J.P.; Giepmans, B. Multi-color electron microscopy by element-guided identification of cells, organelles and molecules. Sci. Rep. 2017, 7, 45970. [Google Scholar] [CrossRef]
- Christner, M.; Rohde, H.; Wolters, M.; Sobottka, I.; Wegscheider, K.; Aepfelbacher, M. Rapid Identification of Bacteria from Positive Blood Culture Bottles by Use of Matrix-Assisted Laser Desorption-Ionization Time of Flight Mass Spectrometry Fingerprinting. J. Clin. Microbiol. 2010, 48, 1584–1591. [Google Scholar] [CrossRef] [Green Version]
- Kourkoumelis, N.; Balatsoukas, I.; Tzaphlidou, M. Ca/P concentration ratio at different sites of normal and osteoporotic rabbit bones evaluated by Auger and energy dispersive X-ray spectroscopy. J. Phys. Chem. B 2012, 38, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, N.; Midiri, A.; Mancuso, G.; Patanè, F.; Venza, M.; Venza, I.; Passantino, A.; Galbo, R.; Teti, G.; Beninati, C.; et al. Neutrophils Directly Recognize Group B Streptococci and Contribute to Interleukin-1β Production during Infection. PLoS ONE 2016, 11, e0160249. [Google Scholar] [CrossRef]
- Johansson, D.; Shannon, O.; Rasmussen, M. Platelet and Neutrophil Responses to Gram Positive Pathogens in Patients with Bacteremic Infection. PLoS ONE 2011, 6, e26928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poller, D.N.; Curry, A.; Ganguli, L.A.; Routledge, R.C. Bacterial calcification in infective endocarditis. Postgrad. Med. J. 1989, 65, 665–667. [Google Scholar] [CrossRef]
- Doern, C.D.; Burnham, C.-A.D. It’s Not Easy Being Green: The Viridans Group Streptococci, with a Focus on Pediatric Clinical Manifestations. J. Clin. Microbiol. 2010, 48, 3829–3835. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-H.; Cheng, C.-L.; Huang, P.-J.; Lin, S.-Y. Application of scanning electron microscopy and X-ray microanalysis: FE-SEM, ESEM-EDS, and EDS mapping for studying the characteristics of topographical microstructure and elemental mapping of human cardiac calcified deposition. Anal. Bioanal. Chem. 2014, 406, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Schwaderer, A.L.; Wolfe, A.J. The association between bacteria and urinary stones. Ann. Transl. Med. 2017, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Carpentier, X.; Daudon, M.; Traxer, O.; Jungers, P.; Mazouyes, A.; Matzen, G.; Véron, E.; Bazin, D. Relationships Between Carbonation Rate of Carbapatite and Morphologic Characteristics of Calcium Phosphate Stones and Etiology. Urology 2009, 73, 968–975. [Google Scholar] [CrossRef]
- Reid, K.; Ball, B.; Zhang, T. Accounting for the Risks of Phosphorus Losses through Tile Drains in a Phosphorus Index. J. Environ. Qual. 2012, 41, 1720–1729. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient 1 | Patient 2 | Patient 3 | Patient 4 | Patient 5 | Patient 6 | |
|---|---|---|---|---|---|---|
| Clinical data | ||||||
| Sex | Male | Male | Male | Female | Male | Female |
| Age (Y) | 88 | 87 | 52 | 93 | 71 | 67 |
| Weight (kg) | 78 | 84 | 83 | 52 | 75 | 64 |
| Diabetes | No | Yes | No | No | No | No |
| Smoking | No | No | Yes | No | Yes | No |
| Hemodialysis | No | No | No | No | No | No |
| Myocardial infarct | No | No | No | No | No | No |
| History of stroke | No | No | No | No | No | No |
| HTA | Yes | Yes | No | Yes | Yes | No |
| Atrial fibrillation | Yes | No | No | No | No | |
| Hist. of malignancy | No | No | No | No | No | Yes |
| IV drug use | No | No | No | No | No | No |
| Localization | Mitral valve | Aortic valve | Mitral valve | Mitral valve | Mitral valve | Aortic valve |
| Echocardiography | ||||||
| Vegetation | No | Aortic | Mitral | Mitral | Mitral | Aortic |
| Vegetation length (mm) | N/A | 6 | 30 | 20 | 18 | 13 |
| Severe regurgitation | Yes | No | No | No | No | No |
| Valve perforation | No | No | No | No | Yes | Yes |
| Abscess | No | Yes | No | No | Yes | No |
| Antibiotherapy | No | Amoxicilline cefriaxone | Daptomycine Sulfamétroxazole trimetroprime | ceftriaxone | Gentamycine and ceftriaxone | Gentamycine and amoxicilline |
| Duration of antibiotherapy before surgery (days) | No | 9 days | No | 15 days | 15 days | 15 days |
| Biology | ||||||
| Hemoglobin (g/L) | 14.7 | 116 | 105 | 120 | 99 | 111 |
| Platelets (G/L) | 161 | 214 | 173 | 198 | 244 | 230 |
| CRP (mg/L) | 10 | 180 | 118 | 118 | 52 | 98 |
| Creatinine (μmol/L) | 166 | 67 | 81 | 81 | 76 | 94.5 |
| BNP (ng/L) | 900 | 405 | 207 | 328 | 534 | 1336 |
| Ca (mmol/L) | 2.10 | 2.05 | 2.01 | 1.99 | 2,19 | 2.04 |
| P (mmol/L) | 1.5 | 1.34 | 0.9 | 1.08 | 1.02 | 1.34 |
| Bacteriology | Negative | E. faecalis | S. aureus | S. agalactiae | S. Gallolyticus | S. oralis |
| Species | General Aspect | Cocci | Platelets | Fibrin | Erythrocytes | Leukocytes | Platelets/Bacteria Ratio |
|---|---|---|---|---|---|---|---|
| (Degenerative valve) | Fibrous laminate, few cells | − | − | − | ++ (On the surface) | − | − |
| E. faecalis | Amorphous laminate | + | + | + | +++ (Major component) | ++ | 1 |
| S. aureus | Amorphous and filamentous | +++ | +++ | +++ | ++ (Incorporated into the fibrin network) | +++ | 1 |
| S. agalactiae | Amorphous and filamentous | ++ | ++ | +++ | ++ (Incorporated into the fibrin network) | ++ | 1 |
| S. gallolyticus | Amorphous and filamentous | +++ | +++ | + | ++ (On the surface) | + | 1 |
| S. oralis | Amorphous and filamentous | +++ | +++ | +++ | ++ (On the surface) | + | 10 |
| C | O | N | Ca | P | ||
|---|---|---|---|---|---|---|
| Weight % | Non-bright regions | 60.2 ± 2.05 | 21.4 ± 0.64 | 17.75 ± 2.4 | 0.15 ± 0.12 | 0.08 ± 0.06 |
| Bright regions | 49.95 ± 3.28 *** | 29.88 ± 3.72 *** | 15.88 ± 1.46 | 2.63 ± 0.61 *** | 1.22 ± 0.28 *** | |
| Atomic % | Non-bright regions | 65.62 ± 2.05 | 17.52 ± 0.55 | 16.6 ± 2.27 | 0.05 ± 0.04 | 0.03 ± 0.026 |
| Bright regions | 57.09 ± 3.05 *** | 24.85 ± 4.72 ** | 16.4 ± 3.09 | 0.9 ± 0.22 *** | 0.65 ± 0.36 ** | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hannachi, N.; Lepidi, H.; Fontanini, A.; Takakura, T.; Bou-Khalil, J.; Gouriet, F.; Habib, G.; Raoult, D.; Camoin-Jau, L.; Baudoin, J.-P. A Novel Approach for Detecting Unique Variations among Infectious Bacterial Species in Endocarditic Cardiac Valve Vegetation. Cells 2020, 9, 1899. https://doi.org/10.3390/cells9081899
Hannachi N, Lepidi H, Fontanini A, Takakura T, Bou-Khalil J, Gouriet F, Habib G, Raoult D, Camoin-Jau L, Baudoin J-P. A Novel Approach for Detecting Unique Variations among Infectious Bacterial Species in Endocarditic Cardiac Valve Vegetation. Cells. 2020; 9(8):1899. https://doi.org/10.3390/cells9081899
Chicago/Turabian StyleHannachi, Nadji, Hubert Lepidi, Anthony Fontanini, Tatsuki Takakura, Jacques Bou-Khalil, Frédérique Gouriet, Gilbert Habib, Didier Raoult, Laurence Camoin-Jau, and Jean-Pierre Baudoin. 2020. "A Novel Approach for Detecting Unique Variations among Infectious Bacterial Species in Endocarditic Cardiac Valve Vegetation" Cells 9, no. 8: 1899. https://doi.org/10.3390/cells9081899
APA StyleHannachi, N., Lepidi, H., Fontanini, A., Takakura, T., Bou-Khalil, J., Gouriet, F., Habib, G., Raoult, D., Camoin-Jau, L., & Baudoin, J. -P. (2020). A Novel Approach for Detecting Unique Variations among Infectious Bacterial Species in Endocarditic Cardiac Valve Vegetation. Cells, 9(8), 1899. https://doi.org/10.3390/cells9081899