Expression of the Y-Encoded TSPY is Associated with Progression of Prostate Cancer


Abstract
:1. Introduction
2. Results and Discussion
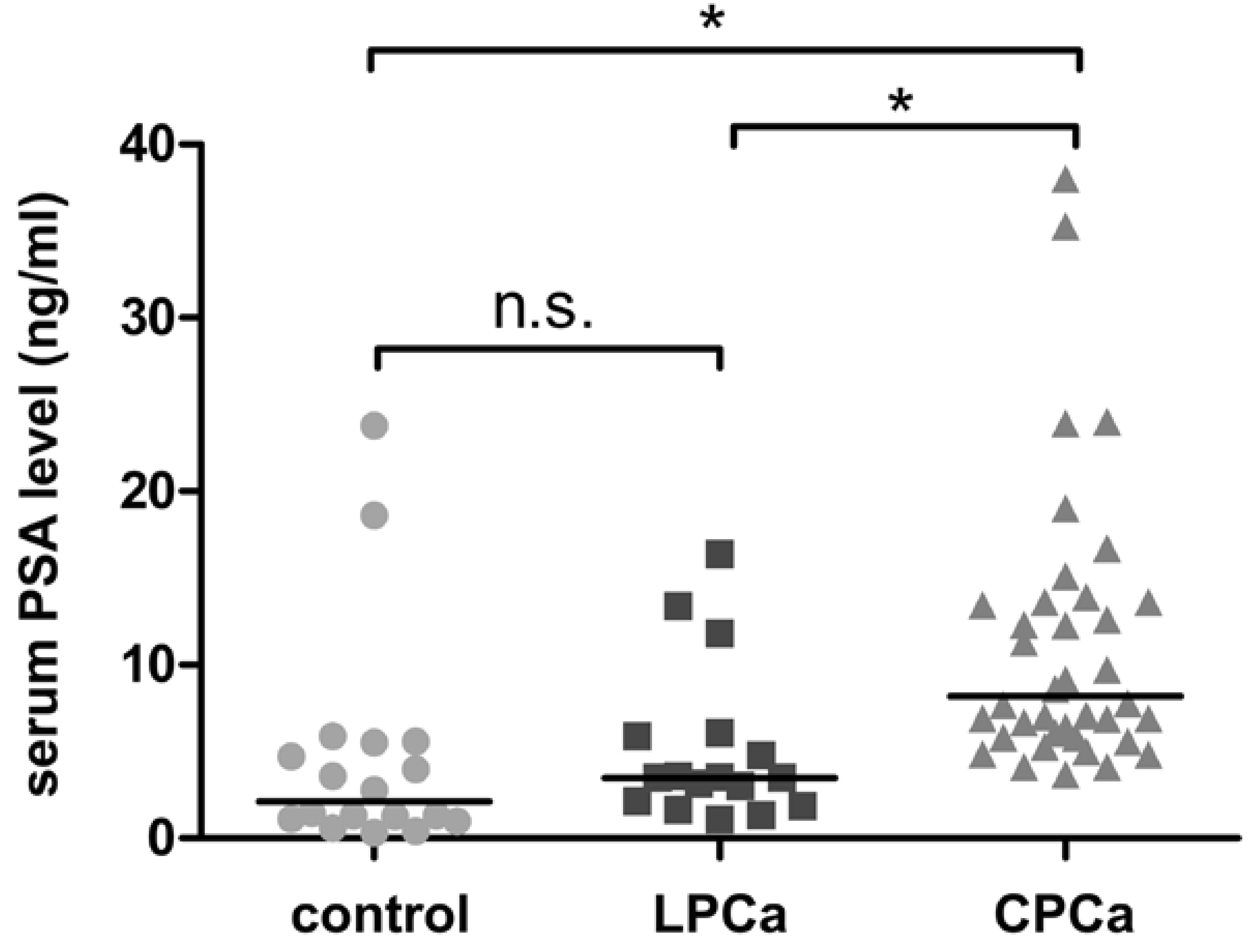
2.1. Characteristics of Normal Prostate, Latent and Clinical Prostate Cancer Specimens

{kind=link}
{kind=link}
{kind=link}
| Controls | Latent Cancer | Clinical Cancer | |
|---|---|---|---|
| (n = 18) | (n = 17) | (n = 41) | |
| Mean ± SD age (years) | 69.3 ± 8.3 | 70.9 ± 4.2 | 67.6 ± 5.1* |
| No. Gleason sum (%) | |||
| GS < 7 | – | 8 (47%) | 17 (41%) |
| GS ≥ 7 | – | 9 (53%) | 24 (59%) |
| No. ng/ml serum PSA | |||
| PSA < 10 | 16 (89%) | 14 (82%) | 24 (59%) |
| PSA ≥ 10 | 2 (11%) | 3 (18%) | 17 (41%)** |

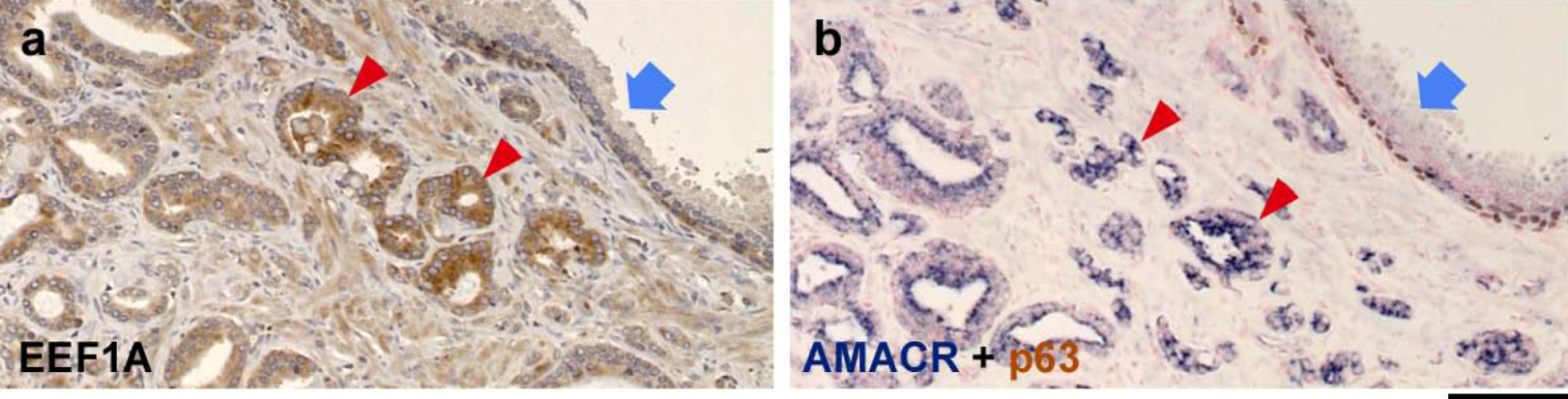
2.2. TSPY Expression in Human Prostate Cancer

| Non cancer | Latent Cancer | Clinical Cancer | |
|---|---|---|---|
| Normal gland | 9/18 (50%) | 10/17 (59%) | 33/41 (80%)* |
| Cancer cells | nd | 8/17 (47%) | 32/41 (78%)* |
| Gleason score | ||
|---|---|---|
| GS < 7 | 7 ≤ GS | |
| Latent cancer | 1/8 (13%)* | 7/9 (78%)* |
| Clinical cancer | 13/17 (76%) | 19/24 (79%) |
2.3. EEF1A Expression is Elevated in Human Prostate Cancer
| Latent cancer | Clinical cancer | ||
|---|---|---|---|
| EEF1A intensity | |||
| cancer < normal | 1 (9%) | 1 (4%) | |
| cancer = normal | 7 (64%) | 16 (64%) | |
| cancer > normal | 3 (27%) | 8 (32%) |

3. Experimental Section
3.1. Prostate Specimens
3.2. Immunohistochemistry
4. Conclusions
Acknowledgments
References and Notes
- Arnemann, J.; Epplen, J.T.; Cooke, H.J.; Sauermann, U.; Engel, W.; Schmidtke, J. A human Y-chromosomal DNA sequence expressed in testicular tissue. Nucleic Acids Res. 1987, 15, 8713–8724. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Yang-Feng, T.L.; Muller, U.; Mohandas, T.K.; de Jong, P.J.; Lau, Y.F. Molecular isolation and characterization of an expressed gene from the human Y chromosome. Hum. Mol. Genet. 1992, 1, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Schnieders, F.; Dork, T.; Arnemann, J.; Vogel, T.; Werner, M.; Schmidtke, J. Testis-specific protein, Y-encoded (TSPY) expression in testicular tissues. Hum. Mol. Genet. 1996, 5, 1801–1807. [Google Scholar] [CrossRef] [PubMed]
- Kersemaekers, A.M.; Honecker, F.; Stoop, H.; Cools, M.; Molier, M.; Wolffenbuttel, K.; Bokemeyer, C.; Li, Y.; Lau, Y.F.; Oosterhuis, J.W.; Looijenga, L.H. Identification of germ cells at risk for neoplastic transformation in gonadoblastoma: an immunohistochemical study for OCT3/4 and TSPY. Hum. Pathol. 2005, 36, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, K.; Reijo, R.; Page, D.C.; Disteche, C.M. Gonadoblastoma: molecular definition of the susceptibility region on the Y chromosome. Am. J. Hum. Genet. 1995, 57, 1400–1407. [Google Scholar] [PubMed]
- Salo, P.; Kaariainen, H.; Petrovic, V.; Peltomaki, P.; Page, D.C.; de la Chapelle, A. Molecular mapping of the putative gonadoblastoma locus on the Y chromosome. Genes Chromosomes Cancer 1995, 14, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Page, D.C. Hypothesis: a Y-chromosomal gene causes gonadoblastoma in dysgenetic gonads . Development 1987, Suppl. 101, 151–155. [Google Scholar] [PubMed]
- Lau, Y.; Chou, P.; Iezzoni, J.; Alonzo, J.; Komuves, L. Expression of a candidate gene for the gonadoblastoma locus in gonadoblastoma and testicular seminoma. Cytogenet. Cell Genet. 2000, 91, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Dubovsky, J.A.; McNeel, D.G. Inducible expression of a prostate cancer-testis antigen, SSX-2, following treatment with a DNA methylation inhibitor. Prostate 2007, 67, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, W.M.; Bergin, O.E.; Rafferty, M.; Kelly, Z.D.; Nolan, I.M.; Fox, E.J.; Culhane, A.C.; McArdle, L.; Fraga, M.F.; Hughes, L.; Currid, C.A.; O'Mahony, F.; Byrne, A.; Murphy, A.A.; Moss, C.; McDonnell, S.; Stallings, R.L.; Plumb, J.A.; Esteller, M.; Brown, R.; Dervan, P.A.; Easty, D.J. Multiple markers for melanoma progression regulated by DNA methylation: insights from transcriptomic studies. Carcinogenesis 2005, 26, 1856–1867. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.H.; Li, Y.Y.; Qiao, H.; Wang, H.C.; Yang, X.A.; Zhang, H.G.; Pang, X.W.; Zhang, Y.; Chen, W.F. TSPY is a cancer testis antigen expressed in human hepatocellular carcinoma. Br. J. Cancer 2005, 93, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, M.J.; Simpson, A.J.; Old, L.J. The cancer/testis genes: Review, standardization, and commentary. Cancer Immun. 2004, 4, 1. [Google Scholar] [PubMed]
- Simpson, A.J.; Caballero, O.L.; Jungbluth, A.; Chen, Y.T.; Old, L.J. Cancer/testis antigens, gametogenesis and cancer. Nat. Rev. Cancer 2005, 5, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.F.; Lau, H.W.; Komuves, L.G. Expression pattern of a gonadoblastoma candidate gene suggests a role of the Y chromosome in prostate cancer. Cytogenet. Genome Res. 2003, 101, 250–260. [Google Scholar] [CrossRef]
- Singh, A.P.; Bafna, S.; Chaudhary, K.; Venkatraman, G.; Smith, L.; Eudy, J.D.; Johansson, S.L.; Lin, M.F.; Batra, S.K. Genome-wide expression profiling reveals transcriptomic variation and perturbed gene networks in androgen-dependent and androgen-independent prostate cancer cells. Cancer Lett. 2008, 259, 28–38. [Google Scholar] [PubMed]
- Yu, Y.P.; Landsittel, D.; Jing, L.; Nelson, J.; Ren, B.; Liu, L.; McDonald, C.; Thomas, R.; Dhir, R.; Finkelstein, S.; Michalopoulos, G.; Becich, M.; Luo, J.H. Gene expression alterations in prostate cancer predicting tumor aggression and preceding development of malignancy. J. Clin. Oncol. 2004, 22, 2790–2799. [Google Scholar] [CrossRef] [PubMed]
- Stanbrough, M.; Bubley, G.J.; Ross, K.; Golub, T.R.; Rubin, M.A.; Penning, T.M.; Febbo, P.G.; Balk, S.P. Increased expression of genes converting adrenal androgens to testosterone in androgen-independent prostate cancer. Cancer Res. 2006, 66, 2815–2825. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Placencio, V.; Iturregui, J.M.; Uwamariya, C.; Sharif-Afshar, A.R.; Koyama, T.; Hayward, S.W.; Bhowmick, N.A. Prostate tumor progression is mediated by a paracrine TGF-beta/Wnt3a signaling axis. Oncogene 2008, 27, 7118–7130. [Google Scholar] [CrossRef] [PubMed]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef]
- Park, Y.J.; Luger, K. Structure and function of nucleosome assembly proteins. Biochem. Cell Biol. 2006, 84, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Muto, S.; Senda, M.; Akai, Y.; Sato, L.; Suzuki, T.; Nagai, R.; Senda, T.; Horikoshi, M. Relationship between the structure of SET/TAF-Ibeta/INHAT and its histone chaperone activity. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 4285–4290. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lau, Y.F. TSPY and its X-encoded homologue interact with cyclin B but exert contrasting functions on cyclin-dependent kinase 1 activities. Oncogene 2008, 27, 6141–6150. [Google Scholar] [CrossRef] [PubMed]
- Kido, T.; Lau, Y.F. The human Y-encoded testis-specific protein interacts functionally with eukaryotic translation elongation factor eEF1A, a putative oncoprotein. Int. J. Cancer 2008, 123, 1573–1585. [Google Scholar] [CrossRef] [PubMed]
- Kahns, S.; Lund, A.; Kristensen, P.; Knudsen, C.R.; Clark, B.F.; Cavallius, J.; Merrick, W.C. The elongation factor 1 A-2 isoform from rabbit: cloning of the cDNA and characterization of the protein. Nucleic Acids Res. 1998, 26, 1884–1890. [Google Scholar] [CrossRef] [PubMed]
- Shamovsky, I.; Ivannikov, M.; Kandel, E.S.; Gershon, D.; Nudler, E. RNA-mediated response to heat shock in mammalian cells. Nature 2006, 440, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Amiri, A.; Noei, F.; Jeganathan, S.; Kulkarni, G.; Pinke, D.E.; Lee, J.M. eEF1A2 activates Akt and stimulates Akt-dependent actin remodeling, invasion and migration. Oncogene 2007, 26, 3027–3040. [Google Scholar] [CrossRef] [PubMed]
- Thornton, S.; Anand, N.; Purcell, D.; Lee, J. Not just for housekeeping: protein initiation and elongation factors in cell growth and tumorigenesis. J. Mol. Med. 2003, 81, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Sweet, H.O.; Davisson, M.T.; Coman, D.R. 'Wasted', a new mutant of the mouse with abnormalities characteristic to ataxia telangiectasia. Nature 1982, 297, 402–404. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Yonehara, S. Novel cell death by downregulation of eEF1A1 expression in tetraploids. Cell Death Differ. 2009, 16, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.W.; Traugh, J.A. Phosphorylation of elongation factor 1 and ribosomal protein S6 by multipotential S6 kinase and insulin stimulation of translational elongation. J. Biol. Chem. 1997, 272, 28252–28257. [Google Scholar] [CrossRef] [PubMed]
- Anand, N.; Murthy, S.; Amann, G.; Wernick, M.; Porter, L.A.; Cukier, I.H.; Collins, C.; Gray, J.W.; Diebold, J.; Demetrick, D.J.; Lee, J.M. Protein elongation factor EEF1A2 is a putative oncogene in ovarian cancer. Nat. Genet. 2002, 31, 301–305. [Google Scholar] [PubMed]
- Li, R.; Wang, H.; Bekele, B.N.; Yin, Z.; Caraway, N.P.; Katz, R.L.; Stass, S.A.; Jiang, F. Identification of putative oncogenes in lung adenocarcinoma by a comprehensive functional genomic approach. Oncogene 2006, 25, 2628–2635. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, V.A.; Newbery, H.J.; Wray, N.R.; Jackson, J.; Larionov, A.; Miller, W.R.; Dixon, J.M.; Abbott, C.M. Translation elongation factor eEF1A2 is a potential oncoprotein that is overexpressed in two-thirds of breast tumours. BMC Cancer 2005, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Sokoloff, M.H.; Yang, X.J.; Fumo, M.; Mhoon, D.; Brendler, C.B. Characterizing prostatic adenocarcinomas in men with a serum prostate specific antigen level of < 4.0 ng/mL . BJU Int. 2004, 93, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Schroder, F.H.; van der Cruijsen-Koeter, I.; de Koning, H.J.; Vis, A.N.; Hoedemaeker, R.F.; Kranse, R. Prostate cancer detection at low prostate specific antigen. J. Urol. 2000, 163, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Zha, S.; Gage, W.R.; Dunn, T.A.; Hicks, J.L.; Bennett, C.J.; Ewing, C.M.; Platz, E.A.; Ferdinandusse, S.; Wanders, R.J.; Trent, J.M.; Isaacs, W.B.; De Marzo, A.M. Alpha-methylacyl-CoA racemase: a new molecular marker for prostate cancer. Cancer Res. 2002, 62, 2220–2226. [Google Scholar] [PubMed]
- Delongchamps, N.B.; Singh, A.; Haas, G.P. The role of prevalence in the diagnosis of prostate cancer. Cancer Control 2006, 13, 158–168. [Google Scholar] [PubMed]
- Takahashi, S.; Suzuki, S.; Takahashi, S.; Inaguma, S.; Asamoto, M.; Shirai, T. Differences between latent and clinical prostate carcinomas: lower cell proliferation activity in latent cases. Prostate 2006, 66, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Wu, X.; Hou, M.; Lee, S.O.; Lou, W.; Wang, J.; Janarthan, B.; Nallapareddy, S.; Trump, D.L.; Gao, A.C. Interleukin-6 polymorphism is associated with more aggressive prostate cancer. J. Urol. 2005, 174, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Oram, S.W.; Liu, X.X.; Lee, T.L.; Chan, W.Y.; Lau, Y.F. TSPY potentiates cell proliferation and tumorigenesis by promoting cell cycle progression in HeLa and NIH3T3 cells. BMC Cancer 2006, 6, 154. [Google Scholar] [CrossRef]
- Kulkarni, G.; Turbin, D.A.; Amiri, A.; Jeganathan, S.; Andrade-Navarro, M.A.; Wu, T.D.; Huntsman, D.G.; Lee, J.M. Expression of protein elongation factor eEF1A2 predicts favorable outcome in breast cancer. Breast Cancer Res. Treat. 2007, 102, 31–41. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Kido, T.; Hatakeyama, S.; Ohyama, C.; Lau, Y.-F.C. Expression of the Y-Encoded TSPY is Associated with Progression of Prostate Cancer. Genes 2010, 1, 283-293. https://doi.org/10.3390/genes1020283
Kido T, Hatakeyama S, Ohyama C, Lau Y-FC. Expression of the Y-Encoded TSPY is Associated with Progression of Prostate Cancer. Genes. 2010; 1(2):283-293. https://doi.org/10.3390/genes1020283
Chicago/Turabian StyleKido, Tatsuo, Shingo Hatakeyama, Chikara Ohyama, and Yun-Fai Chris Lau. 2010. "Expression of the Y-Encoded TSPY is Associated with Progression of Prostate Cancer" Genes 1, no. 2: 283-293. https://doi.org/10.3390/genes1020283
APA StyleKido, T., Hatakeyama, S., Ohyama, C., & Lau, Y. -F. C. (2010). Expression of the Y-Encoded TSPY is Associated with Progression of Prostate Cancer. Genes, 1(2), 283-293. https://doi.org/10.3390/genes1020283