Programming Pluripotent Precursor Cells Derived from Xenopus Embryos to Generate Specific Tissues and Organs

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Programming ACC to Generate Specific Tissues and Organs
2.1. The Experimental Basis
- a)
- ACC can be exposed to extracellular signaling molecules like growth factors or chemical agents affecting signal transduction. Recombinant growth factors can be applied for short intervals or throughout the incubation time. Alternatively, growth factors can be produced by the ACC themselves. To this end, fertilized embryos are injected with mRNA or DNA coding for the respective growth factor prior to dissection of the ACC.
- b)
- Intracellularly active proteins, such as transcription factors, signal transduction proteins, etc. can also be supplied by microinjection of the corresponding mRNAs. As a variation of this procedure, microinjected explants can be sandwiched with naïve ACC, allowing for the distinction of cell-autonomous and non-cell-autonomous effects.
- c)
- The developmental potential of specific tissues can be analyzed by heterotopic “Sandwich” assays combining ACC with tissue explants from the same or different embryonic sources.
- d)
- Manipulated ACC can be transplanted into recipient embryos to analyze if they can form functional ectopic organs.
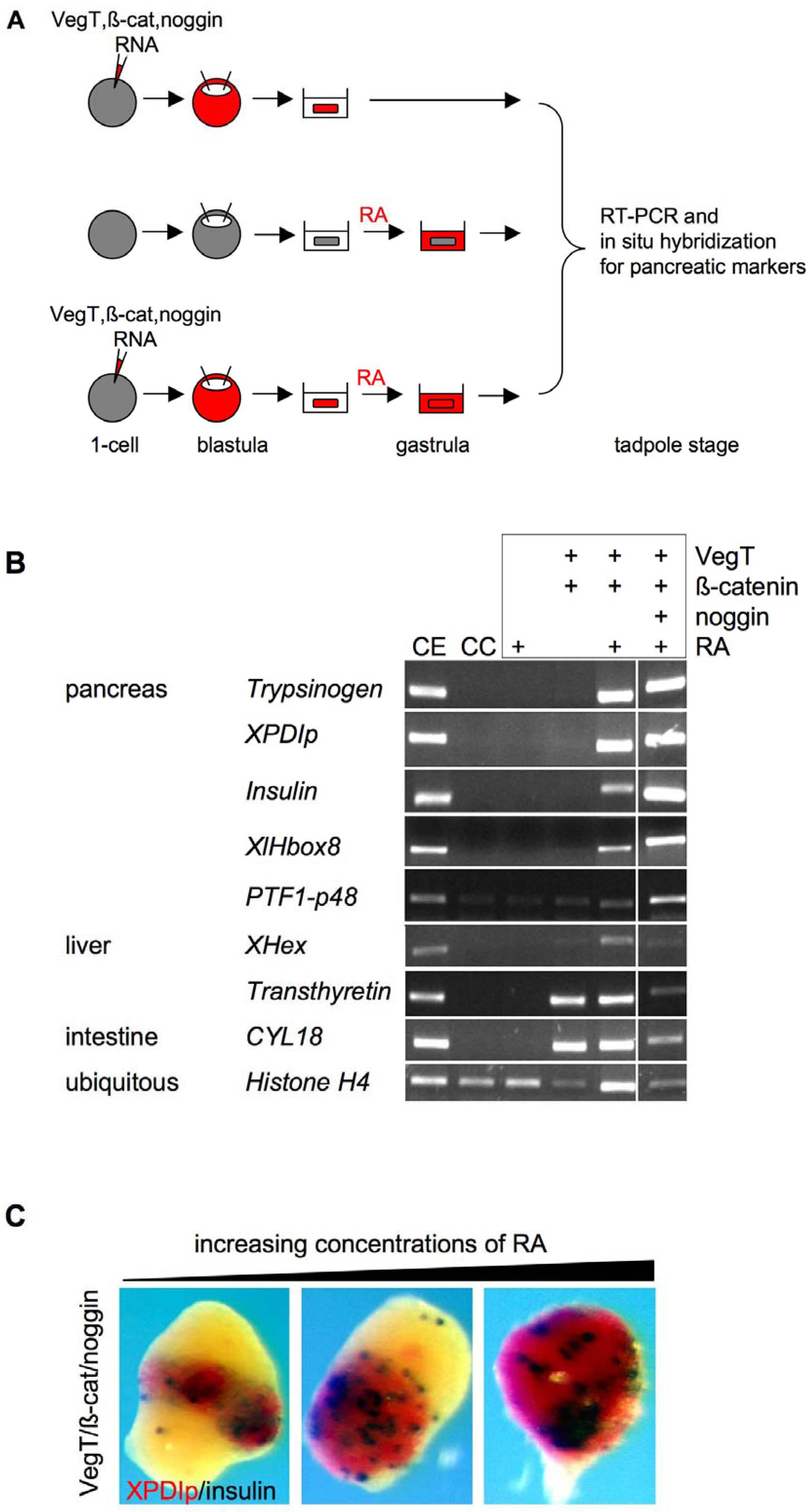
2.2. Pancreas and Liver Development

2.3. Kidney Development
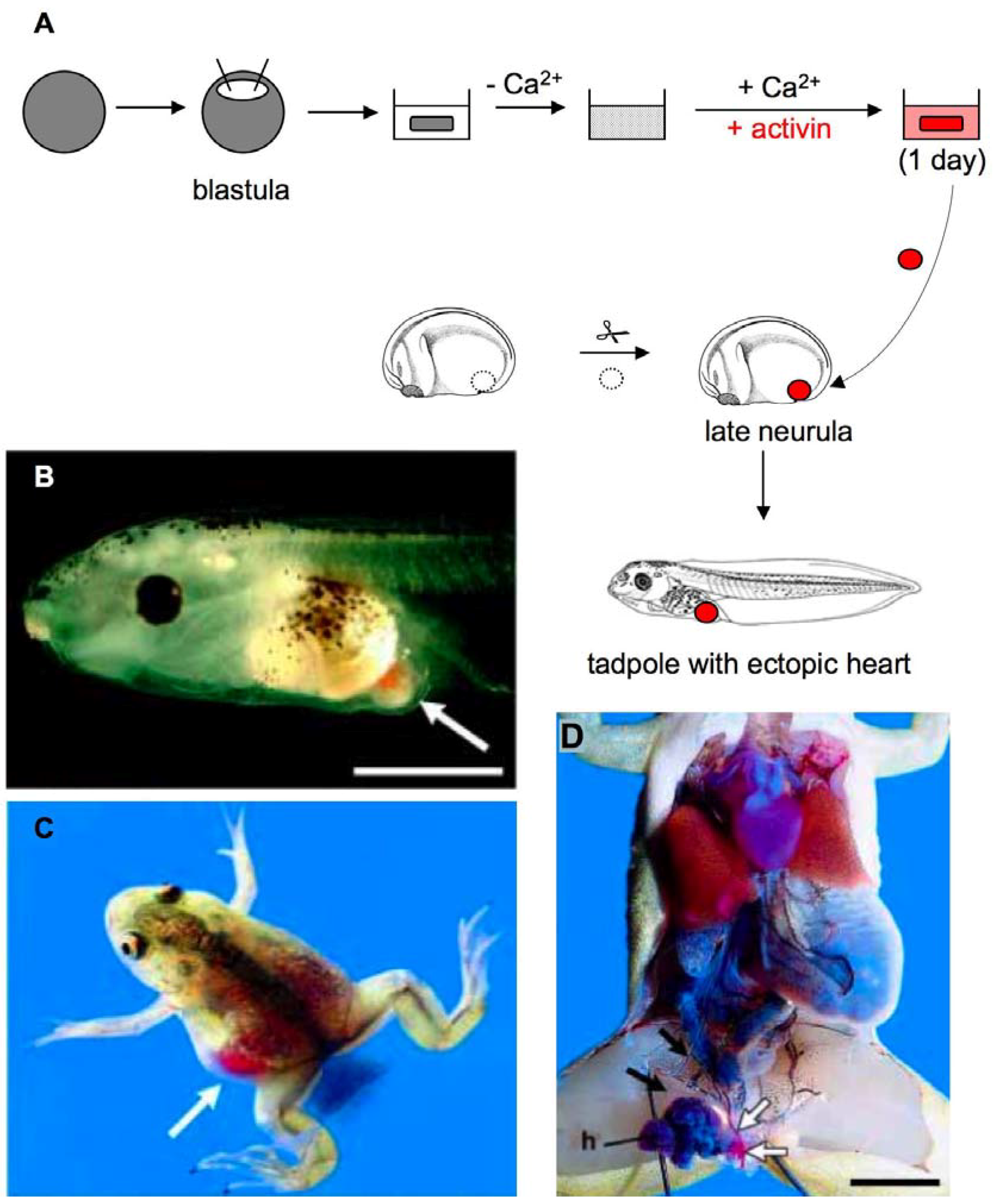
2.4. Heart Development
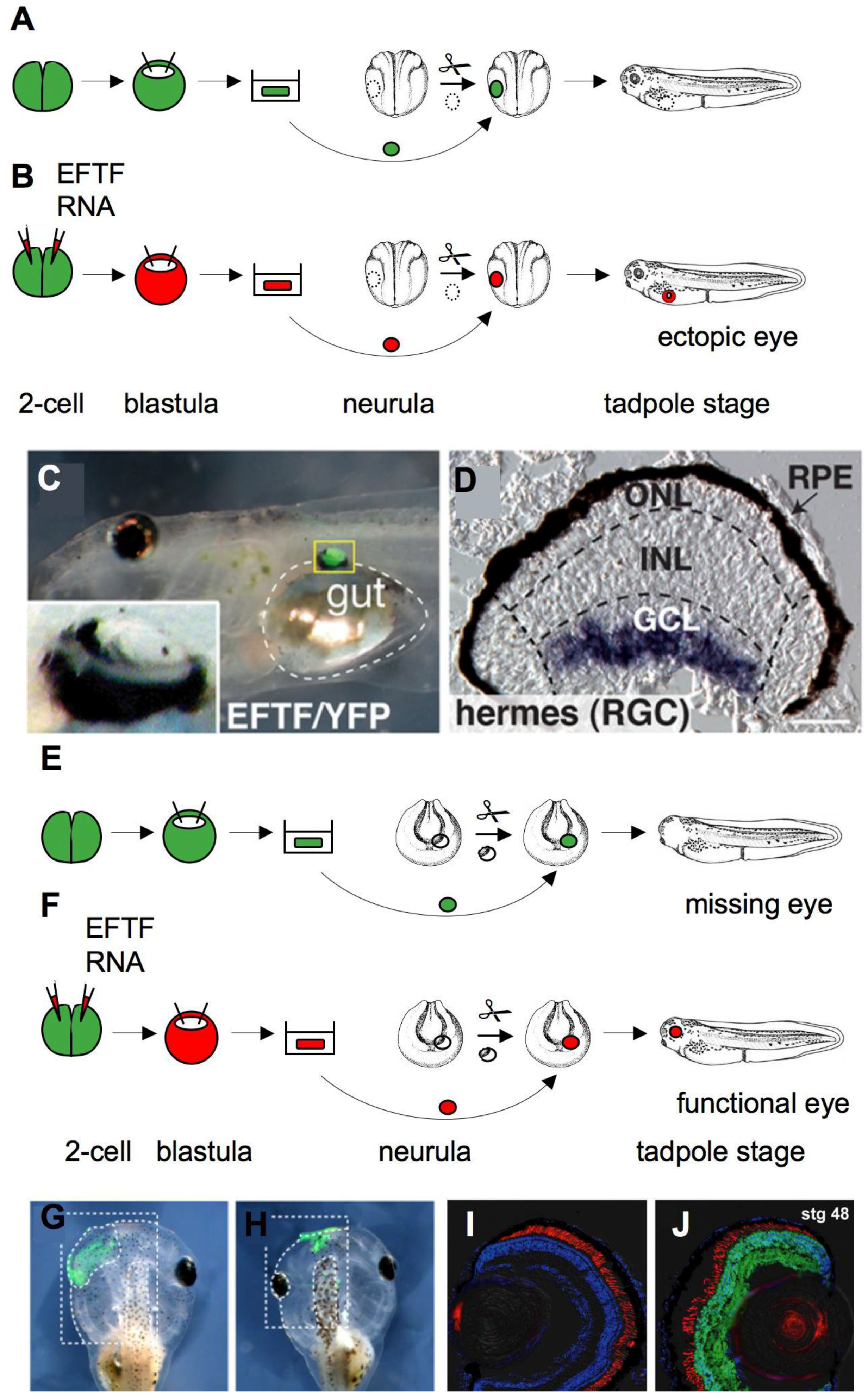
2.5. Eye Development


3. Conclusions
Acknowledgements
References and Notes
- Spemann, H. Experimentelle Beiträge zu einer Theorie der Entwicklung; Springer: Berlin, Germany, 1936. [Google Scholar]
- De Robertis, E.M. Spemann’s organizer and self-regulation in amphibian embryos. Nat. Rev. Mol. Cell Biol. 2006, 7, 296–302. [Google Scholar] [CrossRef]
- Nieuwkoop, P.D. Pattern Formation in Artificially Activated Ectoderm (Rana Pipiens and Ambystoma Punctatum). Dev. Biol. 1963, 7, 255–279. [Google Scholar] [CrossRef]
- Dale, L.; Slack, J.M. Regional specification within the mesoderm of early embryos of Xenopus laevis. Development 1987, 100, 279–295. [Google Scholar]
- Hemmati-Brivanlou, A.; Melton, D.A. A truncated activin receptor inhibits mesoderm induction and formation of axial structures in Xenopus embryos. Nature 1992, 359, 609–614. [Google Scholar] [CrossRef]
- Lamb, T.M.; Harland, R.M. Fibroblast growth factor is a direct neural inducer, which combined with noggin generates anterior-posterior neural pattern. Development 1995, 121, 3627–3636. [Google Scholar]
- Clements, D.; Woodland, H.R. VegT induces endoderm by a self-limiting mechanism and by changing the competence of cells to respond to TGF-beta signals. Dev. Biol. 2003, 258, 454–463. [Google Scholar] [CrossRef]
- Henry, G.L.; Melton, D.A. Mixer, a homeobox gene required for endoderm development. Science 1998, 281, 91–96. [Google Scholar] [CrossRef]
- Horb, M.E.; Thomsen, G.H. A vegetally localized T-box transcription factor in Xenopus eggs specifies mesoderm and endoderm and is essential for embryonic mesoderm formation. Development 1997, 124, 1689–1698. [Google Scholar]
- Asashima, M.; Ito, Y.; Chan, T.; Michiue, T.; Nakanishi, M.; Suzuki, K.; Hitachi, K.; Okabayashi, K.; Kondow, A.; Ariizumi, T. In vitro organogenesis from undifferentiated cells in Xenopus. Dev. Dyn. 2009, 238, 1309–1320. [Google Scholar] [CrossRef]
- Fukui, Y.; Furue, M.; Myoishi, Y.; Sato, J.D.; Okamoto, T.; Asashima, M. Long-term culture of Xenopus presumptive ectoderm in a nutrient-supplemented culture medium. Dev. Growth Differ. 2003, 45, 499–506. [Google Scholar] [CrossRef]
- Zaret, K.S.; Grompe, M. Generation and regeneration of cells of the liver and pancreas. Science 2008, 322, 1490–1494. [Google Scholar] [CrossRef]
- Moriya, N.; Komazaki, S.; Takahashi, S.; Yokota, C.; Asashima, M. In vitro pancreas formation from Xenopus ectoderm treated with activin and retinoic acid. Dev. Growth Differ. 2000, 42, 593–602. [Google Scholar] [CrossRef]
- Chen, Y.; Jurgens, K.; Hollemann, T.; Claussen, M.; Ramadori, G.; Pieler, T. Cell-autonomous and signal-dependent expression of liver and intestine marker genes in pluripotent precursor cells from Xenopus embryos. Mech. Dev. 2003, 120, 277–288. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, F.C.; Brandes, N.; Afelik, S.; Solter, M.; Pieler, T. Retinoic acid signaling is essential for pancreas development and promotes endocrine at the expense of exocrine cell differentiation in Xenopus. Dev. Biol. 2004, 271, 144–160. [Google Scholar] [CrossRef]
- Pan, F.C.; Chen, Y.; Loeber, J.; Henningfeld, K.; Pieler, T. I-SceI meganuclease-mediated transgenesis in Xenopus. Dev. Dyn. 2006, 235, 247–252. [Google Scholar] [CrossRef]
- Afelik, S.; Chen, Y.; Pieler, T. Combined ectopic expression of Pdx1 and Ptf1a/p48 results in the stable conversion of posterior endoderm into endocrine and exocrine pancreatic tissue. Genes Dev. 2006, 20, 1441–1446. [Google Scholar] [CrossRef]
- Pan, F.C.; Afelik, S.; Pieler, T.; University of Göttingen, Göttingen, Germany. Unpublished work; 2006.
- Borowiak, M.; Melton, D.A. How to make beta cells? Curr. Opin. Cell Biol. 2009, 21, 727–732. [Google Scholar] [CrossRef]
- Mayhew, C.N.; Wells, J.M. Converting human pluripotent stem cells into beta-cells: Recent advances and future challenges. Curr. Opin. Organ. Transplant 2009, 15, 54–60. [Google Scholar]
- Nakanishi, M.; Hamazaki, T.S.; Komazaki, S.; Okochi, H.; Asashima, M. Pancreatic tissue formation from murine embryonic stem cells in vitro. Differentiation 2007, 75, 1–11. [Google Scholar] [CrossRef]
- Brandli, A.W. Towards a molecular anatomy of the Xenopus pronephric kidney. Int. J. Dev. Biol. 1999, 43, 381–395. [Google Scholar]
- Moriya, N.; Uchiyama, H.; Asashima, M. Induction of Pronephric Tubules by Activin and Retinoic Acid in Presumptive Ectoderm of Xenopus laevis. Dev. Growth Differ. 1993, 35, 123–128. [Google Scholar]
- Heller, N.; Brandli, A.W. Xenopus Pax-2/5/8 orthologues: novel insights into Pax gene evolution and identification of Pax-8 as the earliest marker for otic and pronephric cell lineages. Dev. Genet. 1999, 24, 208–219. [Google Scholar] [CrossRef]
- Osafune, K.; Nishinakamura, R.; Komazaki, S.; Asashima, M. In vitro induction of the pronephric duct in Xenopus explants. Dev. Growth Differ. 2002, 44, 161–167. [Google Scholar] [CrossRef]
- Eid, S.R.; Terrettaz, A.; Nagata, K.; Brandli, A.W. Embryonic expression of Xenopus SGLT-1L, a novel member of the solute carrier family 5 (SLC5), is confined to tubules of the pronephric kidney. Int. J. Dev. Biol. 2002, 46, 177–184. [Google Scholar]
- Chan, T.C.; Ariizumi, T.; Asashima, M. A model system for organ engineering: transplantation of in vitro induced embryonic kidney. Naturwissenschaften 1999, 86, 224–227. [Google Scholar] [CrossRef]
- Tetelin, S.; Jones, E.A. Xenopus Wnt11b is identified as a potential pronephric inducer. Dev. Dyn. 239, 148–159.
- Afouda, B.A.; Hoppler, S. Xenopus explants as an experimental model system for studying heart development. Trends Cardiovasc. Med. 2009, 19, 220–226. [Google Scholar] [CrossRef]
- Warkman, A.S.; Krieg, P.A. Xenopus as a model system for vertebrate heart development. Semin. Cell Dev. Biol. 2007, 18, 46–53. [Google Scholar] [CrossRef]
- Grunz, H. Suramin changes the fate of Spemann’s organizer and prevents neural induction in Xenopus laevis. Mech. Dev. 1992, 38, 133–141. [Google Scholar] [CrossRef]
- Grunz, H. Amphibian embryos as a model system for organ engineering: in vitro induction and rescue of the heart anlage. Int. J. Dev. Biol. 1999, 43, 361–364. [Google Scholar]
- Latinkic, B.V.; Kotecha, S.; Mohun, T.J. Induction of cardiomyocytes by GATA4 in Xenopus ectodermal explants. Development 2003, 130, 3865–3876. [Google Scholar] [CrossRef]
- Pandur, P.; Lasche, M.; Eisenberg, L.M.; Kuhl, M. Wnt-11 activation of a non-canonical Wnt signalling pathway is required for cardiogenesis. Nature 2002, 418, 636–641. [Google Scholar] [CrossRef]
- Zhang, C.; Basta, T.; Klymkowsky, M.W. SOX7 and SOX18 are essential for cardiogenesis in Xenopus. Dev. Dyn. 2005, 234, 878–891. [Google Scholar] [CrossRef]
- Ariizumi, T.; Komazaki, S.; Asashima, M.; Malacinski, G.M. Activin treated urodele ectoderm: A model experimental system for cardiogenesis. Int. J. Dev. Biol. 1996, 40, 715–718. [Google Scholar]
- Ariizumi, T.; Kinoshita, M.; Yokota, C.; Takano, K.; Fukuda, K.; Moriyama, N.; Malacinski, G.M.; Asashima, M. Amphibian in vitro heart induction: A simple and reliable model for the study of vertebrate cardiac development. Int. J. Dev. Biol. 2003, 47, 405–410. [Google Scholar]
- Afouda, B.A.; Martin, J.; Liu, F.; Ciau-Uitz, A.; Patient, R.; Hoppler, S. GATA transcription factors integrate Wnt signalling during heart development. Development 2008, 135, 3185–3190. [Google Scholar] [CrossRef]
- Samuel, L.J.; Latinkic, B.V. Early activation of FGF and nodal pathways mediates cardiac specification independently of Wnt/beta-catenin signaling. PLoS One 2009, 4, e7650. [Google Scholar] [CrossRef]
- Sedohara, A.; Komazaki, S.; Asashima, M. In vitro induction and transplantation of eye during early Xenopus development. Dev. Growth Differ. 2003, 45, 463–471. [Google Scholar] [CrossRef]
- Lan, L.; Vitobello, A.; Bertacchi, M.; Cremisi, F.; Vignali, R.; Andreazzoli, M.; Demontis, G.C.; Barsacchi, G.; Casarosa, S. Noggin elicits retinal fate in Xenopus animal cap embryonic stem cells. Stem Cells 2009, 27, 2146–2152. [Google Scholar] [CrossRef]
- Viczian, A.S.; Solessio, E.C.; Lyou, Y.; Zuber, M.E. Generation of functional eyes from pluripotent cells. PLoS Biol. 2009, 7, e1000174. [Google Scholar] [CrossRef]
- Cartry, J.; Nichane, M.; Ribes, V.; Colas, A.; Riou, J.F.; Pieler, T.; Dolle, P.; Bellefroid, E.J.; Umbhauer, M. Retinoic acid signalling is required for specification of pronephric cell fate. Dev. Biol. 2006, 299, 35–51. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Borchers, A.; Pieler, T. Programming Pluripotent Precursor Cells Derived from Xenopus Embryos to Generate Specific Tissues and Organs. Genes 2010, 1, 413-426. https://doi.org/10.3390/genes1030413
Borchers A, Pieler T. Programming Pluripotent Precursor Cells Derived from Xenopus Embryos to Generate Specific Tissues and Organs. Genes. 2010; 1(3):413-426. https://doi.org/10.3390/genes1030413
Chicago/Turabian StyleBorchers, Annette, and Tomas Pieler. 2010. "Programming Pluripotent Precursor Cells Derived from Xenopus Embryos to Generate Specific Tissues and Organs" Genes 1, no. 3: 413-426. https://doi.org/10.3390/genes1030413
APA StyleBorchers, A., & Pieler, T. (2010). Programming Pluripotent Precursor Cells Derived from Xenopus Embryos to Generate Specific Tissues and Organs. Genes, 1(3), 413-426. https://doi.org/10.3390/genes1030413