Genome-Wide Identification and Expression Profiling of Sugar Transporter Protein (STP) Family Genes in Cabbage (Brassica oleracea var. capitata L.) Reveals their Involvement in Clubroot Disease Responses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Sugar Transporter Proteins in Brassica oleracea
2.2. Characterization and Phylogenetic Analysis of BoSTPs
2.3. Chromosomal Localization and the Calculation of Ka/Ks
2.4. Prediction of TMH, Putative Functional Domain, and Subcellular Localization
2.5. Expression Profile Analysis of BoSTP Genes
2.6. Subcellular Localization Analysis of BoSTPs in Tobacco
3. Results
3.1. Identification and Phylogenetic Analysis of STPs in Brassica oleracea, Brassica rapa, and Arabidopsis thaliana
3.2. Chromosomal Distribution of BoSTP Genes
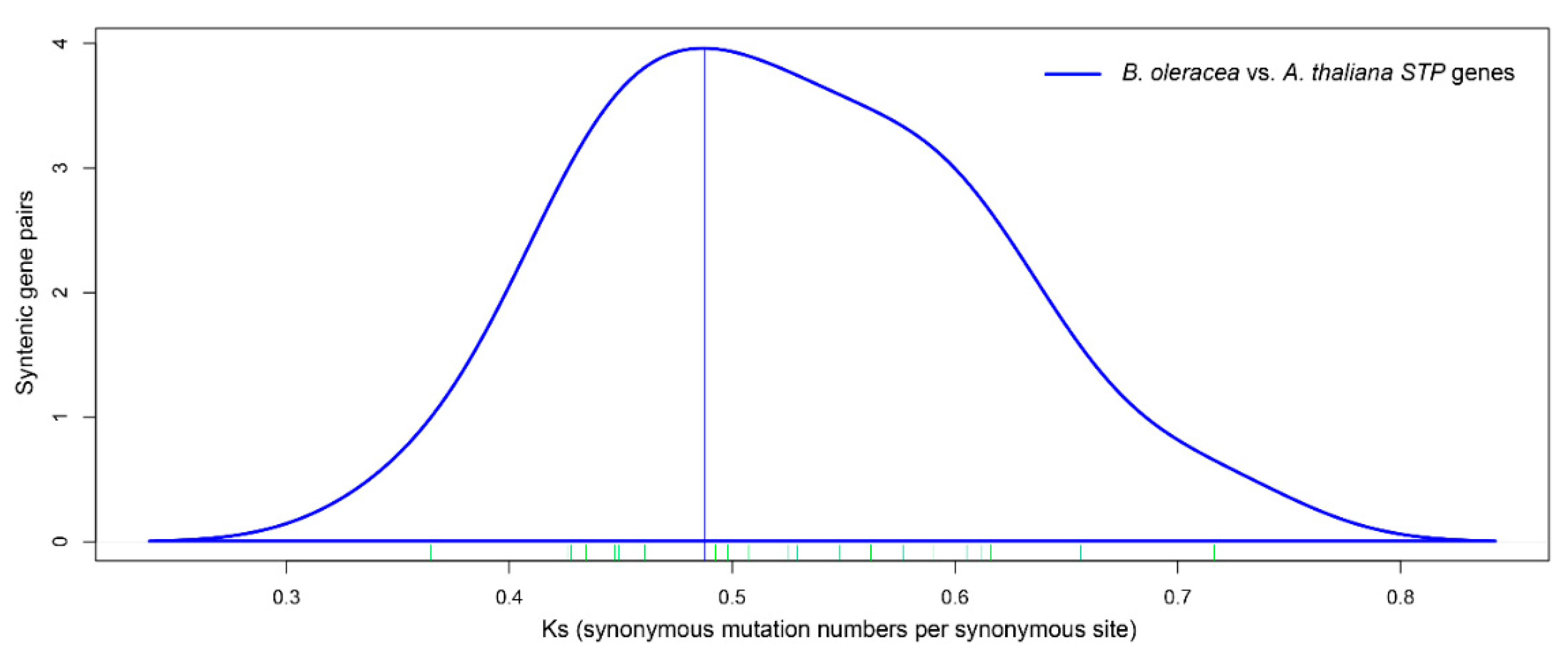
3.3. Ka and Ks Calculation of Orthologous STPs between Brassica oleracea and Arabidopsis thaliana
3.4. Gene Structures, TMHs, and Putative Functional Domains of BoSTP Proteins
3.5. Prediction of cis-Acting Elements of BoSTP Genes
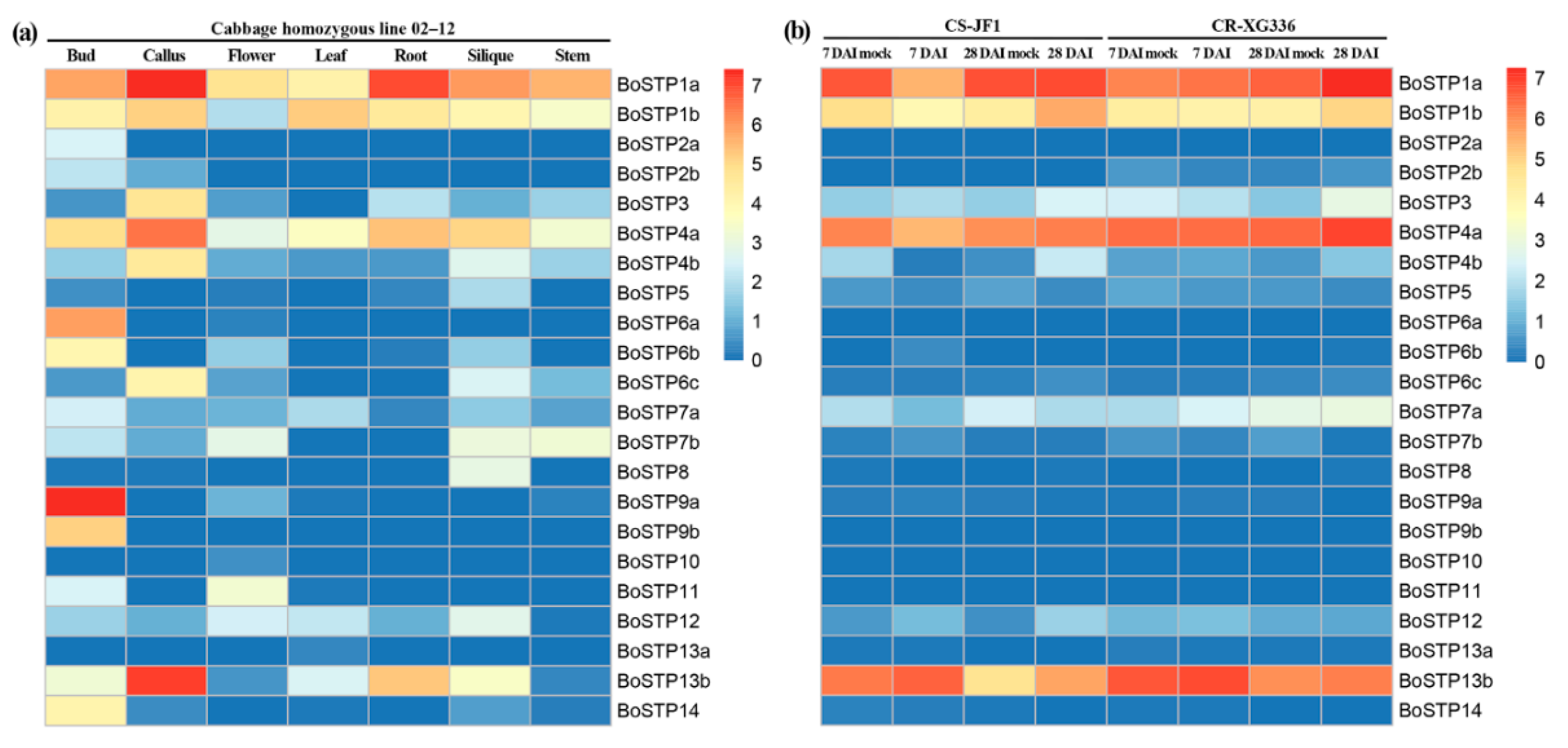
3.6. Expression Profiling of BoSTP Genes in Different Organs and in Response to P. brassicae Infection
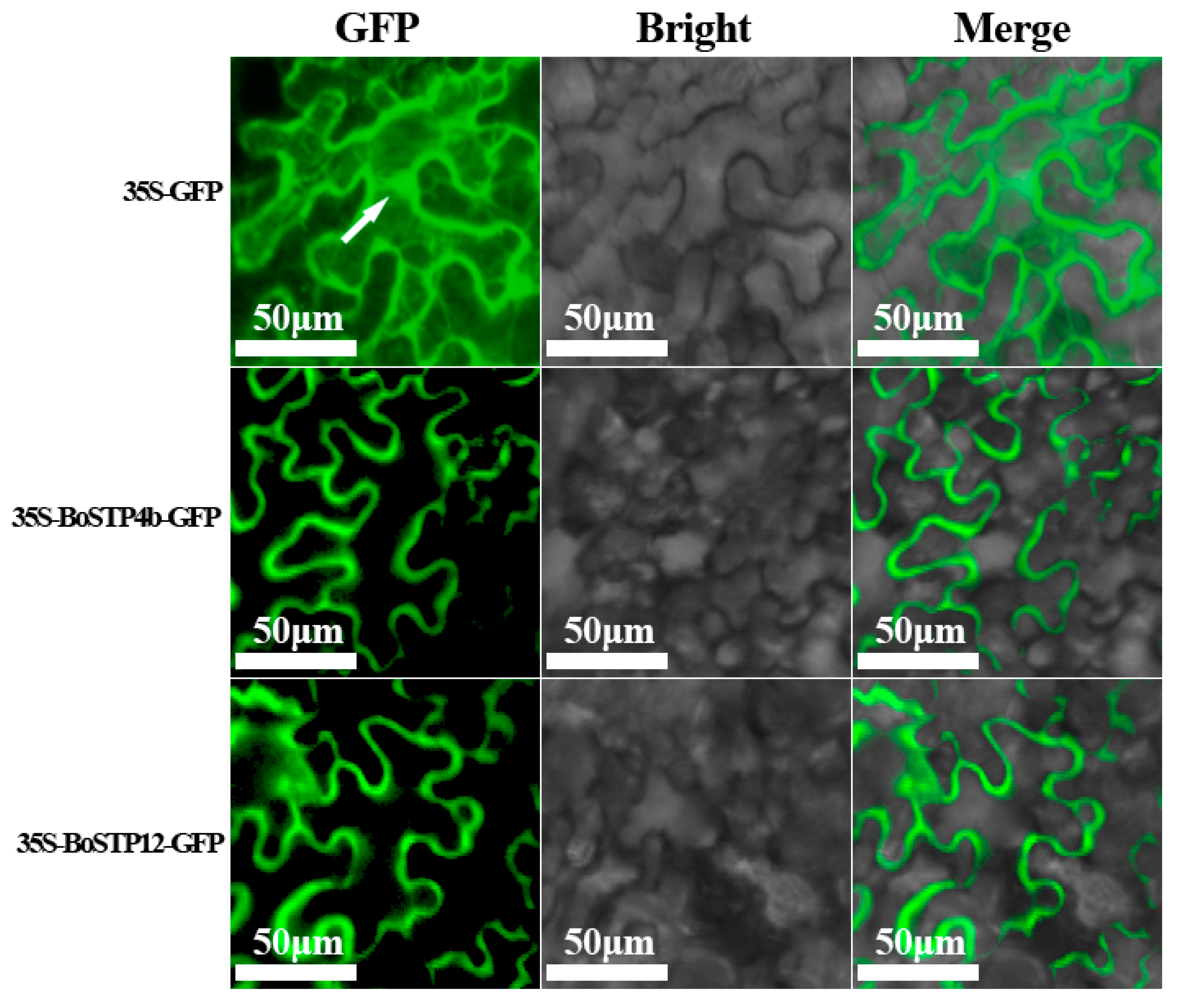
3.7. Subcellular Localization Analysis of BoSTP Proteins in Tobacco
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rolland, F.; Baenagonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, H.A.; Smeekens, S. Sugar sensing and signaling in plants. Front. Plant Sci. 2014, 5, 113. [Google Scholar]
- Ramon, M.; Rolland, F.; Sheen, J. Sugar sensing and signaling. Arab. Book 2008, 6, e0117. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, A.R.; Barrett, M.P.; Bringaud, F.; Gould, G.W. Sugar transporters from bacteria, parasites and mammals: Structure-activity relationships. Trends Biochem. Sci. 1998, 23, 476–481. [Google Scholar] [CrossRef]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dedaldechamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.; Laloi, M.; Coutosthevenot, P.; Maurousset, L. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef] [PubMed]
- Rennie, E.A.; Turgeon, R. A comprehensive picture of phloem loading strategies. Proc. Natl. Acad. Sci. USA 2009, 106, 14162–14167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.Q.; Hou, B.H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.Q.; Guo, W.J.; Kim, J.G.; Underwood, W.; Chaudhuri, B. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef]
- Doidy, J.; Grace, E.; Kuhn, C.; Simonplas, F.; Casieri, L.; Wipf, D. Sugar transporters in plants and in their interactions with fungi. Trends Plant Sci. 2012, 17, 413–422. [Google Scholar] [CrossRef]
- Ludewig, F.; Flügge, U.I. Role of metabolite transporters in source-sink carbon allocation. Front. Plant Sci. 2013, 4, 231. [Google Scholar] [CrossRef]
- Sherson, S.M.; Alford, H.L.; Forbes, S.M.; Wallace, G.; Smith, S.M. Roles of cell-wall invertases and monosaccharide transporters in the growth and development of Arabidopsis. J. Exp. Bot. 2003, 54, 525–531. [Google Scholar] [CrossRef]
- Yan, N. Structural advances for the major facilitator superfamily (MFS) transporters. Trends Biochem. Sci. 2013, 38, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Büttner, M. The Arabidopsis sugar transporter (AtSTP) family: An update. Plant Biol. 2010, 12, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Dang, H.; Chen, Z.; Wu, J.; Chen, Y.; Chen, S.; Luo, L. Genome-wide identification, expression, and functional analysis of the sugar transporter gene family in Cassava (Manihot esculenta). Int. J. Mol. Sci. 2018, 19, 987. [Google Scholar] [CrossRef]
- Afoufabastien, D.; Medici, A.; Jeauffre, J.; Coutosthevenot, P.; Lemoine, R.; Atanassova, R.; Laloi, M. The Vitis vinifera sugar transporter gene family: Phylogenetic overview and macroarray expression profiling. BMC Plant Biol. 2010, 10, 245. [Google Scholar]
- Toyofuku, K.; Kasahara, M.; Yamaguchi, J. Characterization and expression of monosaccharide transporters (OsMSTs) in rice. Plant Cell Physiol. 2000, 41, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Reuscher, S.; Akiyama, M.; Yasuda, T.; Makino, H.; Aoki, K.; Shibata, D.; Shiratake, K. The sugar transporter inventory of tomato: Genome-wide identification and expression analysis. Plant Cell Physiol. 2014, 55, 1123–1141. [Google Scholar] [CrossRef]
- Li, J.; Zheng, D.; Li, L.; Qiao, X.; Wei, S.; Bai, B.; Zhang, S.; Wu, J. Genome-wide function, evolutionary characterization and expression analysis of sugar transporter family genes in pear (Pyrus bretschneideri Rehd). Plant Cell Physiol. 2015, 56, 1721–1737. [Google Scholar] [CrossRef] [PubMed]
- Jiu, S.; Haider, M.S.; Kurjogi, M.M.; Zhang, K.; Zhu, X.; Fang, J. Genome-wide characterization and expression analysis of sugar transporter family genes in woodland strawberry. Plant Genome 2018, 11. [Google Scholar] [CrossRef]
- Büttner, M.; Truernit, E.; Baier, K.; Scholzstarke, J.; Sontheim, M.; Lauterbach, C.; Huss, V.A.R.; Sauer, N. AtSTP3, a green leaf-specific, low affinity monosaccharide-H+ symporter of Arabidopsis thaliana. Plant Cell Environ. 2000, 23, 175–184. [Google Scholar] [CrossRef]
- Büttner, M. The monosaccharide transporter (-like) gene family in Arabidopsis. FEBS Lett. 2007, 581, 2318–2324. [Google Scholar] [CrossRef]
- Sauer, N.; Friedländer, K.; Gräml-Wicke, U. Primary structure, genomic organization and heterologous expression of a glucose transporter from Arabidopsis thaliana. EMBO J. 1990, 9, 3045–3050. [Google Scholar] [CrossRef] [PubMed]
- Stadler, R.; Büttner, M.; Ache, P.; Hedrich, R.; Ivashikina, N.; Melzer, M.; Shearson, S.M.; Smith, S.M.; Sauer, N. Diurnal and light-regulated expression of AtSTP1 in guard cells of Arabidopsis. Plant Physiol. 2003, 133, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Boorer, K.J.; Loo, D.; Wright, E.M. Steady-state and presteady-state kinetics of the H+/hexose cotransporter (STP1) from Arabidopsis thaliana expressed in Xenopus oocytes. J. Biol. Chem. 1994, 269, 20417–20424. [Google Scholar] [PubMed]
- Truernit, E.; Schmid, J.; Epple, P.; Illig, J.; Sauer, N. The sink-specific and stress-regulated Arabidopsis STP4 gene: Enhanced expression of a gene encoding a monosaccharide transporter by wounding, elicitors, and pathogen challenge. Plant Cell 1996, 8, 2169–2182. [Google Scholar] [CrossRef] [PubMed]
- Fotopoulos, V.; Gilbert, M.J.; Pittman, J.K.; Marvier, A.C.; Buchanan, A.J.; Sauer, N.; Hall, J.L.; Williams, L.E. The monosaccharide transporter gene, AtSTP4, and the cell-wall invertase, Atβfruct1, are induced in Arabidopsis during infection with the fungal biotroph Erysiphe cichoracearum. Plant Physiol. 2003, 132, 821–829. [Google Scholar] [CrossRef] [PubMed]
- FAO. Food and Agriculture Organization of the United Nations, Statistics Division. 2017. Available online: http://www.fao.org/faostat/en/#data (accessed on 20 November 2018).
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Ludwigmuller, J.; Prinsen, E.; Rolfe, S.A.; Scholes, J.D. Metabolism and plant hormone action during clubroot disease. J. Plant Growth Regul. 2009, 28, 229–244. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Li, X.L.; Li, H.Z. Research progress in clubroot of crucifers. Plant Prot. 2011, 37, 153–158. (In Chinese) [Google Scholar]
- Dixon, G.R. Clubroot (Plasmodiophora brassicae Woronin)—An agricultural and biological challenge worldwide. Can. J. Plant Pathol. 2014, 36, 5–18. [Google Scholar] [CrossRef]
- Gravot, A.; Deleu, C.; Wagner, G.; Lariagon, C.; Lugan, R.; Todd, C.D.; Wendehenne, D.; Delourme, R.; Bouchereau, A.; Manzanaresdauleux, M.J. Arginase induction represses gall development during clubroot infection in Arabidopsis. Plant Cell Physiol. 2012, 53, 901–911. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, J.; Zhang, S.; Zhang, S.; Li, F.; Strelkov, S.E.; Sun, R.; Hwang, S.F. Resistance to Plasmodiophora brassicae in Brassica rapa and Brassica juncea genotypes from China. Plant Dis. 2014, 99, 776–779. [Google Scholar] [CrossRef]
- Feng, J.; Xiao, Q.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D. Infection of canola by secondary zoospores of Plasmodiophora brassicae produced on a nonhost. Eur. J. Plant Pathol. 2012, 132, 309–315. [Google Scholar] [CrossRef]
- Howard, R.J.; Strelkov, S.E.; Harding, M.W. Clubroot of cruciferous crops-new perspectives on an old disease. Can. J. Plant Pathol. 2010, 32, 43–57. [Google Scholar] [CrossRef]
- Rolfe, S.A.; Strelkov, S.E.; Links, M.G.; Clarke, W.E.; Robinson, S.J.; Djavaheri, M.; Malinowski, R.; Haddadi, P.; Kagale, S.; Parkin, I.A.P. The compact genome of the plant pathogen Plasmodiophora brassicae is adapted to intracellular interactions with host Brassica spp. BMC Genom. 2016, 17, 272. [Google Scholar] [CrossRef] [PubMed]
- Wallenhammar, A.C. Prevalence of Plasmodiophora brassicae in a spring oilseed rape growing area in central Sweden and factors influencing soil infestation levels. Plant Pathol. 1996, 45, 710–719. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.P.; Zhao, M.; Ma, J.; Yu, J.; Huang, S. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.; Bancroft, I.; Cheng, F. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, 29–37. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Wu, J.; Fang, L.; Wang, X. Syntenic gene analysis between Brassica rapa and other Brassicaceae species. Front. Plant Sci. 2012, 3, 198. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.I.; Schein, J.E.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrermata, A.; Sanchezdelbarrio, J.C.; Guiraorico, S.; Librado, P.; Ramosonsins, S.E.; Sanchezgracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Koch, M.A.; Haubold, B.; Mitchellolds, T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and related genera (Brassicaceae). Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef]
- Chou, K.-C.; Shen, H.-B. Cell-PLoc: A package of Web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef]
- Bolger, A.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Hellens, R.P.; Edwards, E.A.; Leyland, N.; Bean, S.J.; Mullineaux, P.M. pGreen: A versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Mol. Biol. 2000, 42, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, I.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Mandakova, T.; Wu, J.; Xie, Q.; Lysak, M.A.; Wang, X. Deciphering the diploid ancestral genome of the mesohexaploid Brassica rapa. Plant Cell 2013, 25, 1541–1554. [Google Scholar] [CrossRef]
- Guo, C.; Guo, R.; Xu, X.; Gao, M.; Li, X.; Song, J.; Zheng, Y.; Wang, X. Evolution and expression analysis of the grape (Vitis vinifera L.) WRKY gene family. J. Exp. Bot. 2014, 65, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Marger, M.D.; Saier, M.H., Jr. A major superfamily of transmembrane facilitators that catalyse uniport, symport and antiport. Trends Biochem. Sci. 1993, 18, 13–20. [Google Scholar] [CrossRef]
- Slewinski, T.L. Diverse functional roles of monosaccharide transporters and their homologs in vascular plants: A physiological perspective. Mol. Plant 2011, 4, 641–662. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]
- Büttner, M.; Sauer, N. Monosaccharide transporters in plants: Structure, function and physiology. Biochim. Biophys. Acta Biomembr. 2000, 1465, 263–274. [Google Scholar] [CrossRef]
- Williams, L.E.; Lemoine, R.; Sauer, N. Sugar transporters in higher plants-a diversity of roles and complex regulation. Trends Plant Sci. 2000, 5, 283–290. [Google Scholar] [CrossRef]
- Walerowski, P.; Gündel, A.; Yahaya, N.; Truman, W.; Sobczak, M.; Olszak, M.; Rolfe, S.A.; Borisjuk, L.; Malinowski, R. Clubroot disease stimulates early steps of phloem differentiation and recruits SWEET sucrose transporters within developing galls. Plant Cell 2018. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.L.; Scholes, J.D. How does clubroot alter the regulation of carbon metabolism in its host? Asp. Appl. Biol. 1995, 42, 125–132. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | tPCKChr | CCB | Arabidopsis thaliana | Brassica oleracea | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Name | Gene ID | ORF (bp) | PL (aa) | Gene Name | Gene ID | Chromosome | Subgenome | ORF (bp) | PL (aa) | MW (kD) | pI | Instability Index | Aliphatic Index | GRAVY | SCLpred | |||
| Clade I | tPCK1 | A | AtSTP1 | AT1G11260 | 1569 | 522 | BoSTP1a | Bol036657 | C05 | LF | 1569 | 522 | 57.60 | 9.22 | 38.45 | 102.87 | 0.49 | cell membrane |
| BoSTP1b | Bol031302 | C08 | MF2 | 1566 | 521 | 57.47 | 9.29 | 39.98 | 102.15 | 0.48 | cell membrane | |||||||
| tPCK7 | X | AtSTP3 | AT5G61520 | 1545 | 514 | BoSTP3 | Bol017006 | C06 | LF | 1512 | 503 | 54.76 | 9.12 | 40.17 | 100.95 | 0.45 | cell membrane | |
| tPCK1 | B | AtSTP5 | AT1G34580 | 1521 | 506 | BoSTP5 | Bol009461 | Scaffold000233 | MF1 | 1521 | 506 | 54.43 | 9.28 | 30.06 | 109.13 | 0.69 | cell membrane | |
| tPCK4 | U | AtSTP12 | AT4G21480 | 1509 | 502 | BoSTP12 | Bol028422 | C01 | LF | 1542 | 513 | 56.62 | 8.70 | 30.38 | 106.57 | 0.56 | cell membrane | |
| Clade II | tPCK2 | F | AtSTP4 | AT3G19930 | 1545 | 514 | BoSTP4a | Bol018147 | C05 | LF | 1545 | 514 | 57.12 | 8.72 | 38.81 | 102.80 | 0.58 | cell membrane |
| BoSTP4b | Bol000550 | Scaffold000531 | MF1 | 1545 | 514 | 56.99 | 8.99 | 38.20 | 102.80 | 0.56 | cell membrane | |||||||
| tPCK1 | C | AtSTP9 | AT1G50310 | 1554 | 517 | BoSTP9a | Bol010165 | C07 | LF | 1551 | 516 | 56.27 | 6.89 | 32.15 | 99.01 | 0.53 | cell membrane | |
| BoSTP9b | Bol014014 | Scaffold000180 | MF2 | 1551 | 516 | 56.15 | 6.89 | 33.77 | 101.67 | 0.54 | cell membrane | |||||||
| tPCK2 | F | AtSTP10 | AT3G19940 | 1545 | 514 | BoSTP10 | Bol000549 | Scaffold000531 | MF1 | 1536 | 511 | 56.16 | 8.60 | 30.51 | 97.50 | 0.48 | cell membrane | |
| tPCK7 | Q | AtSTP11 | AT5G23270 | 1545 | 514 | BoSTP11 | Bol017091 | C06 | LF | 1569 | 522 | 57.17 | 5.79 | 33.58 | 105.50 | 0.55 | cell membrane | |
| Clade III | tPCK1 | A | AtSTP2 | AT1G07340 | 1497 | 498 | BoSTP2a | Bol023380 | C08 | MF1 | 1497 | 498 | 55.48 | 9.27 | 38.43 | 105.52 | 0.50 | cell membrane |
| BoSTP2b | Bol041122 | C05 | LF | 1500 | 499 | 55.42 | 9.29 | 33.25 | 105.51 | 0.52 | cell membrane | |||||||
| tPCK2 | F | AtSTP6 | AT3G05960 | 1524 | 507 | BoSTP6a | Bol002474 | Scaffold000387 | MF1 | 1524 | 507 | 55.86 | 8.76 | 31.67 | 109.98 | 0.65 | cell membrane | |
| BoSTP6b | Bol002866 | Scaffold000372 | LF | 1515 | 504 | 55.59 | 8.29 | 31.06 | 109.50 | 0.65 | cell membrane | |||||||
| BoSTP6c | Bol034076 | Scaffold000040 | MF2 | 1527 | 508 | 55.71 | 8.33 | 30.72 | 112.48 | 0.61 | cell membrane | |||||||
| tPCK7 | Q | AtSTP8 | AT5G26250 | 1524 | 507 | BoSTP8 | Bol022297 | C06 | LF | 1452 | 483 | 53.34 | 8.94 | 32.79 | 112.63 | 0.64 | cell membrane | |
| Clade IV | tPCK5 | O | AtSTP7 | AT4G02050 | 1542 | 513 | BoSTP7a | Bol030687 | C03 | MF1 | 1542 | 513 | 55.76 | 9.03 | 32.99 | 105.89 | 0.47 | cell membrane |
| BoSTP7b | Bol011395 | C09 | LF | 1527 | 508 | 55.09 | 8.83 | 35.14 | 110.18 | 0.53 | cell membrane | |||||||
| tPCK7 | Q | AtSTP13 | AT5G26340 | 1581 | 526 | BoSTP13a | Bol022294 | C06 | LF | 1536 | 511 | 55.95 | 9.31 | 34.63 | 112.94 | 0.59 | cell membrane | |
| BoSTP13b | Bol032462 | C09 | MF2 | 1599 | 532 | 58.04 | 9.30 | 32.58 | 106.65 | 0.49 | cell membrane | |||||||
| tPCK6 | E | AtSTP14 | AT1G77210 | 1515 | 504 | BoSTP14 | Bol027572 | C07 | LF | 1038 | 345 | 38.33 | 8.47 | 45.45 | 112.46 | 0.51 | cell membrane | |
| Orthologous Gene Pairs | Ks | Ka | Ka/Ks | Duplication Date (MYA) | |
|---|---|---|---|---|---|
| AtSTP1 | BoSTP1a | 0.4982 | 0.0193 | 0.0387 | 16.6 |
| BoSTP1b | 0.4609 | 0.0189 | 0.0410 | 15.4 | |
| AtSTP2 | BoSTP2a | 0.6117 | 0.0981 | 0.1604 | 20.4 |
| BoSTP2b | 0.6566 | 0.1081 | 0.1646 | 21.9 | |
| AtSTP3 | BoSTP3 | 0.4924 | 0.0737 | 0.1497 | 16.4 |
| AtSTP4 | BoSTP4a | 0.4260 | 0.0261 | 0.0613 | 14.2 |
| BoSTP4b | 0.4345 | 0.0336 | 0.0773 | 14.5 | |
| AtSTP5 | BoSTP5 | 0.6055 | 0.0930 | 0.1536 | 20.2 |
| AtSTP6 | BoSTP6a | 0.5253 | 0.0452 | 0.0860 | 17.5 |
| BoSTP6b | 0.5766 | 0.0408 | 0.0708 | 19.2 | |
| BoSTP6c | 0.5485 | 0.0560 | 0.1021 | 18.3 | |
| AtSTP7 | BoSTP7a | 0.5290 | 0.0360 | 0.0681 | 17.6 |
| BoSTP7b | 0.7164 | 0.0476 | 0.0664 | 23.9 | |
| AtSTP8 | BoSTP8 | 0.5900 | 0.0573 | 0.0971 | 19.7 |
| AtSTP9 | BoSTP9a | 0.4489 | 0.0348 | 0.0775 | 15.0 |
| BoSTP9b | 0.4279 | 0.0392 | 0.0916 | 14.3 | |
| AtSTP10 | BoSTP10 | 0.3646 | 0.0653 | 0.1791 | 12.2 |
| AtSTP11 | BoSTP11 | 0.6162 | 0.1001 | 0.1624 | 20.5 |
| AtSTP12 | BoSTP12 | 0.5073 | 0.0500 | 0.0986 | 16.9 |
| AtSTP13 | BoSTP13a | 0.4475 | 0.0574 | 0.1283 | 14.9 |
| BoSTP13b | 0.4877 | 0.0203 | 0.0416 | 16.3 | |
| AtSTP14 | BoSTP14 | 0.5623 | 0.0300 | 0.0534 | 18.7 |
| Elements | ABRE | ARE | CGTCA-Motif | ERE | G-Box | GT1-Motif | LTR | MBS | MRE | P-Box | STRE | TATC-Box | TC-Rich Repeats | TCA-Element | TGA-Element | TGACG-Motif | WUN-Motif | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ABA | Anaerobic | MeJA | Ethylene | Light | Light | Low Temperature | Drought | Light | Gibberellin | Stress Response | Gibberellin | Defense & Stress | Salicylic Acid | Auxin | MeJA | Wound | ||
| BoSTP1a | 2 | 3 | 2 | 2 | 1 | 1 | 1 | 12 | ||||||||||
| BoSTP1b | 2 | 4 | 1 | 3 | 1 | 1 | 1 | 13 | ||||||||||
| BoSTP2a | 2 | 2 | ||||||||||||||||
| BoSTP2b | 2 | 1 | 2 | 5 | ||||||||||||||
| BoSTP3 | 1 | 2 | 1 | 4 | ||||||||||||||
| BoSTP4a | 1 | 1 | 1 | 3 | ||||||||||||||
| BoSTP4b | 3 | 1 | 1 | 2 | 7 | |||||||||||||
| BoSTP5 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 9 | |||||||||
| BoSTP6a | 1 | 1 | 2 | |||||||||||||||
| BoSTP6b | 1 | 1 | 2 | 3 | 3 | 1 | 11 | |||||||||||
| BoSTP6c | 2 | 3 | 1 | 2 | 1 | 1 | 3 | 13 | ||||||||||
| BoSTP7a | 2 | 2 | 4 | 3 | 1 | 1 | 1 | 1 | 2 | 1 | 18 | |||||||
| BoSTP7b | 4 | 2 | 1 | 6 | 1 | 1 | 1 | 2 | 1 | 19 | ||||||||
| BoSTP8 | 3 | 2 | 1 | 1 | 3 | 2 | 1 | 13 | ||||||||||
| BoSTP9a | 2 | 3 | 1 | 4 | 2 | 1 | 2 | 2 | 2 | 1 | 20 | |||||||
| BoSTP9b | 5 | 1 | 3 | 2 | 1 | 1 | 2 | 1 | 16 | |||||||||
| BoSTP10 | 2 | 3 | 2 | 2 | 4 | 1 | 14 | |||||||||||
| BoSTP11 | 1 | 1 | 2 | 1 | 2 | 1 | 1 | 1 | 1 | 11 | ||||||||
| BoSTP12 | 2 | 1 | 2 | 2 | 1 | 1 | 9 | |||||||||||
| BoSTP13a | 5 | 2 | 1 | 2 | 6 | 1 | 1 | 1 | 1 | 1 | 1 | 22 | ||||||
| BoSTP13b | 3 | 2 | 2 | 1 | 4 | 2 | 1 | 1 | 2 | 1 | 19 | |||||||
| BoSTP14 | 3 | 1 | 1 | 1 | 3 | 1 | 1 | 11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Wang, S.; Yu, F.; Tang, J.; Yu, L.; Wang, H.; Li, J. Genome-Wide Identification and Expression Profiling of Sugar Transporter Protein (STP) Family Genes in Cabbage (Brassica oleracea var. capitata L.) Reveals their Involvement in Clubroot Disease Responses. Genes 2019, 10, 71. https://doi.org/10.3390/genes10010071
Zhang W, Wang S, Yu F, Tang J, Yu L, Wang H, Li J. Genome-Wide Identification and Expression Profiling of Sugar Transporter Protein (STP) Family Genes in Cabbage (Brassica oleracea var. capitata L.) Reveals their Involvement in Clubroot Disease Responses. Genes. 2019; 10(1):71. https://doi.org/10.3390/genes10010071
Chicago/Turabian StyleZhang, Wei, Shenyun Wang, Fangwei Yu, Jun Tang, Li Yu, Hong Wang, and Jianbin Li. 2019. "Genome-Wide Identification and Expression Profiling of Sugar Transporter Protein (STP) Family Genes in Cabbage (Brassica oleracea var. capitata L.) Reveals their Involvement in Clubroot Disease Responses" Genes 10, no. 1: 71. https://doi.org/10.3390/genes10010071
APA StyleZhang, W., Wang, S., Yu, F., Tang, J., Yu, L., Wang, H., & Li, J. (2019). Genome-Wide Identification and Expression Profiling of Sugar Transporter Protein (STP) Family Genes in Cabbage (Brassica oleracea var. capitata L.) Reveals their Involvement in Clubroot Disease Responses. Genes, 10(1), 71. https://doi.org/10.3390/genes10010071