Non-Syndromic Cleft Lip with or without Cleft Palate: Genome-Wide Association Study in Europeans Identifies a Suggestive Risk Locus at 16p12.1 and Supports SH3PXD2A as a Clefting Susceptibility Gene

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. GWAS Patients
2.3. GWAS Controls
2.4. GWAS Genotyping and Quality Control (QC)
2.5. Imputation of GWAS Data
2.6. Genome-Wide Association Analysis
2.7. Replication Analysis for Two Interesting SNVs
2.8. Prioritization of Candidate Genes
2.9. Expression Analysis using SysFACE
3. Results
3.1. SNV-Based Analysis
3.2. Replication of Previously Reported nsCL/P Susceptibility Loci
3.3. Gene-Based Evaluation and Gene-Set Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mangold, E.; Ludwig, K.U.; Nöthen, M.M. Breakthroughs in the genetics of orofacial clefting. Trends Mol. Med. 2011, 17, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Grosen, D.; Bille, C.; Petersen, I.; Skytthe, A.; von Hjelmborg, J.B.; Pedersen, J.K.; Murray, J.C.; Christensen, K. Risk of oral clefts in twins. Epidemiology 2011, 22, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Rahimov, F.; Marazita, M.L.; Visel, A.; Cooper, M.E.; Hitchler, M.J.; Rubini, M.; Domann, F.E.; Govil, M.; Christensen, K.; Bille, C.; et al. Disruption of an AP-2alpha binding site in an IRF6 enhancer is associated with cleft lip. Nat. Genet. 2008, 40, 1341–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, L.M.; Mansilla, M.A.; Bullard, S.A.; Cooper, M.E.; Busch, T.D.; Machida, J.; Johnson, M.K.; Brauer, D.; Krahn, K.; Daack-Hirsch, S.; et al. FOXE1 association with both isolated cleft lip with or without cleft palate, and isolated cleft palate. Hum. Mol. Genet. 2009, 18, 4879–4896. [Google Scholar] [CrossRef] [Green Version]
- Bureau, A.; Parker, M.M.; Ruczinski, I.; Taub, M.A.; Marazita, M.L.; Murray, J.C.; Mangold, E.; Noethen, M.M.; Ludwig, K.U.; Hetmanski, J.B.; et al. Whole Exome sequencing of distant relatives in multiplex families implicates rare variants in candidate genes for oral clefts. Genetics 2014, 197, 1039–1044. [Google Scholar] [CrossRef] [Green Version]
- Cox, L.L.; Cox, T.C.; Uribe, L.M.M.; Zhu, Y.; Richter, C.T.; Nidey, N.; Standley, J.M.; Deng, M.; Blue, E.; Chong, J.X.; et al. Mutations in the Epithelial Cadherin-p120-Catenin Complex Cause Mendelian Non-Syndromic Cleft Lip with or without Cleft Palate. Am. J. Hum. Genet. 2018, 102, 1143–1157. [Google Scholar] [CrossRef] [Green Version]
- Cox, T.C.; Lidral, A.C.; McCoy, J.C.; Liu, H.; Cox, L.L.; Zhu, Y.; Anderson, R.D.; Uribe, L.M.M.; Anand, D.; Deng, M.; et al. Mutations in GDF11 and the extracellular antagonist, Follistatin, as a likely cause of Mendelian forms of orofacial clefting in humans. Hum. Mutat. 2019, 40, 1813–1825. [Google Scholar] [CrossRef]
- Birnbaum, S.; Ludwig, K.U.; Reutter, H.; Herms, S.; Steffens, M.; Rubini, M.; Baluardo, C.; Ferrian, M.; de Assis, N.A.; Alblas, M.; et al. Key susceptibility locus for nonsyndromic cleft lip with or without cleft palate on chromosome 8q24. Nat. Genet. 2009, 41, 473–477. [Google Scholar] [CrossRef]
- Grant, S.F.A.; Wang, K.; Zhang, H.; Glaberson, W.; Annaiah, K.; Kim, C.E.; Bradfield, J.P.; Glessner, J.T.; Thomas, K.A.; Garris, M.; et al. A genome-wide association study identifies a locus for nonsyndromic cleft lip with or without cleft palate on 8q24. J. Pediatr. 2009, 155, 909–913. [Google Scholar] [CrossRef]
- Mangold, E.; Ludwig, K.U.; Birnbaum, S.; Baluardo, C.; Ferrian, M.; Herms, S.; Reutter, H.; de Assis, N.A.; Chawa, T.A.; Mattheisen, M.; et al. Genome-wide association study identifies two susceptibility loci for nonsyndromic cleft lip with or without cleft palate. Nat. Genet. 2010, 42, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Beaty, T.H.; Murray, J.C.; Marazita, M.L.; Munger, R.G.; Ruczinski, I.; Hetmanski, J.B.; Liang, K.Y.; Wu, T.; Murray, T.; Fallin, M.D.; et al. A genome-wide association study of cleft lip with and without cleft palate identifies risk variants near MAFB and ABCA4. Nat. Genet. 2010, 42, 525–529. [Google Scholar] [CrossRef]
- Ludwig, K.U.; Mangold, E.; Herms, S.; Nowak, S.; Paul, A.; Becker, J.; Herberz, R.; Alchawa, T.; Böhmer, A.C.; Mattheisen, M.; et al. Genome-wide meta-analyses of nonsyndromic cleft lip with or without cleft palate indentify six new risk loci. Nat Genet. 2012, 44, 968–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaty, T.H.; Taub, M.A.; Scott, A.F.; Murray, J.C.; Marazita, M.L.; Schwender, H.; Parker, M.M.; Hetmanski, J.B.; Balakrishnan, P.; Mansilla, M.A.; et al. Confirming genes influencing risk to cleft lip with/without cleft palate in a case-parent trio study. Hum. Genet. 2013, 132, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Huang, Y.; Yin, A.; Pan, Y.; Wang, Y.; Wang, C.; Du, Y.; Wang, M.; Lan, F.; Hu, Z.; et al. Genome-wide association study identifies a new susceptibility locus for cleft lip with or without a cleft palate. Nat. Commun. 2015, 6, 6414. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, K.U.; Ahmed, S.T.; Böhmer, A.C.; Sangani, N.B.; Varghese, S.; Klamt, J.; Schuenke, H.; Gültepe, P.; Hofmann, A.; Rubini, M.; et al. Meta-analysis Reveals Genome-Wide Significance at 15q13 for Nonsyndromic Clefting of Both the Lip and the Palate, and Functional Analyses Implicate GREM1 As a Plausible Causative Gene. PLoS Genet. 2016, 12, e1005914. [Google Scholar] [CrossRef] [Green Version]
- Leslie, E.J.; Carlson, J.C.; Shaffer, J.R.; Feingold, E.; Wehby, G.; Laurie, C.A.; Jain, D.; Laurie, C.C.; Doheny, K.F.; McHenry, T.; et al. A multi-ethnic genome-wide association study identifies novel loci for non-syndromic cleft lip with or without cleft palate on 2p24.2, 17q23 and 19q13. Hum. Mol. Genet. 2016, 25, 2862–2872. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Zuo, X.; He, M.; Gao, J.; Fu, Y.; Qin, C.; Meng, L.; Wang, W.; Song, Y.; Cheng, Y.; et al. Genome-wide analyses of non-syndromic cleft lip with palate identify 14 novel loci and genetic heterogeneity. Nat. Commun. 2017, 8, 14364. [Google Scholar] [CrossRef]
- Mostowska, A.; Gaczkowska, A.; Żukowski, K.; Ludwig, K.U.; Hozyasz, K.K.; Wójcicki, P.; Mangold, E.; Böhmer, A.C.; Heilmann-Heimbach, S.; Knapp, M.; et al. Common variants in DLG1 locus are associated with non-syndromic cleft lip with or without cleft palate. Clin. Genet. 2018, 93, 784–793. [Google Scholar] [CrossRef]
- Ludwig, K.U.; Böhmer, A.C.; Bowes, J.; Nikolic, M.; Ishorst, N.; Wyatt, N.; Hammond, N.L.; Gölz, L.; Thieme, F.; Barth, S.; et al. Imputation of orofacial clefting data identifies novel risk loci and sheds light on the genetic background of cleft lip ± cleft palate and cleft palate only. Hum. Mol. Genet. 2017, 26, 829–842. [Google Scholar] [CrossRef]
- Leslie, E.J.; Carlson, J.C.; Shaffer, J.R.; Butali, A.; Buxó, C.J.; Castilla, E.E.; Christensen, K.; Deleyiannis, F.W.B.; Leigh Field, L.; Hecht, J.T.; et al. Genome-wide meta-analyses of nonsyndromic orofacial clefts identify novel associations between FOXE1 and all orofacial clefts, and TP63 and cleft lip with or without cleft palate. Hum. Genet. 2017, 136, 275–286. [Google Scholar] [CrossRef] [Green Version]
- van Rooij, I.A.L.M.; van der Zanden, L.F.M.; Bongers, E.M.H.F.; Renkema, K.Y.; Wijers, C.H.W.; Thonissen, M.; Dokter, E.M.J.; Marcelis, C.L.M.; de Blaauw, I.; Wijnen, M.H.W.A.; et al. AGORA, a data- and biobank for birth defects and childhood cancer. Birth Defects Res. Part A-Clin. Mol. Teratol. 2016, 106, 675–684. [Google Scholar] [CrossRef]
- Galesloot, T.E.; Vermeulen, S.H.; Swinkels, D.W.; de Vegt, F.; Franke, B.; den Heijer, M.; de Graaf, J.; Verbeek, A.L.M.; Kiemeney, L.A.L.M. Cohort Profile: The Nijmegen Biomedical Study (NBS). Int. J. Epidemiol. 2017, 46, 1099–1100. [Google Scholar] [CrossRef] [Green Version]
- Manichaikul, A.; Mychaleckyj, J.C.; Rich, S.S.; Daly, K.; Sale, M.; Chen, W.M. Robust relationship inference in genome-wide association studies. Bioinformatics 2010, 26, 2867–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howie, B.; Fuchsberger, C.; Stephens, M.; Marchini, J.; Abecasis, G.R. Fast and accurate genotype imputation in genome-wide association studies through pre-phasing. Nat. Genet. 2012, 44, 955–959. [Google Scholar] [CrossRef]
- Marchini, J.; Howie, B. Genotype imputation for genome-wide association studies. Nat. Rev. Genet. 2010, 11, 499–511. [Google Scholar] [CrossRef]
- Rojas-Martinez, A.; Reutter, H.; Chacon-Camacho, O.; Leon-Cachon, R.B.R.; Munoz-Jimenez, S.G.; Nowak, S.; Becker, J.; Herberz, R.; Ludwig, K.U.; Paredes-Zenteno, M.; et al. Genetic risk factors for nonsyndromic cleft lip with or without cleft palate in a Mesoamerican population: Evidence for IRF6 and variants at 8q24 and 10q25. Birth Defects Res. A. Clin. Mol. Teratol. 2010, 88, 535–537. [Google Scholar] [CrossRef]
- Aldhorae, K.A.; Böhmer, A.C.; Ludwig, K.U.; Esmail, A.H.A.; Al-Hebshi, N.N.; Lippke, B.; Gölz, L.; Nöthen, M.M.; Daratsianos, N.; Knapp, M.; et al. Nonsyndromic cleft lip with or without cleft palate in Arab populations: Genetic analysis of 15 risk loci in a novel case-control sample recruited in Yemen. Birth Defects Res. Part A - Clin. Mol. Teratol. 2014, 100, 307–313. [Google Scholar] [CrossRef]
- Watanabe, K.; Taskesen, E.; Van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef] [Green Version]
- de Leeuw, C.A.; Mooij, J.M.; Heskes, T.; Posthuma, D. MAGMA: Generalized Gene-Set Analysis of GWAS Data. PLoS Comput. Biol. 2015, 11, e1004219. [Google Scholar] [CrossRef]
- Leslie, E.J.; Koboldt, D.C.; Kang, C.J.; Ma, L.; Hecht, J.T.; Wehby, G.L.; Christensen, K.; Czeizel, A.E.; Deleyiannis, F.W.-B.; Fulton, R.S.; et al. IRF6 mutation screening in non-syndromic orofacial clefting: Analysis of 1521 families. Clin. Genet. 2016, 90, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Butali, A.; Mossey, P.; Adeyemo, W.; Eshete, M.; Gaines, L.; Braimah, R.; Aregbesola, B.; Rigdon, J.; Emeka, C.; Olutayo, J.; et al. Rare functional variants in genome-wide association identified candidate genes for nonsyndromic clefts in the African population. Am. J. Med. Genet. Part A 2014, 164, 2567–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowans, L.J.J.; Adeyemo, W.L.; Eshete, M.; Mossey, P.A.; Busch, T.; Aregbesola, B.; Donkor, P.; Arthur, F.K.N.; Bello, S.A.; Martinez, A.; et al. Association studies and direct DNA sequencing implicate genetic susceptibility loci in the etiology of nonsyndromic orofacial clefts in sub-Saharan African populations. J. Dent. Res. 2016, 95, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Jugessur, A.; Farlie, P.G.; Kilpatrick, N. The genetics of isolated orofacial clefts: From genotypes to subphenotypes. Oral Dis. 2009, 15, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Wilderman, A.; VanOudenhove, J.; Kron, J.; Noonan, J.P.; Cotney, J. High-Resolution Epigenomic Atlas of Human Embryonic Craniofacial Development. Cell Rep. 2018, 23, 1581–1597. [Google Scholar] [CrossRef]
- Firth, H.V.; Richards, S.M.; Bevan, A.P.; Clayton, S.; Corpas, M.; Rajan, D.; Van Vooren, S.; Moreau, Y.; Pettett, R.M.; Carter, N.P. DECIPHER: Database of Chromosomal Imbalance and Phenotype in Humans Using Ensembl Resources. Am. J. Hum. Genet. 2009, 84, 524–533. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Jia, P.; Wolfinger, R.D.; Chen, X.; Zhao, Z. Gene set analysis of genome-wide association studies: Methodological issues and perspectives. Genomics 2011, 98, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Seals, D.F.; Azucena, E.F.; Pass, I.; Tesfay, L.; Gordon, R.; Woodrow, M.; Resau, J.H.; Courtneidge, S.A. The adaptor protein Tks5/Fish is required for podosome formation and function, and for the protease-driven invasion of cancer cells. Cancer Cell 2005, 7, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Murphy, D.A.; Diaz, B.; Bromann, P.A.; Tsai, J.H.; Kawakami, Y.; Maurer, J.; Stewart, R.A.; Izpisúa-Belmonte, J.C.; Courtneidge, S.A. A Src-Tks5 pathway is required for neural crest cell migration during embryonic development. PLoS ONE 2011, 6, e22499. [Google Scholar] [CrossRef] [Green Version]
- Cejudo-Martin, P.; Yuen, A.; Vlahovich, N.; Lock, P.; Courtneidge, S.A.; Díaz, B. Genetic disruption of the Sh3pxd2a gene reveals an essential role in mouse development and the existence of a novel isoform of Tks5. PLoS ONE 2014, 9, e107674. [Google Scholar] [CrossRef] [Green Version]
- Picollo, A.; Malvezzi, M.; Accardi, A. TMEM16 proteins: Unknown structure and confusing functions. J. Mol. Biol. 2015, 427, 94–105. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
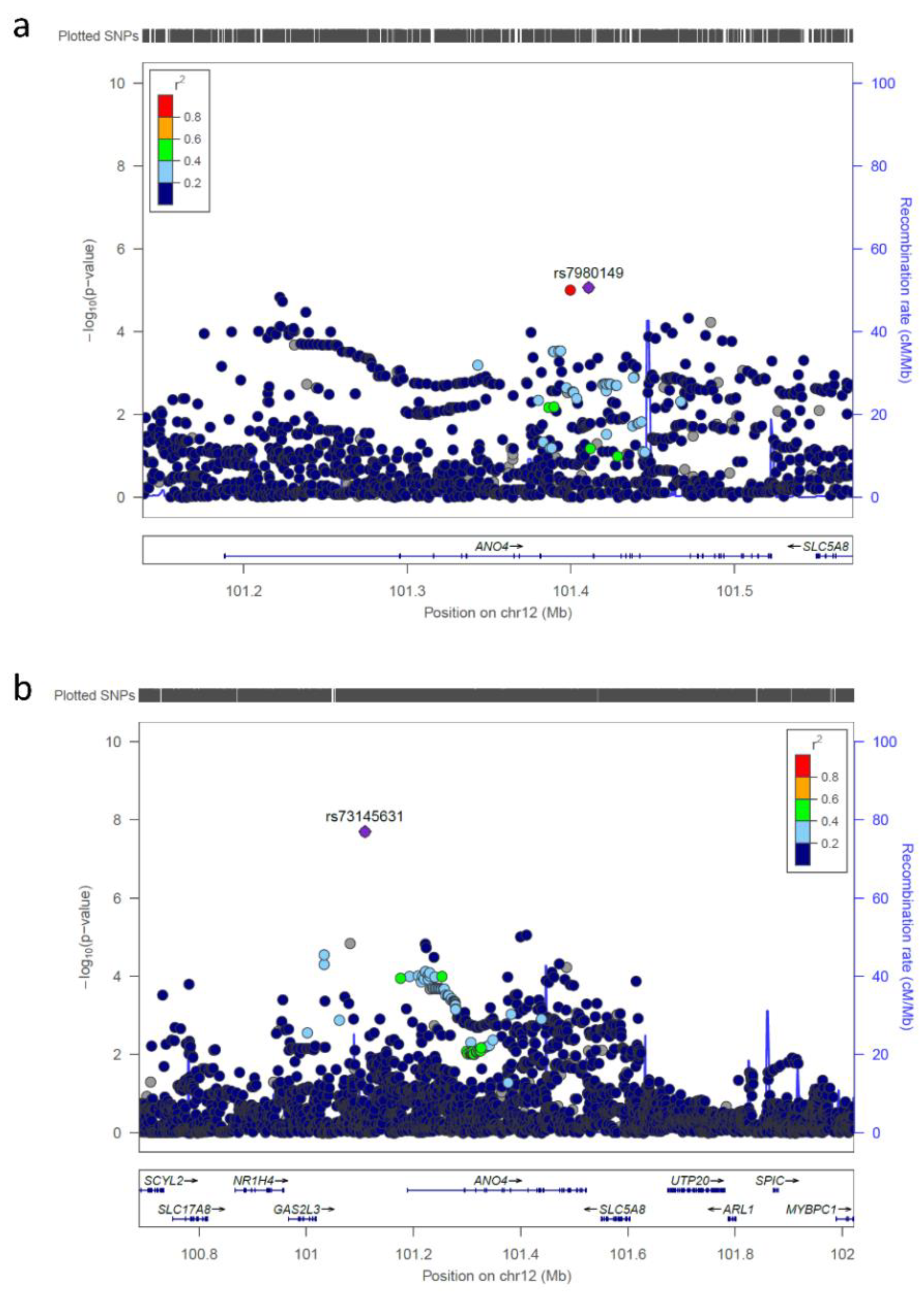{kind=link}
| LeadSNV_ID 1 | Chromosome | Pos (hg19) | Alleles 2 | Case Frequency A | Control Frequency A | All OR | p-Value 3 |
|---|---|---|---|---|---|---|---|
| rs36068947 | 1 | 19003293 | G/GC | 0.592 | 0.702 | 1.62 | 1.06 × 10-6 |
| rs11247713 | 1 | 28230494 | A/G | 0.165 | 0.261 | 1.78 | 2.54 × 10-6 |
| rs12404189 | 1 | 81851687 | T/C | 0.951 | 0.899 | 0.46 | 4.48 × 10-6 |
| rs145794647 | 1 | 213880572 | A/G | 0.915 | 0.964 | 2.46 | 3.17 × 10-6 |
| rs1266381 | 1 | 236681990 | A/G | 0.68 | 0.781 | 1.68 | 1.49 × 10-6 |
| rs13429389 | 2 | 2564600 | G/A | 0.713 | 0.807 | 1.68 | 6.34 × 10-6 |
| rs1431903 | 2 | 168468315 | T/C | 0.113 | 0.192 | 1.86 | 8.76 × 10-6 |
| rs112762347 | 5 | 13103319 | G/A | 0.952 | 0.986 | 3.56 | 5.80 × 10-6 |
| rs4868099 | 5 | 170982499 | T/C | 0.156 | 0.089 | 0.53 | 4.29 × 10-6 |
| rs141109174 | 7 | 2926872 | G/GA | 0.924 | 0.856 | 0.49 | 3.95 × 10-6 |
| rs987525 | 8 | 129946154 | C/A | 0.61 | 0.748 | 1.89 | 8.73 × 10-11 |
| rs1535462 | 10 | 102973872 | A/G | 0.623 | 0.518 | 0.65 | 5.69 × 10-6 |
| rs4918052 | 10 | 105555131 | G/A | 0.303 | 0.203 | 0.58 | 2.34 × 10-6 |
| rs17770307 | 10 | 115259535 | G/C | 0.984 | 0.942 | 0.26 | 2.59 × 10-6 |
| rs148248623 | 12 | 23265077 | CA/C | 0.907 | 0.961 | 2.52 | 1.38 × 10-6 |
| rs7980090 | 12 | 67951884 | C/A | 0.908 | 0.958 | 2.28 | 2.35 × 10-6 |
| rs73145631 | 12 | 101109530 | G/A | 0.985 | 0.936 | 0.23 | 1.99 × 10-8 |
| rs56814511 | 12 | 125789014 | C/T | 0.608 | 0.705 | 1.55 | 9.01 × 10-6 |
| rs184467 | 13 | 29622636 | G/T | 0.428 | 0.316 | 0.62 | 1.53 × 10-6 |
| rs10520788 | 15 | 96126414 | T/C | 0.973 | 0.924 | 0.34 | 6.19 × 10-6 |
| rs56383345 | 16 | 26344915 | G/C | 0.922 | 0.845 | 0.46 | 4.17 × 10-7 |
| rs11640952 | 16 | 78093932 | G/T | 0.693 | 0.587 | 0.63 | 6.22 × 10-6 |
| rs7215555 | 17 | 29564603 | G/A | 0.231 | 0.331 | 1.64 | 3.28 × 10-6 |
| rs61296704 | 17 | 75721588 | G/A | 0.932 | 0.972 | 2.55 | 8.22 × 10-6 |
| rs73512449 | 19 | 18123050 | G/C | 0.888 | 0.946 | 2.19 | 7.00 × 10-6 |
| SNV-ID | p-Values after Genotyping of SNV in Respective Replication Sample | ||||||
|---|---|---|---|---|---|---|---|
| SNV-ID | Chr. | Pos. (hg19) | Bonn 1 | Mexico 2 | Yemen 3 | All 4 | OR (95% CI) |
| rs73145631 | 12q23.1 | 101109530 | 0.544 | n.a.* | 0.714 | 0.488 | 1.18 (0.74–1.90) |
| rs56383345 | 16p12.1 | 26344915 | 0.027 | 0.577 ** | 0.0099 | 0.00167 | 1.53 (1.17–1.98) |
| Literature Risk Locus 1 | Literature Lead SNV at Respective Locus | Better Dutch/Belgian GWAS lead SNV at Respective Locus | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Locus | Original Study | SNV | Pos. (hg19) | p-Value 2 | OR | SNV 3 | Pos (hg19) | p-Value 2,3 | OR |
| 1p36 | Ludwig et al. 2012 [12] | rs742071 | 18979874 | 2.11 × 10-5 | 1.54 | rs36068947 | 19003293 | 1.06272 × 10-6 | 1.62 |
| 1p22 | Beaty et al. 2010 [11] | rs560426 | 94553438 | 0.299446 | 0.90 | rs952499 | 94558425 | 0.0616217 | 0.83 |
| 1q32.1 | Rahimov et al. 2008 [3] | rs642961 | 209989270 | 0.00055997 | 0.69 | - | - | - | - |
| 2p25.1 | Yu et al. 2017 [17] | rs287982 | 9972442 | 0.898877 | 1.00 | - | - | - | - |
| 2p24.2 | Leslie et al. 2016 [16] | rs7552 | 16733928 | 4.84 × 10-5 | 1.52 | rs62122693 | 16734878 | 3.10168 × 10-5 | 1.56 |
| 2p21PKDCC | Ludwig et al. 2017 [19] | rs6740960 | 42181679 | 0.00745325 | 1.28 | rs17029056 | 42158304 | 0.00011259 | 1.52 |
| 2p21THADA | Ludwig et al. 2012 [12] | rs7590268 | 43540125 | 0.13198 | 1.20 | rs6544652 | 43626212 | 0.0881938 | 1.23 |
| 3p11.1 | Ludwig et al. 2012 [12] | rs7632427 | 89534377 | 0.0342897 | 0.81 | rs3792572 | 89456555 | 0.0101378 | 0.77 |
| 3q12.1 | Beaty et al. 2013 [13] | rs793464 | 99626028 | 8.21 × 10-5 | 0.70 | rs9832134 | 99836722 | 4.67043 × 10-5 | 0.66 |
| 3q28 | Leslie et al. 2017 [20] | rs76479869 | 189553372 | 0.00247675 | 1.77 | rs17447439 | 189549423 | 5.67543 × 10-5 | 2.26 |
| 3q29 | Mostowska et al. 2018 [18] | rs338217 | 197026927 | 0.0986085 | 0.85 | rs34099552 | 196799735 | 0.0228988 | 1.27 |
| 4p16.2 | Yu et al. 2017 [17] | rs1907989 | 4818925 | 0.751776 | 1.05 | rs10937893 | 4810491 | 0.649752 | 0.94 |
| 4q28.1 | Yu et al. 2017 [17] | rs908822 | 124906257 | 0.657599 | 0.89 | rs76837304 | 124868111 | 0.532867 | 0.86 |
| 5p12 | Yu et al. 2017 [17] | rs10462065 | 44068846 | 0.258344 | 1.18 | rs139738798 | 44183419 | 0.109377 | 1.26 |
| 8p11.23 | Yu et al. 2017 [17] | rs13317 | 38269514 | 0.774445 | 0.97 | rs75168396 | 38014429 | 0.3038 | 1.11 |
| 8q21 | Ludwig et al. 2012 [12] | rs12543318 | 88868340 | 0.00118317 | 0.73 | - | - | - | - |
| 8q22.1 | Yu et al. 2017 [17] | rs957448 | 95541302 | 0.560023 | 0.94 | rs4442106 | 95609488 | 0.0626976 | 0.83 |
| 8q24 | Birnbaum et al. 2009 [8] | rs987525 | 129946154 | 8.73 × 10-11 | 1.89 | - | - | - | - |
| 9q22.2 | Yu et al. 2017 [17] | rs10908902 | 92224825 | 0.0178356 | 1.31 | rs2031970 | 92204172 | 0.00252225 | 1.41 |
| 9q22.32 | Yu et al. 2017 [17] | rs10512248 | 98259703 | 0.0938744 | 0.84 | rs28591501 | 98278644 | 0.0696448 | 0.82 |
| 9q21.33 | Moreno et al. 2009 [4] | rs3758249 | 100614140 | 0.00519618 | 1.32 | rs7033765 | 100591705 | 0.00127545 | 1.38 |
| 10q25 | Mangold et al. 2010 [10] | rs7078160 | 118827560 | 0.00019291 | 1.60 | rs5788208 | 118836076 | 0.000106433 | 1.63 |
| 12q13.13 | Yu et al. 2017 [17] | rs3741442 | 53346750 | 0.105465 | 0.31 | -4 | -4 | -4 | -4 |
| 12q13.2 | Yu et al. 2017 [17] | rs705704 | 56435412 | 0.0418832 | 1.22 | rs773107 | 56369506 | 0.0210978 | 1.26 |
| 12q21.1 | Yu et al. 2017 [17] | rs2304269 | 72080272 | 0.483316 | 0.86 | rs11178895 | 72089411 | 0.321804 | 0.84 |
| 13q31.1 | Ludwig et al. 2012 [12] | rs8001641 | 80692811 | 0.00268945 | 1.34 | rs11841646 | 80679302 | 0.00135074 | 1.37 |
| 14q22.1 | Ludwig et al. 2017 [19] | rs4901118 | 51856109 | 0.367607 | 1.10 | rs60454187 | 51856566 | 0.279705 | 0.9 |
| 14q22.1 | Yu et al. 2017 [17] | rs7148069 | 51839645 | 0.731853 | 1.03 | - | - | - | - |
| 14q32.13 | Yu et al. 2017 [17] | rs1243572 | 95379499 | 0.343413 | 1.12 | rs1243561 | 95369886 | 0.201146 | 1.16 |
| 15q13 | Ludwig et al. 2016 [15] | rs1258763 | 33050423 | 0.0521438 | 1.19 | rs13329310 | 33052553 | 0.0499003 | 0.83 |
| 15q22.2 | Ludwig et al. 2012 [12] | rs1873147 | 63312632 | 0.469881 | 1.08 | rs12902152 | 63313968 | 0.167601 | 1.19 |
| 15q24 | Ludwig et al. 2017 [19] | rs28689146 | 75005575 | 0.442303 | 1.08 | - | - | - | - |
| 16p13.3 | Sun et al. 2015 [14] | rs8049367 | 3980445 | 0.159099 | 1.13 | rs11076792 | 3968567 | 0.00316807 | 1.33 |
| 17q13.1 | Beaty et al. 2010 [11] | rs9891446 | 8935416 | 0.186388 | 1.14 | - | - | - | - |
| 17q21.32 | Yu et al. 2017 [17] | rs1838105 | 45008935 | 0.00231174 | 0.73 | rs197907 | 44982081 | 0.000282387 | 1.45 |
| 17q22 | Mangold et al. 2010 [10] | rs227727 | 54776955 | 0.180057 | 1.14 | - | - | - | - |
| 17q23.2 | Leslie et al. 2016 [16] | rs1588366 | 61076428 | 0.00901131 | 0.75 | rs72843145 | 61052949 | 0.0057287 | 0.72 |
| 19p13.3 | Ludwig et al. 2017 [19] | rs3746101 | 2050823 | 0.929281 | 0.95 | - | - | - | - |
| 19q12 | Leslie et al. 2016 [16] | rs73039426 | 33520961 | 0.26959 | 1.20 | - | - | - | - |
| 20q12 | Beaty et al. 2010 [11] | rs13041247 | 39269074 | 0.0101081 | 0.77 | rs34753522 | 39278391 | 0.00037345 | 0.69 |
| Chromosome | p-Value 1 | |
|---|---|---|
| SH3PXD2A | 10 | 2.3729 × 10−8 |
| ANO4 | 12 | 1.4588 × 10−7 |
| CMSS1 | 3 | 2.9078 × 10−6 |
| FILIP1L | 3 | 7.1274 × 10−6 |
| CLEC3A | 16 | 1.0871 × 10−5 |
| PAX72 | 1 | 1.9157 × 10−5 |
| CCDC140 | 2 | 4.4909 × 10−5 |
| ESR1 | 6 | 7.1554 × 10−5 |
| IFITM3 | 11 | 8.5273 × 10−5 |
| PANX1 | 11 | 8.7342 × 10−5 |
| BLMH | 17 | 9.5824 × 10−5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Rooij, I.A.; Ludwig, K.U.; Welzenbach, J.; Ishorst, N.; Thonissen, M.; Galesloot, T.E.; Ongkosuwito, E.; Bergé, S.J.; Aldhorae, K.; Rojas-Martinez, A.; et al. Non-Syndromic Cleft Lip with or without Cleft Palate: Genome-Wide Association Study in Europeans Identifies a Suggestive Risk Locus at 16p12.1 and Supports SH3PXD2A as a Clefting Susceptibility Gene. Genes 2019, 10, 1023. https://doi.org/10.3390/genes10121023
van Rooij IA, Ludwig KU, Welzenbach J, Ishorst N, Thonissen M, Galesloot TE, Ongkosuwito E, Bergé SJ, Aldhorae K, Rojas-Martinez A, et al. Non-Syndromic Cleft Lip with or without Cleft Palate: Genome-Wide Association Study in Europeans Identifies a Suggestive Risk Locus at 16p12.1 and Supports SH3PXD2A as a Clefting Susceptibility Gene. Genes. 2019; 10(12):1023. https://doi.org/10.3390/genes10121023
Chicago/Turabian Stylevan Rooij, Iris ALM, Kerstin U Ludwig, Julia Welzenbach, Nina Ishorst, Michelle Thonissen, Tessel E Galesloot, Edwin Ongkosuwito, Stefaan J Bergé, Khalid Aldhorae, Augusto Rojas-Martinez, and et al. 2019. "Non-Syndromic Cleft Lip with or without Cleft Palate: Genome-Wide Association Study in Europeans Identifies a Suggestive Risk Locus at 16p12.1 and Supports SH3PXD2A as a Clefting Susceptibility Gene" Genes 10, no. 12: 1023. https://doi.org/10.3390/genes10121023
APA Stylevan Rooij, I. A., Ludwig, K. U., Welzenbach, J., Ishorst, N., Thonissen, M., Galesloot, T. E., Ongkosuwito, E., Bergé, S. J., Aldhorae, K., Rojas-Martinez, A., Kiemeney, L. A., Vermeesch, J. R., Brunner, H., Roeleveld, N., Devriendt, K., Dormaar, T., Hens, G., Knapp, M., Carels, C., & Mangold, E. (2019). Non-Syndromic Cleft Lip with or without Cleft Palate: Genome-Wide Association Study in Europeans Identifies a Suggestive Risk Locus at 16p12.1 and Supports SH3PXD2A as a Clefting Susceptibility Gene. Genes, 10(12), 1023. https://doi.org/10.3390/genes10121023