It’s All in the Genes: The Regulatory Pathways of Sexual Reproduction in Filamentous Ascomycetes

 , , and
, , and
Abstract
:
1. Introduction
2. Protoascomatal Development
2.1. The Mating Type Genes
2.2. Important Signalling Pathways
2.3. Nutrient Requirements for the Induction of Sexual Reproduction
2.3.1. Sugars
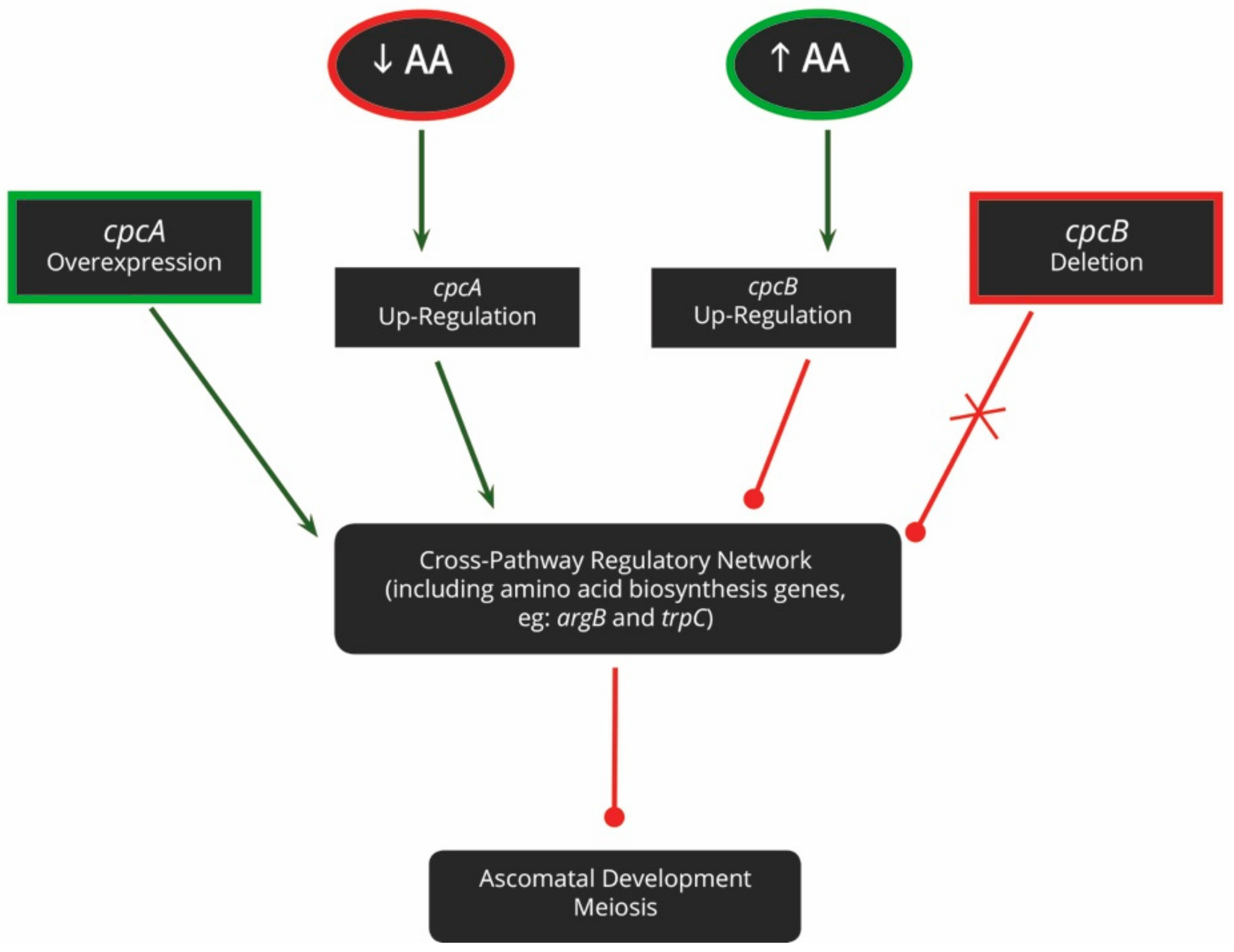
2.3.2. Amino Acids
2.3.3. Calcium
2.4. Other Environmental Triggers
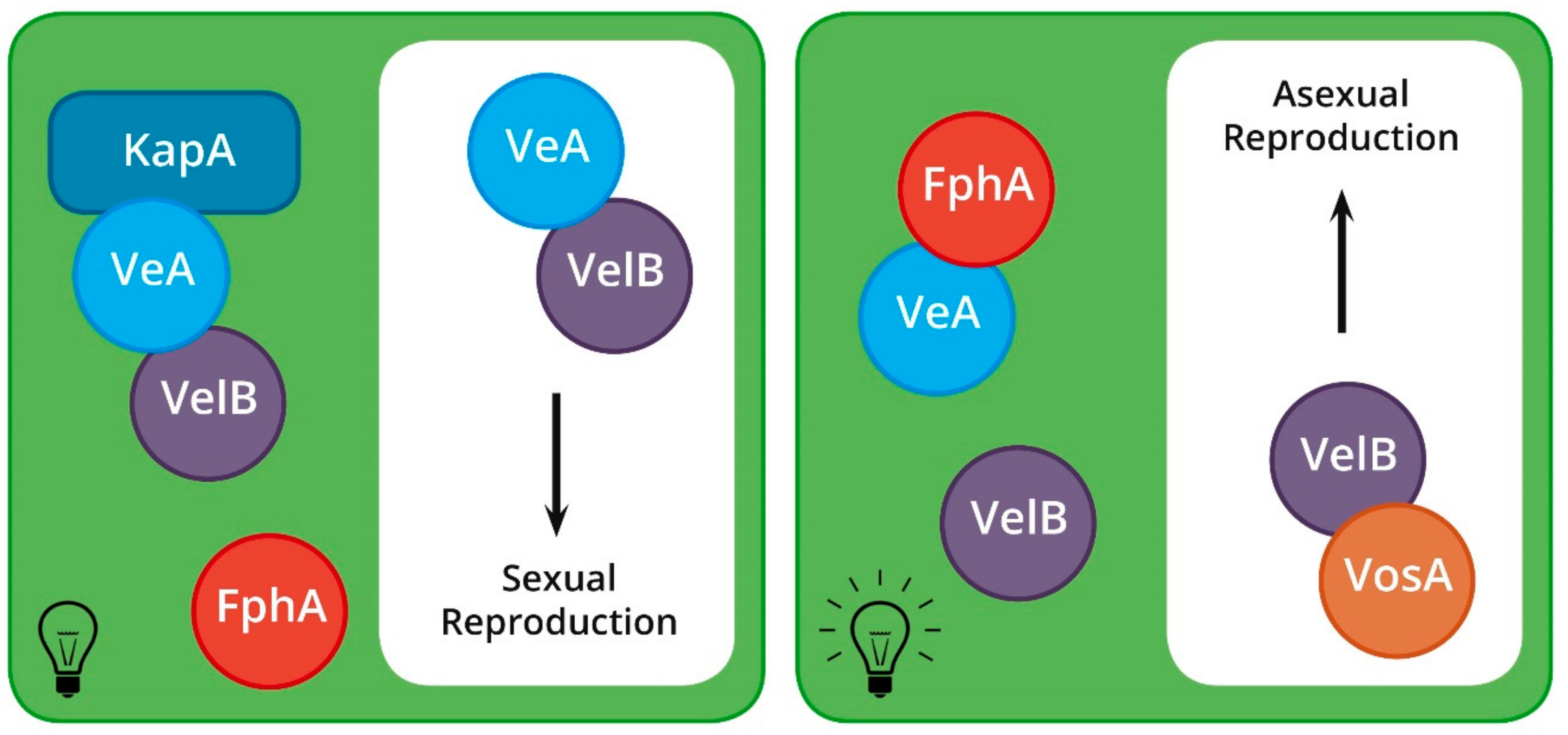
2.4.1. Light
2.4.2. Reactive Oxygen Species
2.4.3. Pheromones
3. Ascus Production
4. Ascospore Production
4.1. Meiosis and Ascospore Production
4.2. Ascospore Discharge
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hawksworth, D.L.; Luecking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 Million Species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D. Global Species Numbers of Fungi: Are Tropical Studies and Molecular Approaches Contributing to a More Robust Estimate? Biodivers. Conserv. 2012, 21, 2425–2433. [Google Scholar] [CrossRef]
- Billiard, S.; Lopez Villavicencio, M.; Hood, M.; Giraud, T. Sex, Outcrossing and Mating Types: Unsolved Questions in Fungi and Beyond. J. Evol. Biol. 2012, 25, 1020–1038. [Google Scholar] [CrossRef]
- Ni, M.; Feretzaki, M.; Sun, S.; Wang, X.; Heitman, J. Sex in Fungi. Annu. Rev. Genet. 2011, 45, 405–430. [Google Scholar] [CrossRef] [PubMed]
- James, T.Y.; Kauff, F.; Schoch, C.L.; Matheny, P.B.; Hofstetter, V.; Cox, C.J.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J. Reconstructing the Early Evolution of Fungi using a Six-Gene Phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Stajich, J.E.; Berbee, M.L.; Blackwell, M.; Hibbett, D.S.; James, T.Y.; Spatafora, J.W.; Taylor, J.W. The Fungi. Curr. Biol. 2009, 19, R840–R845. [Google Scholar] [CrossRef] [PubMed]
- Dyer, P.S. Sexual Reproduction and the Significance of MAT in the Aspergilli. In Sex in Fungi: Molecular Determination and Evolutionary Implications; Heitman, J., Kronstad, J.W., Taylor, J.W., Casselton, L.A., Eds.; American Society of Microbiology Press: Washington, DC, USA, 2007; pp. 123–142. [Google Scholar]
- Dyer, P.; Inderbitzin, P.; Debuchy, R. Mating-Type Structure, Function, Regulation and Evolution in the Pezizomycotina. In Growth, Differentiation and Sexuality. The Mycota; Wendland, J., Ed.; Springer: Cham, Switzerland, 2016; Volume 1, pp. 351–385. [Google Scholar]
- Klix, V.; Nowrousian, M.; Ringelberg, C.; Loros, J.; Dunlap, J.; Pöggeler, S. Functional Characterization of MAT1-1-Specific Mating-Type Genes in the Homothallic Ascomycete Sordaria macrospora provides New Insights into Essential and Nonessential Sexual Regulators. Eukaryot. Cell 2010, 9, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.V.-B.; An, Z.; Metzenberg, R.L.; Glass, N.L. Characterization of matA-2, matA-3 and ΔmatA Mating-Type Mutants of Neurospora crassa. Genetics 1998, 148, 1069–1079. [Google Scholar] [PubMed]
- Butler, G. The Evolution of MAT: The Ascomycetes. In Sex in Fungi: Molecular Determination and Evolutionary Implications; Heitman, J.K., Taylor, J.W., Casselton, L.A., Eds.; American Society of Microbiology Press: Washington, DC, USA, 2007; pp. 3–18. [Google Scholar]
- Blakeslee, A.F. Sexual Reproduction in the Mucorineae. Proc. Am. Acad. Arts Sci. 1904, 40, 205–319. [Google Scholar] [CrossRef]
- Lin, X.; Heitman, J. Mechanisms of Homothallism in Fungi and Transitions between Heterothallism and Homothallism. In Sex in Fungi: Molecular Determination and Evolutionary Implications; Heitman, J., Kronstad, J.W., Taylor, J.W., Casselton, L.A., Eds.; American Society of Microbiology Press: Washington, DC, USA, 2007; pp. 35–57. [Google Scholar]
- Turgeon, B.G.; Yoder, O. Proposed Nomenclature for Mating Type Genes of Filamentous Ascomycetes. Fungal Genet. Biol. 2000, 31, 1–5. [Google Scholar] [CrossRef]
- Wilken, P.M.; Steenkamp, E.T.; Wingfield, M.J.; De Beer, Z.W.; Wingfield, B.D. Which MAT gene? Pezizomycotina (Ascomycota) Mating-Type Gene Nomenclature Reconsidered. Fungal Biol. Rev. 2017, 31, 199–211. [Google Scholar] [CrossRef]
- Colot, H.V.; Park, G.; Turner, G.E.; Ringelberg, C.; Crew, C.M.; Litvinkova, L.; Weiss, R.L.; Borkovich, K.A.; Dunlap, J.C. A High-Throughput Gene Knockout Procedure for Neurospora reveals Functions for Multiple Transcription Factors. Proc. Natl. Acad. Sci. USA 2006, 103, 10352–10357. [Google Scholar] [CrossRef]
- Vienken, K.; Fischer, R. The Zn(II)2Cys6 Putative Transcription Factor nosA controls Fruiting Body Formation in Aspergillus nidulans. Mol. Microbiol. 2006, 61, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Vienken, K.; Scherer, M.; Fischer, R. The Zn(II)2Cys6 Putative Aspergillus nidulans Transcription Factor RosA (Repressor of Sexual Development) Inhibits Sexual Development Under Low-Carbon Conditions and in Submersed Culture. Genetics 2005, 169, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Choi, G.H.; Nuss, D.L. Hypovirus-Responsive Transcription Factor Gene pro1 of the Chestnut Blight Fungus Cryphonectria parasitica is required for Female Fertility, Asexual Spore Development, and Stable Maintenance of Hypovirus Infection. Eukaryot. Cell 2009, 8, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Gautier, V.; Tong, L.; Nguyen, T.-S.; Debuchy, R.; Silar, P. PaPro1 and IDC4, Two Genes controlling Stationary Phase, Sexual Development and Cell Degeneration in Podospora anserina. J. Fungi 2018, 4, 85. [Google Scholar] [CrossRef] [PubMed]
- Benkhali, J.A.; Coppin, E.; Brun, S.; Peraza-Reyes, L.; Martin, T.; Dixelius, C.; Lazar, N.; Van Tilbeurgh, H.; Debuchy, R. A Network of HMG-Box Transcription Factors regulates Sexual Cycle in the Fungus Podospora anserina. Plos Genet. 2013, 9, e1003642. [Google Scholar] [CrossRef]
- Koopman, P. HMG Domain Superfamily of DNA-bending Proteins: HMG, UBF, TCF, LEF, SOX, SRY and Related Proteins. e LS 2001. [Google Scholar] [CrossRef]
- Bazafkan, H.; Dattenböck, C.; Böhmdorfer, S.; Tisch, D.; Stappler, E.; Schmoll, M. Mating Type-Dependent Partner Sensing as Mediated by VEL1 in Trichoderma reesei. Mol. Microbiol. 2015, 96, 1103–1118. [Google Scholar] [CrossRef]
- Seibel, C.; Tisch, D.; Kubicek, C.P.; Schmoll, M. ENVOY is a Major Determinant in Regulation of Sexual Development in Hypocrea jecorina (Trichoderma reesei). Eukaryot. Cell 2012, 11, 885–895. [Google Scholar] [CrossRef]
- Castellanos, F.; Schmoll, M.; Martínez, P.; Tisch, D.; Kubicek, C.P.; Herrera-Estrella, A.; Esquivel-Naranjo, E.U. Crucial Factors of the Light Perception Machinery and their Impact on Growth and Cellulase Gene Transcription in Trichoderma reesei. Fungal Genet. Biol. 2010, 47, 468–476. [Google Scholar] [CrossRef]
- Schmoll, M.; Esquivel-Naranjo, E.U.; Herrera-Estrella, A. Trichoderma in the Light of Day—Physiology and Development. Fungal Genet. Biol. 2010, 47, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, M.; Franchi, L.; Kubicek, C.P. ENVOY, a PAS/LOV Domain Protein of Hypocrea jecorina (Anamorph Trichoderma reesei), modulates Cellulase Gene Transcription in Response to Light. Eukaryot. Cell 2005, 4, 1998–2007. [Google Scholar] [CrossRef]
- Debuchy, R.; Coppin, E. The Mating Types of Podospora anserina: Functional Analysis and Sequence of the Fertilization Domains. Mol. Gen. Genet. 1992, 233, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, M.; Seymour, F.A.; Alcocer, M.J.; Kaur, N.; Calvo, A.M.; Archer, D.B.; Dyer, P.S. Mating Type and the Genetic Basis of Self-Fertility in the Model Fungus Aspergillus nidulans. Curr. Biol. 2007, 17, 1384–1389. [Google Scholar] [CrossRef] [PubMed]
- Staben, C.; Yanofsky, C. Neurospora crassa a Mating-Type Region. Proc. Natl. Acad. Sci. USA 1990, 87, 4917–4921. [Google Scholar] [CrossRef]
- Lee, J.; Lee, T.; Lee, Y.W.; Yun, S.H.; Turgeon, B.G. Shifting Fungal Reproductive Mode by Manipulation of Mating Type Genes: Obligatory Heterothallism of Gibberella zeae. Mol. Microbiol. 2003, 50, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Böhm, J.; Hoff, B.; O’Gorman, C.M.; Wolfers, S.; Klix, V.; Binger, D.; Zadra, I.; Kürnsteiner, H.; Pöggeler, S.; Dyer, P.S. Sexual Reproduction and Mating-Type–Mediated Strain Development in the Penicillin-Producing Fungus Penicillium chrysogenum. Proc. Natl. Acad. Sci. USA 2013, 110, 1476–1481. [Google Scholar] [CrossRef] [PubMed]
- Böhm, J.; Dahlmann, T.A.; Gümüşer, H.; Kück, U. A MAT1–2 Wild-Type Strain from Penicillium chrysogenum: Functional Mating-Type Locus Characterization, Genome Sequencing and Mating with an Industrial Penicillin-Producing Strain. Mol. Microbiol. 2015, 95, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Pöggeler, S.; Nowrousian, M.; Ringelberg, C.; Loros, J.; Dunlap, J.; Kück, U. Microarray and Real-Time PCR Analyses Reveal Mating Type-Dependent Gene Expression in a Homothallic Fungus. Mol. Genet. Genom. 2006, 275, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-K.; Cho, E.J.; Lee, S.; Lee, Y.-S.; Yun, S.-H. Functional Analyses of Individual Mating-Type Transcripts at MAT Loci in Fusarium graminearum and Fusarium asiaticum. FEMS Microbiol. Lett. 2012, 337, 89–96. [Google Scholar] [CrossRef]
- Arnaise, S.; Debuchy, R.; Picard, M. What is a Bona Fide Mating-Type Gene? Internuclear Complementation of mat Mutants in Podospora anserina. Mol. Gen. Genet. 1997, 256, 169–178. [Google Scholar] [CrossRef]
- Doughan, B.; Rollins, J.A. Characterization of MAT Gene Functions in the Life Cycle of Sclerotinia sclerotiorum Reveals a Lineage-Specific MAT Gene Functioning in Apothecium Morphogenesis. Fungal Biol. 2016, 120, 1105–1117. [Google Scholar] [CrossRef]
- Rodenburg, S.Y.; Terhem, R.B.; Veloso, J.; Stassen, J.H.; van Kan, J.A. Functional Analysis of Mating Type Genes and Transcriptome Analysis during Fruiting Body Development of Botrytis cinerea. mBio 2018, 9, e01939-17. [Google Scholar] [CrossRef]
- Arnaise, S.; Zickler, D.; Le Bilcot, S.; Poisier, C.; Debuchy, R. Mutations in Mating-Type Genes of the Heterothallic Fungus Podospora anserina lead to Self-Fertility. Genetics 2001, 159, 545–556. [Google Scholar] [PubMed]
- Zickler, D.; Arnaise, S.; Coppin, E.; Debuchy, R.; Picard, M. Altered Mating-Type Identity in the Fungus Podospora anserina leads to Selfish Nuclei, Uniparental Progeny, and Haploid Meiosis. Genetics 1995, 140, 493–503. [Google Scholar] [PubMed]
- Yu, Y.; Amich, J.; Will, C.; Eagle, C.E.; Dyer, P.S.; Krappmann, S. The Novel Aspergillus fumigatus MAT1-2-4 Mating-Type Gene is Required for Mating and Cleistothecia Formation. Fungal Genet. Biol. 2017, 108, 1–12. [Google Scholar] [CrossRef]
- Braus, G.H.; Irniger, S.; Bayram, Ö. Fungal Development and the COP9 Signalosome. Curr. Opin. Microbiol. 2010, 13, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Busch, S.; Schwier, E.U.; Nahlik, K.; Bayram, Ö.; Helmstaedt, K.; Draht, O.W.; Krappmann, S.; Valerius, O.; Lipscomb, W.N.; Braus, G.H. An Eight-Subunit COP9 Signalosome with an Intact JAMM Motif is required for Fungal Fruit Body Formation. Proc. Natl. Acad. Sci. USA 2007, 104, 8089–8094. [Google Scholar] [CrossRef]
- Busch, S.; Eckert, S.E.; Krappmann, S.; Braus, G.H. The COP9 Signalosome is an Essential Regulator of Development in the Filamentous Fungus Aspergillus nidulans. Mol. Microbiol. 2003, 49, 717–730. [Google Scholar] [CrossRef]
- Teichert, I.; Nowrousian, M.; Pöggeler, S.; Kück, U. The Filamentous Fungus Sordaria macrospora as a Genetic Model to Study Fruiting Body Development. Adv. Genet. 2014, 87, 199–244. [Google Scholar]
- Hwang, J.; Pallas, D.C. STRIPAK Complexes: Structure, Biological Function, and Involvement in Human Diseases. Int. J. Biochem. Cell Biol. 2014, 47, 118–148. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, P.S.; Josué, F.; Wepf, A.; Wehr, M.C.; Rinner, O.; Kelly, G.; Tapon, N.; Gstaiger, M. Combined Functional Genomic and Proteomic Approaches identify a PP2A Complex as a Negative Regulator of Hippo Signaling. Mol. Cell 2010, 39, 521–534. [Google Scholar] [CrossRef]
- Bernhards, Y.; Pöggeler, S. The Phocein Homologue SmMOB3 is Essential for Vegetative Cell Fusion and Sexual Development in the Filamentous Ascomycete Sordaria macrospora. Curr. Genet. 2011, 57, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Bloemendal, S.; Bernhards, Y.; Bartho, K.; Dettmann, A.; Voigt, O.; Teichert, I.; Seiler, S.; Wolters, D.A.; Pöggeler, S.; Kück, U. A Homologue of the Human STRIPAK Complex controls Sexual Development in Fungi. Mol. Microbiol. 2012, 84, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Pöggeler, S.; Kück, U. A WD40 Repeat Protein regulates Fungal Cell Differentiation and can be replaced Functionally by the Mammalian Homologue Striatin. Eukaryot. Cell 2004, 3, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Lee, D.B.; Kim, J.H.; Kim, M.S.; Han, K.Y.; Kim, W.S.; Park, Y.S.; Kim, H.B.; Han, D.M. Environmental Factors Affecting Development of Aspergillus nidulans. J. Microbiol. 2003, 41, 34–40. [Google Scholar]
- Malagnac, F.; Lalucque, H.; Lepère, G.; Silar, P. Two NADPH Oxidase Isoforms are required for Sexual Reproduction and Ascospore Germination in the Filamentous Fungus Podospora anserina. Fungal Genet. Biol. 2004, 41, 982–997. [Google Scholar] [CrossRef]
- Johnston, M. Feasting, Fasting and Fermenting: Glucose Sensing in Yeast and Other Cells. Trends Genet. 1999, 15, 29–33. [Google Scholar] [CrossRef]
- Kraakman, L.; Lemaire, K.; Ma, P.; Teunissen, A.W.; Donaton, M.C.; Van Dijck, P.; Winderickx, J.; De Winde, J.H.; Thevelein, J.M. A Saccharomyces cerevisiae G-Protein Coupled Receptor, Gpr1, is specifically Required for Glucose Activation of the cAMP Pathway during the Transition to Growth on Glucose. Mol. Microbiol. 1999, 32, 1002–1012. [Google Scholar] [CrossRef]
- Welton, R.M.; Hoffman, C.S. Glucose Monitoring in Fission Yeast via the gpa2 Gα, the git5 Gβ and the git3 Putative Glucose Receptor. Genetics 2000, 156, 513–521. [Google Scholar]
- Brown, N.A.; dos Reis, T.F.; Ries, L.N.A.; Caldana, C.; Mah, J.H.; Yu, J.H.; Macdonald, J.M.; Goldman, G.H. G-Protein Coupled Receptor-Mediated Nutrient Sensing and Developmental Control in Aspergillus nidulans. Mol. Microbiol. 2015, 98, 420–439. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Seo, J.A.; Yu, J.H. A Putative G-Protein Coupled Receptor Negatively Controls Sexual Development in Aspergillus nidulans. Mol. Microbiol. 2004, 51, 1333–1345. [Google Scholar] [CrossRef]
- Hoffmann, B.; Wanke, C.; LaPaglia, S.K.; Braus, G.H. c-Jun and RACK1 Homologues Regulate a Control Point for Sexual Development in Aspergillus nidulans. Mol. Microbiol. 2000, 37, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Wang, L.; Liu, Z.; Kwon, N.J.; Kim, S.C.; Yu, J.H. Gβ-Like CpcB Plays a Crucial Role for Growth and Development of Aspergillus nidulans and Aspergillus fumigatus. PLoS ONE 2013, 8, e70355. [Google Scholar] [CrossRef] [PubMed]
- Eckert, S.E.; Hoffmann, B.; Wanke, C.; Braus, G.H. Sexual Development of Aspergillus nidulans in Tryptophan Auxotrophic Strains. Arch. Microbiol. 1999, 172, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Käfer, E. The Anthranilate Synthetase Enzyme Complex and the Trifunctional trpC gene of Aspergillus. Can. J. Genet. Cytol. 1977, 19, 723–738. [Google Scholar] [CrossRef]
- Serlupi-Crescenzi, O.; Kurtz, M.B.; Champe, S.P. Developmental Defects Resulting from Arginine Auxotrophy in Aspergillus nidulans. Microbiology 1983, 129, 3535–3544. [Google Scholar] [CrossRef]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The Versatility and Universality of Calcium Signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11. [Google Scholar] [CrossRef]
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the Crossroads of Signaling. Plant Cell 2002, 14, S401–S417. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Schnell, N.; Chattaway, J.; Davies, P.; Dixon, G.; Sanders, D. The Saccharomyces cerevisiae cch1 Gene is Involved in Calcium Influx and Mating. FEBS Lett. 1997, 419, 259–262. [Google Scholar] [CrossRef]
- Iida, H.; Nakamura, H.; Ono, T.; Okumura, M.S.; Anraku, Y. mid1, A Novel Saccharomyces cerevisiae Gene Encoding a Plasma Membrane Protein, is Required for Ca2+ Influx and Mating. Mol. Cell. Biol. 1994, 14, 8259–8271. [Google Scholar] [CrossRef]
- Muller, E.M.; Mackin, N.A.; Erdman, S.E.; Cunningham, K.W. FIG1p Facilitates Ca2+ Influx and Cell Fusion during Mating of Saccharomyces cerevisiae. J. Biol. Chem. 2003, 40, 38461–38469. [Google Scholar] [CrossRef]
- Cavinder, B.; Trail, F. Role of fig1, a Component of the Low-Affinity Calcium Uptake System, in Growth and Sexual Development of Filamentous Fungi. Eukaryot. Cell 2012, 11, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, J.E.; Son, H.; Frailey, D.; Cirino, R.; Lee, Y.W.; Duncan, R.; Czymmek, K.J.; Kang, S. Roles of Three Fusarium graminearum Membrane Ca2+ Channels in the Formation of Ca2+ Signatures, Growth, Development, Pathogenicity and Mycotoxin Production. Fungal Genet. Biol. 2018, 111, 30–46. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, H.; Long, N.; Carbó, N.; Chen, P.; Aguilar, P.S.; Lu, L. FigA, A Putative Homolog of Low-Affinity Calcium System Member Fig1 in Saccharomyces cerevisiae, is Involved in Growth and Asexual and Sexual Development in Aspergillus nidulans. Eukaryot. Cell 2014, 13, 295–303. [Google Scholar] [CrossRef]
- Rodriguez-Romero, J.; Hedtke, M.; Kastner, C.; Müller, S.; Fischer, R. Fungi, Hidden in Soil or Up in the Air: Light makes a Difference. Annu. Rev. Microbiol. 2010, 64, 585–610. [Google Scholar] [CrossRef]
- He, Q.; Cheng, P.; Yang, Y.; Wang, L.; Gardner, K.H.; Liu, Y. White Collar-1, A DNA Binding Transcription Factor and a Light Sensor. Science 2002, 297, 840–843. [Google Scholar] [CrossRef]
- Ballario, P.; Talora, C.; Galli, D.; Linden, H.; Macino, G. Roles in Dimerization and Blue Light Photoresponse of the PAS and LOV Domains of Neurospora crassa White Collar Proteins. Mol. Microbiol. 1998, 29, 719–729. [Google Scholar] [CrossRef]
- Linden, H.; Macino, G. White Collar 2, A Partner in Blue-Light Signal Transduction, Controlling Expression of Light–Regulated Genes in Neurospora crassa. Embo J. 1997, 16, 98–109. [Google Scholar]
- Ballario, P.; Vittorioso, P.; Magrelli, A.; Talora, C.; Cabibbo, A.; Macino, G. White Collar-1, A Central Regulator of Blue Light Responses in Neurospora, is a Zinc Finger Protein. EMBO J. 1996, 15, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, F.D.; Pohl, U.; Russo, V. Photoinduction of Protoperithecia in Neurospora crassa by Blue Light. Photochem. Photobiol. 1983, 37, 49–51. [Google Scholar] [CrossRef]
- Harding, R.W.; Melles, S. Genetic Analysis of Phototropism of Neurospora crassa Perithecial Beaks using White Collar and Albino Mutants. Plant Physiol. 1983, 72, 996–1000. [Google Scholar] [CrossRef]
- Mooney, J.L.; Yager, L.N. Light is Required for Conidiation in Aspergillus nidulans. Genes Dev. 1990, 4, 1473–1482. [Google Scholar] [CrossRef]
- Bayram, Ö.; Braus, G.H. Coordination of Secondary Metabolism and Development in Fungi: The Velvet Family of Regulatory Proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H. VelB/VeA/LaeA Complex Coordinates Light Signal with Fungal Development and Secondary Metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Han, K.Y.; Kim, K.J.; Han, D.M.; Jahng, K.Y.; Chae, K.S. The veA Gene Activates Sexual Development in Aspergillus nidulans. Fungal Genet. Biol. 2002, 37, 72–80. [Google Scholar] [CrossRef]
- Stinnett, S.M.; Espeso, E.A.; Cobeño, L.; Araújo-Bazán, L.; Calvo, A.M. Aspergillus nidulans VeA Subcellular Localization is Dependent on the Importin α Carrier and on Light. Mol. Microbiol. 2007, 63, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Yu, J.H. A Novel Regulator Couples Sporogenesis and Trehalose Biogenesis in Aspergillus nidulans. PLoS ONE 2007, 2, e970. [Google Scholar] [CrossRef]
- Blumenstein, A.; Vienken, K.; Tasler, R.; Purschwitz, J.; Veith, D.; Frankenberg-Dinkel, N.; Fischer, R. The Aspergillus nidulans Phytochrome FphA Represses Sexual Development in Red Light. Curr. Biol. 2005, 15, 1833–1838. [Google Scholar] [CrossRef]
- Purschwitz, J.; Müller, S.; Fischer, R. Mapping the Interaction Sites of Aspergillus nidulans Phytochrome FphA with the Global Regulator VeA and the White Collar Protein LreB. Mol. Genet. Genom. 2009, 281, 35–42. [Google Scholar] [CrossRef]
- Aguirre, J.; Ríos-Momberg, M.; Hewitt, D.; Hansberg, W. Reactive Oxygen Species and Development in Microbial Eukaryotes. Trends Microbiol. 2005, 13, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, D.; Tanaka, A.; Scott, B. NADPH Oxidases in Fungi: Diverse Roles of Reactive Oxygen Species in Fungal Cellular Differentiation. Fungal Genet. Biol. 2007, 44, 1065–1076. [Google Scholar] [PubMed]
- Scherer, M.; Wei, H.; Liese, R.; Fischer, R. Aspergillus nidulans Catalase-Peroxidase Gene (cpeA) is Transcriptionally Induced during Sexual Development through the Transcription Factor StuA. Eukaryot. Cell 2002, 1, 725–735. [Google Scholar]
- Lara-Ortíz, T.; Riveros-Rosas, H.; Aguirre, J. Reactive Oxygen Species generated by Microbial NADPH Oxidase NoxA regulate Sexual Development in Aspergillus nidulans. Mol. Microbiol. 2003, 50, 1241–1255. [Google Scholar]
- Cano-Domínguez, N.; Álvarez-Delfín, K.; Hansberg, W.; Aguirre, J. NADPH Oxidases NOX-1 and NOX-2 require the Regulatory Subunit NOR-1 to control Cell Differentiation and Growth in Neurospora crassa. Eukaryot. Cell 2008, 7, 1352–1361. [Google Scholar] [CrossRef]
- Stowers, L.; Liberles, S.D. State-Dependent Responses to Sex Pheromones in Mouse. Curr. Opin. Neurobiol. 2016, 38, 74–79. [Google Scholar] [PubMed]
- Raabe, M. Insect Reproduction: Regulation of Successive Steps. Adv. Insect Physiol. 1987, 19, 29–154. [Google Scholar]
- Houck, L.D. Pheromone Communication in Amphibians and Reptiles. Annu. Rev. Physiol. 2009, 71, 161–176. [Google Scholar] [PubMed]
- Karlson, P.; Luscher, M. ‘Pheromones’: A New Term for a Class of Biologically Active Substances. Nature 1959, 183, 55–56. [Google Scholar] [CrossRef]
- Bölker, M.; Kahmann, R. Sexual Pheromones and Mating Responses in Fungi. Plant Cell 1993, 5, 1461. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.K.; Bennett, R.J. Fungal Mating Pheromones: Choreographing the Dating Game. Fungal Genet. Biol. 2011, 48, 668–676. [Google Scholar] [CrossRef]
- Zhang, L.; Baasiri, R.A.; Van Alfen, N.K. Viral Repression of Fungal Pheromone Precursor Gene Expression. Mol. Cell. Biol. 1998, 18, 953–959. [Google Scholar] [CrossRef]
- Shen, W.-C.; Bobrowicz, P.; Ebbole, D.J. Isolation of Pheromone Precursor Genes of Magnaporthe grisea. Fungal Genet. Biol. 1999, 27, 253–263. [Google Scholar] [CrossRef]
- Bobrowicz, P.; Pawlak, R.; Correa, A.; Bell-Pedersen, D.; Ebbole, D.J. The Neurospora crassa Pheromone Precursor Genes are Regulated by the Mating Type Locus and the Circadian Clock. Mol. Microbiol. 2002, 45, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Coppin, E.; de Renty, C.; Debuchy, R. The Function of the Coding Sequences for the Putative Pheromone Precursors in Podospora anserina is Restricted to Fertilization. Eukaryot. Cell 2004, 4, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Causes and Consequences of Variability in Peptide Mating Pheromones of Ascomycete Fungi. Mol. Biol. Evol. 2011, 28, 1987–2003. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.M.; van der Nest, M.A.; Wilken, P.M.; Wingfield, M.J.; Wingfield, B.D. Pheromone Expression reveals Putative Mechanism of Unisexuality in a Saprobic Ascomycete Fungus. PLoS ONE 2018, 13, e0192517. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Leslie, J.F.; Bowden, R.L. Expression and Function of Sex Pheromones and Receptors in the Homothallic Ascomycete Gibberella zeae. Eukaryot. Cell 2008, 7, 1211–1221. [Google Scholar] [CrossRef]
- Pöggeler, S. Two Pheromone Precursor Genes are Transcriptionally Expressed in the Homothallic Ascomycete Sordaria macrospora. Curr. Genet. 2000, 37, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Borkovich, K.A. Pheromones are Essential for Male Fertility and Sufficient to Direct Chemotropic Polarized Growth of Trichogynes during Mating in Neurospora crassa. Eukaryot. Cell 2006, 5, 544–554. [Google Scholar] [CrossRef]
- Kim, H.; Metzenberg, R.L.; Nelson, M.A. Multiple Functions of mfa-1, a Putative Pheromone Precursor Gene of Neurospora crassa. Eukaryot. Cell 2002, 1, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Turina, M.; Prodi, A.; Van Alfen, N.K. Role of the mf1-1 Pheromone Precursor Gene of the Filamentous Ascomycete Cryphonectria parasitica. Fungal Genet. Biol. 2003, 40, 242–251. [Google Scholar] [CrossRef]
- Zhang, L.; Churchill, A.; Kazmierczak, P.; Kim, D.H.; Van Alfen, N. Hypovirulence-Associated Traits Induced by a Mycovirus of Cryphonectria parasitica are Mimicked by Targeted Inactivation of a Host Gene. Mol. Cell. Biol. 1993, 13, 7782–7792. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, M.; Seibel, C.; Tisch, D.; Dorrer, M.; Kubicek, C.P. A Novel Class of Peptide Pheromone Precursors in Ascomycetous Fungi. Mol. Microbiol. 2010, 77, 1483–1501. [Google Scholar] [CrossRef]
- Xue, C.; Hsueh, Y.P.; Heitman, J. Magnificent Seven: Roles of G Protein-Coupled Receptors in Extracellular Sensing in Fungi. FEMS Microbiol. Rev. 2008, 32, 1010–1032. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Borkovich, K.A. A Pheromone Receptor Gene, pre-1, is Essential for Mating Type-Specific Directional Growth and Fusion of Trichogynes and Female Fertility in Neurospora crassa. Mol. Microbiol. 2004, 52, 1781–1798. [Google Scholar] [CrossRef]
- Kim, H.; Wright, S.J.; Park, G.; Ouyang, S.; Krystofova, S.; Borkovich, K.A. Roles for Receptors, Pheromones, G Proteins and Mating Type Genes During Sexual Reproduction in Neurospora crassa. Genetics 2012, 190, 1389–1404. [Google Scholar] [CrossRef]
- Pöggeler, S.; Kück, U. Comparative Analysis of the Mating-Type Loci from Neurospora crassa and Sordaria macrospora: Identification of Novel Transcribed ORFs. Mol. Gen. Genet. 2000, 263, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Pöggeler, S.; Kück, U. Identification of Transcriptionally Expressed Pheromone Receptor Genes in Filamentous Ascomycetes. Gene 2001, 280, 9–17. [Google Scholar] [CrossRef]
- Seo, J.A.; Han, K.H.; Yu, J.H. The gprA and gprB Genes Encode Putative G Protein-Coupled Receptors Required for Self-Fertilization in Aspergillus nidulans. Mol. Microbiol. 2004, 53, 1611–1623. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.A.; Han, K.H.; Yu, J.H. Multiple Roles of a Heterotrimeric G-Protein γ-Subunit in Governing Growth and Development of Aspergillus nidulans. Genetics 2005, 171, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Dyer, P.S.; Paoletti, M.; Archer, D.B. Genomics Reveals Sexual Secrets of Aspergillus. Microbiology 2003, 149, 2301–2303. [Google Scholar] [CrossRef]
- Coppin, E.; Debuchy, R.; Arnaise, S.; Picard, M. Mating Types and Sexual Development in Filamentous Ascomycetes. Microbiol. Mol. Biol. Rev. 1997, 61, 411–428. [Google Scholar] [PubMed]
- Son, H.; Lee, J.; Lee, Y.-W. A Novel Gene, gea1, is Required for Ascus Cell-Wall Development in the Ascomycete Fungus Fusarium graminearum. Microbiology 2013, 159, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, C.; Kim, J.C.; Kim, J.E.; Lee, Y.W. Identification and Functional Characterization of Genes Involved in the Sexual Reproduction of the Ascomycete Fungus Gibberella zeae. Biochem. Biophys. Res. Commun. 2010, 401, 48–52. [Google Scholar] [CrossRef]
- Cao, S.; He, Y.; Hao, C.; Xu, Y.; Zhang, H.; Wang, C.; Liu, H.; Xu, J.-R. RNA Editing of the amd1 Gene is Important for Ascus Maturation and Ascospore Discharge in Fusarium graminearum. Sci. Rep. 2017, 7, 4617. [Google Scholar] [CrossRef]
- Feng, B.; Haas, H.; Marzluf, G.A. ASD4, A New GATA factor of Neurospora crassa, Displays Sequence-Specific DNA Binding and Functions in Ascus and Ascospore Development. Biochemistry 2000, 39, 11065–11073. [Google Scholar] [CrossRef]
- Wang, Z.; Lopez-Giraldez, F.; Lehr, N.; Farré, M.; Common, R.; Trail, F.; Townsend, J.P. Global Gene Expression and Focused Knockout Analysis reveals Genes Associated with Fungal Fruiting Body Development in Neurospora crassa. Eukaryot. Cell 2014, 13, 154–169. [Google Scholar] [CrossRef]
- Lee, D.W.; Pratt, R.J.; McLaughlin, M.; Aramayo, R. An Argonaute-Like Protein is Required for Meiotic Silencing. Genetics 2003, 164, 821–828. [Google Scholar] [PubMed]
- Bayne, E.H.; White, S.A.; Kagansky, A.; Bijos, D.A.; Sanchez-Pulido, L.; Hoe, K.L.; Kim, D.U.; Park, H.O.; Ponting, C.P.; Rappsilber, J. STC1: A Critical Link between RNAi and Chromatin Modification required for Heterochromatin Integrity. Cell 2010, 140, 666–677. [Google Scholar] [CrossRef]
- Berteaux-Lecellier, V.; Picard, M.; Thompson-Coffe, C.; Zickler, D.; Panvier-Adoutte, A.; Simonet, J.M. A Nonmammalian Homolog of the PAF1 Gene (Zellweger syndrome) Discovered as a Gene Involved in Caryogamy in the Fungus Podospora anserina. Cell 1995, 81, 1043–1051. [Google Scholar] [CrossRef]
- Boisnard, S.; Zickler, D.; Picard, M.; Berteaux-Lecellier, V. Overexpression of a Human and a Fungal ABC Transporter similarly Suppresses the Differentiation Defects of a Fungal Peroxisomal Mutant but Introduces Pleiotropic Cellular Effects. Mol. Microbiol. 2003, 49, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Peraza Reyes, L.; Berteaux-Lecellier, V. Peroxisomes and Sexual Development in Fungi. Front. Physiol. 2013, 4, 244. [Google Scholar] [CrossRef] [PubMed]
- Kirk, K.; Morris, N. The tubB α-Tubulin Gene is Essential for Sexual Development in Aspergillus nidulans. Genes Dev. 1991, 5, 2014–2023. [Google Scholar] [CrossRef]
- Krappmann, S.; Jung, N.; Medic, B.; Busch, S.; Prade, R.A.; Braus, G.H. The Aspergillus nidulans F-Box Protein GrrA links SCF Activity to Meiosis. Mol. Microbiol. 2006, 61, 76–88. [Google Scholar] [CrossRef]
- Shiu, P.K.; Raju, N.B.; Zickler, D.; Metzenberg, R.L. Meiotic Silencing by Unpaired DNA. Cell 2001, 107, 905–916. [Google Scholar] [CrossRef]
- Shiu, P.K.; Zickler, D.; Raju, N.B.; Ruprich-Robert, G.; Metzenberg, R.L. SAD-2 is Required for Meiotic Silencing by Unpaired DNA and Perinuclear Localization of SAD-1 RNA-Directed RNA Polymerase. Proc. Natl. Acad. Sci. USA 2006, 103, 2243–2248. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.A.; Merino, S.T.; Metzenberg, R.L. A Putative Rhamnogalacturonase Required for Sexual Development of Neurospora crassa. Genetics 1997, 146, 531–540. [Google Scholar]
- Hu, S.; Zhou, X.; Gu, X.; Cao, S.; Wang, C.; Xu, J.R. The cAMP-PKA Pathway Regulates Growth, Sexual and Asexual Differentiation, and Pathogenesis in Fusarium graminearum. Mol. Plant Microbe Interact. 2014, 27, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Cavinder, B.; Hamam, A.; Lew, R.R.; Trail, F. Mid1, a Mechanosensitive Calcium Ion Channel, affects Growth, Development, and Ascospore discharge in the Filamentous Fungus Gibberella zeae. Eukaryot. Cell 2011, 10, 832–841. [Google Scholar] [CrossRef]
- Hallen, H.E.; Trail, F. The L-Type Calcium Ion Channel cch1 affects Ascospore Discharge and Mycelial Growth in the Filamentous Fungus Gibberella zeae (Anamorph Fusarium graminearum). Eukaryot. Cell 2008, 7, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Lew, R.R.; Abbas, Z.; Anderca, M.I.; Free, S.J. Phenotype of a Mechanosensitive Channel Mutant, mid-1, in a Filamentous Fungus, Neurospora crassa. Eukaryot. Cell 2008, 7, 647–655. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, H.; Qi, L.; Zhang, S.; Zhou, X.; Zhang, Y.; Xu, J.R. FgKin1 Kinase Localizes to the Septal Pore and plays a Role in Hyphal Growth, Ascospore Germination, Pathogenesis, and Localization of Tub1 β-Tubulins in Fusarium graminearum. New Phytol. 2014, 204, 943–954. [Google Scholar] [CrossRef]
- Tassan, J.P.; Le Goff, X. An Overview of the KIN1/PAR-1/MARK Kinase Family. Biol. Cell 2004, 96, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Krause, R.A.; Webster, R. The Morphology, Taxonomy, and Sexuality of the Rice Stem Rot Fungus, Magnaporthe salvinii (Leptosphaeria salvinii). Mycologia 1972, 64, 103–114. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Blackwell, M. The Polyphyletic Origins of Ophiostomatoid Fungi. Mycol. Res. 1994, 98, 1–9. [Google Scholar] [CrossRef]
- Ingold, C. The Stalked Spore-Drop. New Phytol. 1961, 60, 181–183. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sexual Strategy | MAT1-1 Idiomorph | Universal Name | MAT1-1 Genes | α-Factor Pheromone | α-Factor Receptor | MAT1-2 Idiomorph | Universal Name | MAT1-2 Genes | a-Factor Pheromone | a-Factor Receptor |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cryphonectria parasitica | Heterothallic | MAT1-1 | MAT1-1-1 | MAT1-1-1 | Mf1/1 | - | MAT1-2 | MAT1-2-1 | MAT1-2-1 | Mf2/1 Mf2/2 | - |
| MAT1-1-2 | MAT1-1-2 | ||||||||||
| MAT1-1-3 | MAT1-1-3 | ||||||||||
| Magnaporthe grisea | Heterothallic | MAT1-1 a | MAT1-1-1 | MAT1-1 | MF2-1 | ste3-like | MAT1-2 a | MAT1-2-1 | MAT1-2 | MF1-1 | ste2-like |
| Neurospora crassa | Heterothallic | mat A | MAT1-1-1 | matA-1 | ccg4 | pre2 | mat a | MAT1-2-1 MAT1-2-2 | mata-1 mata-2 | mfa-1 | pre1 |
| MAT1-1-2 | matA-2 | ||||||||||
| MAT1-1-3 | matA-3 | ||||||||||
| Trichoderma reesei | Heterothallic | MAT1-1 | MAT1-1-1 | mat1-1-1 | ppg1c | pre2 | MAT1-2 | MAT1-2-1 | mat1-2-1 | hpp1c | pre1 |
| MAT1-1-2 | mat1-1-2 | ||||||||||
| MAT1-1-3 | mat1-1-3 | ||||||||||
| Podospora anserina | Pseudo-homothallic | mat- | MAT1-1-1 | FMR1 | mfm | pre2 | mat+ | MAT1-2-1 | FPR1 | mfp | pre1 |
| MAT1-1-2 | SMR1 | ||||||||||
| MAT1-1-3 | SMR2 | ||||||||||
| Sordaria macrospora | Homothallic b | mat A | MAT1-1-1 | SmtA-1 | ppg1 | pre2 | mat a | MAT1-2-1 | Smta-1 | ppg2 | pre1 |
| MAT1-1-2 | SmtA-2 | ||||||||||
| MAT1-1-3 | SmtA-3 | ||||||||||
| Aspergillus nidulans | Homothallic b | MAT-1 | MAT1-1-1 | MAT1-1 | ppgA | preB | MAT-2 | MAT1-2-1 | MAT2-1 | - | preA |
| Fusarium graminearum | Homothallic b | MAT1-1 | MAT1-1-1 | MAT1-1-1 | ppg1 | pre2 | MAT1-2 | MAT1-2-1 | MAT1-2-1 | ppg2 | pre1 |
| MAT1-1-2 | MAT1-1-2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, A.M.; Wilken, P.M.; van der Nest, M.A.; Wingfield, M.J.; Wingfield, B.D. It’s All in the Genes: The Regulatory Pathways of Sexual Reproduction in Filamentous Ascomycetes. Genes 2019, 10, 330. https://doi.org/10.3390/genes10050330
Wilson AM, Wilken PM, van der Nest MA, Wingfield MJ, Wingfield BD. It’s All in the Genes: The Regulatory Pathways of Sexual Reproduction in Filamentous Ascomycetes. Genes. 2019; 10(5):330. https://doi.org/10.3390/genes10050330
Chicago/Turabian StyleWilson, Andi M., P. Markus Wilken, Magriet A. van der Nest, Michael J. Wingfield, and Brenda D. Wingfield. 2019. "It’s All in the Genes: The Regulatory Pathways of Sexual Reproduction in Filamentous Ascomycetes" Genes 10, no. 5: 330. https://doi.org/10.3390/genes10050330
APA StyleWilson, A. M., Wilken, P. M., van der Nest, M. A., Wingfield, M. J., & Wingfield, B. D. (2019). It’s All in the Genes: The Regulatory Pathways of Sexual Reproduction in Filamentous Ascomycetes. Genes, 10(5), 330. https://doi.org/10.3390/genes10050330